
Б. Б. Бадмаев (ответственный редактор), М. А. Ербаева, О. В. Брандлер
| Вид материала | Тезисы |
- Редактор: А. Т. Горяев Ответственный секретарь: Бадмаев, 2464.34kb.
- М. Л. Энтин (руководитель проекта, ответственный редактор), М. Е. Юрьев, 2654.81kb.
- Вспомогательные исторические дисциплины в пространстве гуманитарного знания, 60.56kb.
- А. С. Панарин (введение, разд. I, гл. 1-4) (ответственный редактор); профессор, 6034.52kb.
- А. Н. Соловьева Ответственный редактор: доктор филологических наук, профессор пгу имени, 5937.63kb.
- Оппозиция, 4805.58kb.
- Гриневич Геннадий Станиславович – Праславянская письменность, 8050.06kb.
- Н. А. Акатова ответственный редактор, 524.95kb.
- Кодексу российской федерации (постатейный), 15526.4kb.
- Б. М. Бернштейн За чертой Апеллеса, 679.73kb.
^ АСИММЕТРИЯ ЧЕРЕПОВ СУРКОВ МОНГОЛИИ
The asymmetry of the marmot sculls from Моngolia
Суханова М.С.
Sukhanova М.S.
ГНУ Всероссийский НИИ охотничьего хозяйства и звероводства
им. проф. Б.М.Житкова Россельхозакадемии, Россия, sms-mari@mail.ru
В ходе проводимых исследований при измерении промеров на черепах часто приходится сталкиваться с их плохой сохранностью. Оценка степени асимметричности черепов поможет установить возможность использования в исследованиях промеров снятых с разных сторон черепа. Определение степени асимметрии поможет установить также стабильность развития и состояние популяций исследуемых видов (Захаров и др., 2000).
Билатеральная симметрия – симметрия левого и правого. Асимметрия это различные отклонения от строгой симметрии.
Наиболее простым и доступным способом оценки асимметрии является учет различий в значениях признака слева и справа.
I метод (Захаров, 1987) применялся для оценки асимметрии каждого параметра по совокупности исследованных черепов.
Для оценки флуктуирующей асимметрии используется величина соответствующей дисперсии:
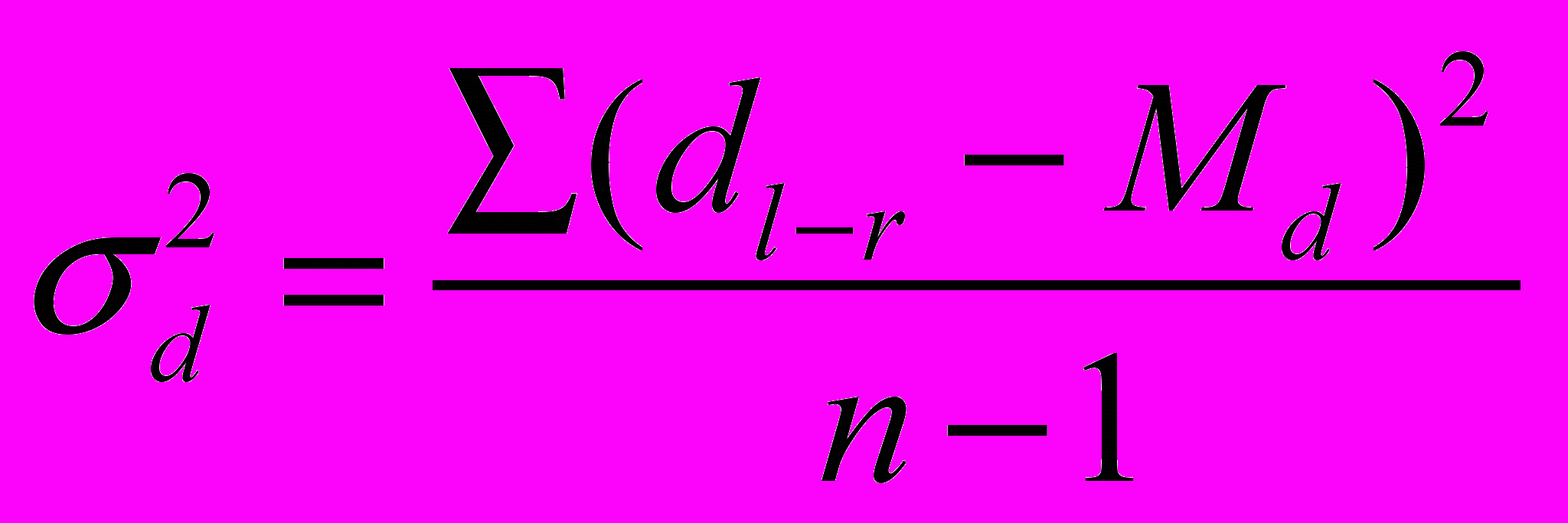 , где
, где 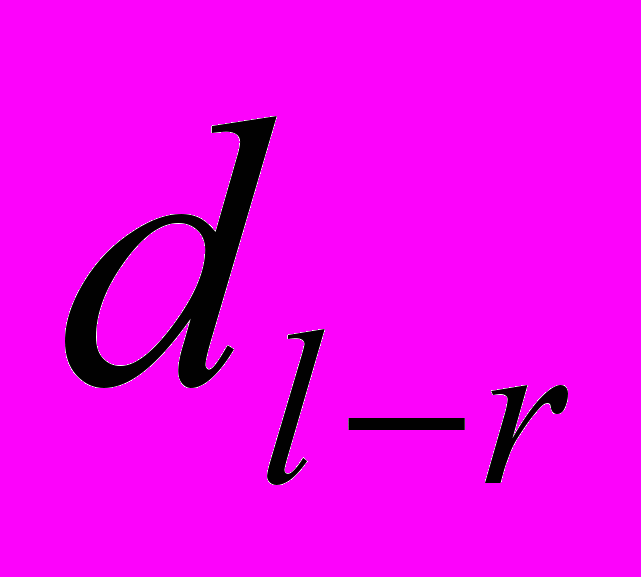 – различие между сторонами,
– различие между сторонами, 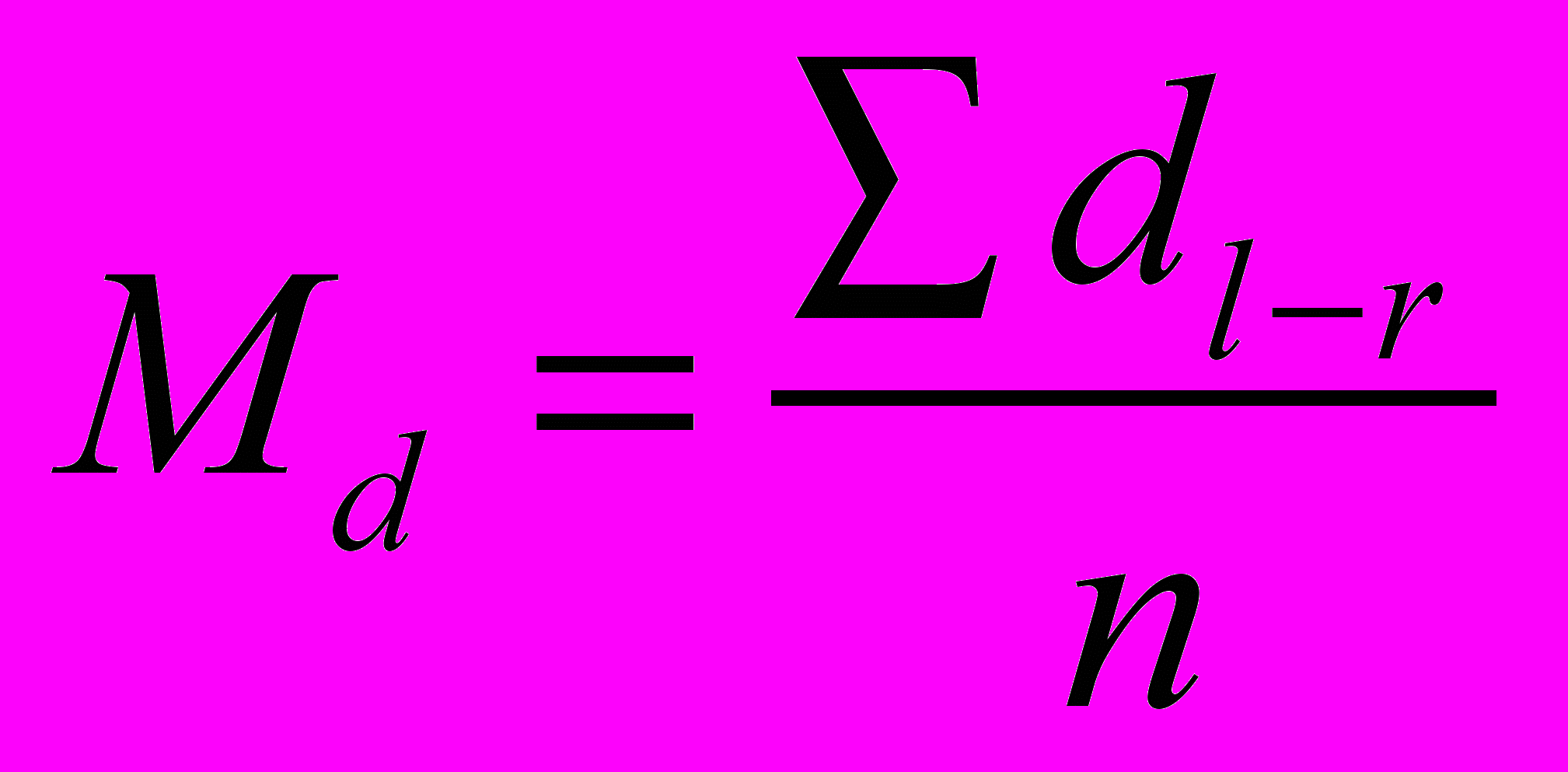 – среднее различие между сторонами, n – количество объектов исследования.
– среднее различие между сторонами, n – количество объектов исследования.Значимость различия при этом оценивается с использованием t-критерия:
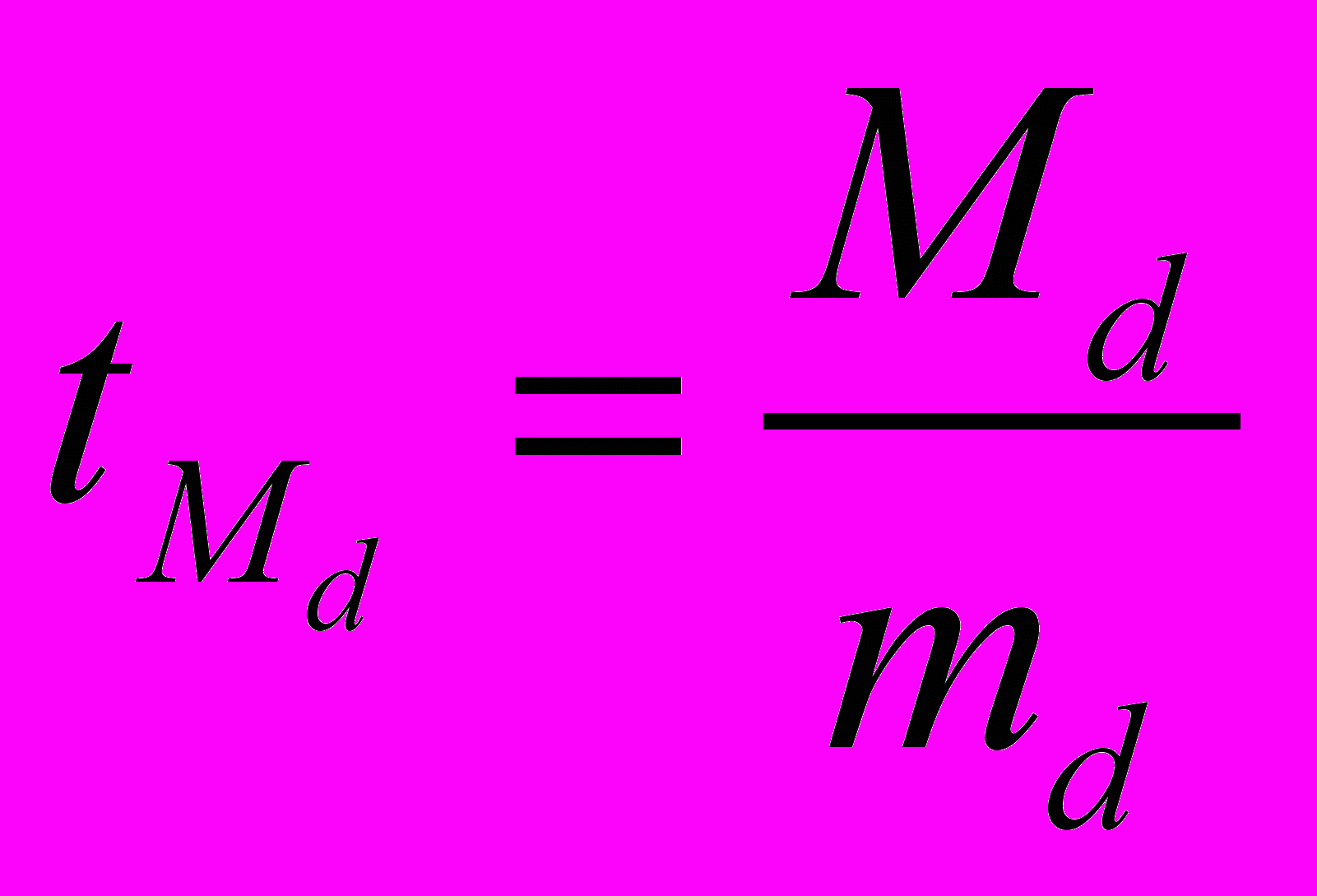 , где
, где 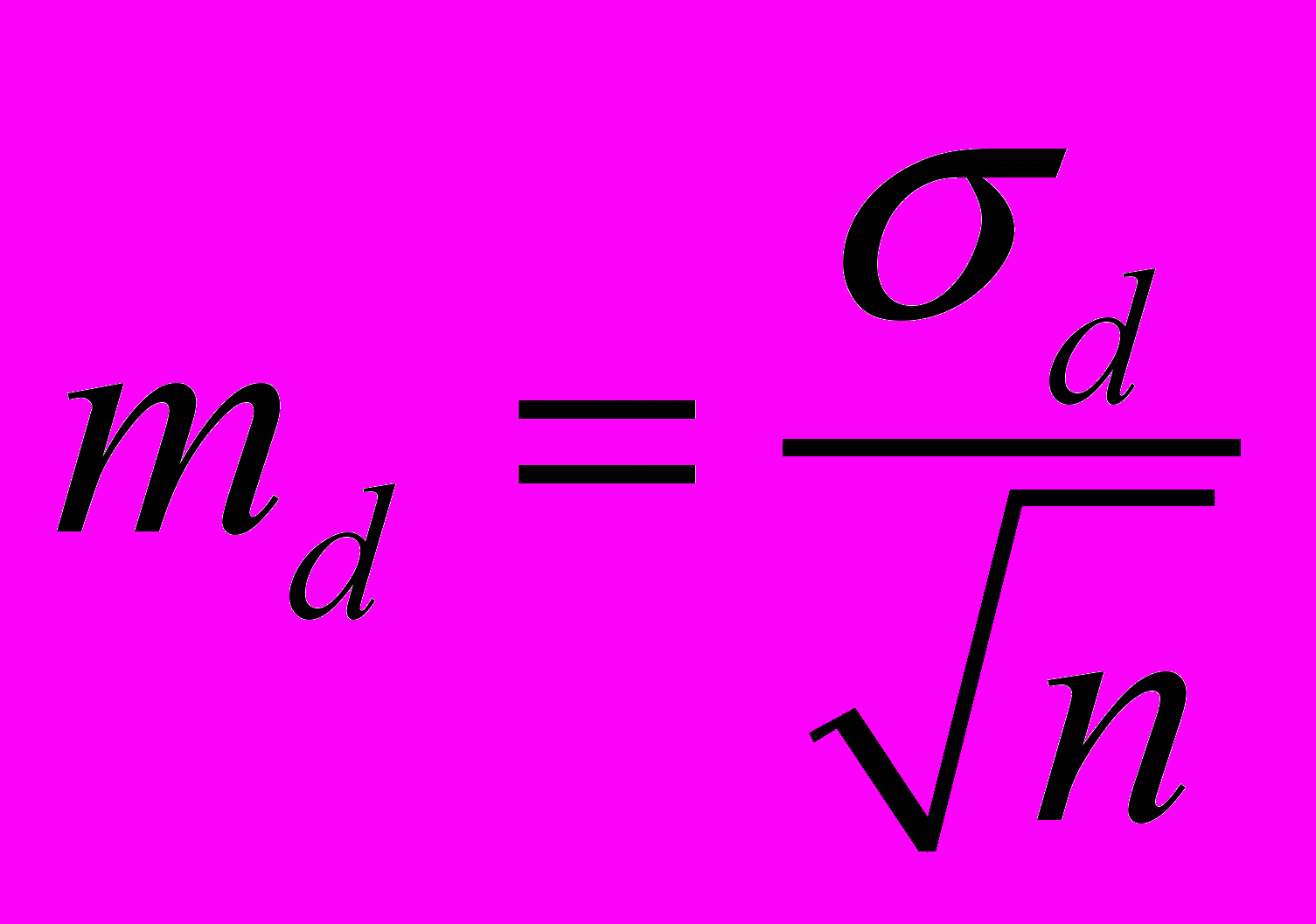 ,
, 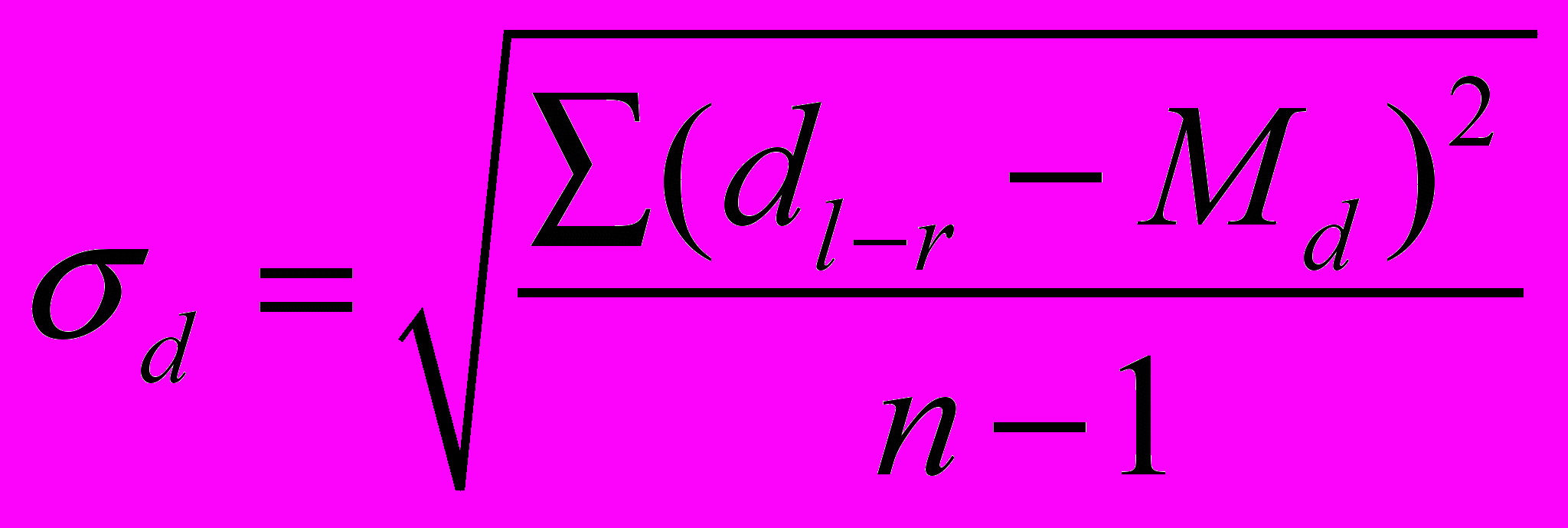 .
.Оценку степени асимметрии производят по дисперсии: чем ближе данный показатель к нулю, тем симметричнее параметр.
При проведении оценки асимметрии I методом можно сказать, что практически все параметры за исключением единичных лежат в пределах от 0,02 до 0,2, что говорит о их симметричности у обоих видов. Самый асимметричный параметр для серого сурка – промер от зарезцовых отверстий до края твердого неба (0,67), для тарбагана этот показатель симметричен (0,13). Еще один показатель, обладающий средней симметрией (0,47) также выражен только серого сурка – это длина рострума. Ширина слухового барабана и длина нижней челюсти – эти два параметра находятся в пределах от 0,33 до 0,45, что говорит о их средней симметрии, и характерны обоим рассматриваемым видам. Значимость полученных результатов очень высокая 99,0–99,9%.
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
II метод (Захаров и др., 2000) применялся для оценки асимметрии каждого черепа по совокупности параметров и в целом для популяции. Величина асимметрии у особи рассчитывается как различие в промерах слева и справа, отнесенное к сумме промеров на двух сторонах. Популяционная оценка выражается средней арифметической этой величины. Статистическая значимость различий между выборками определяется по t-критерию Стьюдента.
По результатам оценки асимметрии по II методу можно сказать, что показатели асимметрии всех черепов находятся в пределах от 0 до 0,025, что говорит о хорошей симметрии всех черепов как для серого сурка, так и для тарбагана. Общий показатель асимметрии популяции серого сурка составляет 0,017, тарбагана – 0,016. Значимость полученных результатов соответствует 95,0%<Р<99,0%.
По результатам всей проделанной работы можно сделать следующие выводы:
Все параметры, взятые с черепов сурков, являются симметричными или средне симметричными, за исключением промера от зарезцовых отверстий до края твердого неба для серого сурка.
Краниометрические показатели популяций обоих видов сурков являются симметричными.
При проведении исследований промеры можно производить как с левой, так и с правой стороны черепа.
^ ВНУТРИВИДОВАЯ СТРУКТУРА СЕРОГО СУРКА
MARMOTA BAIBACINA KASTSCHENKO, 1899 (RODENTIA, SCIURIDAE)
The intraspecific structure of grey marmot Marmota baibacina Kastschenko, 1899
(Rodentia, Sciuridae)
^ Тараненко Д.Е.
Таrаnеnkо D.Е.
Институт систематики и экологии животных СО РАН, Россия, schturm@ngs.ru
В процессе развития таксономии сурков взгляды на внутривидовое деление ^ M. baibacina неоднократно менялись. Анализ литературы показывает, что до настоящего времени единого мнения среди специалистов нет. Так, до середины прошлого века алтайского сурка делили на два подвида: M. b. baibacina Kastschenko (1899) – алтайский подвид; M. b. centralis Thomas (1909) – тяньшанский подвид. Впоследствии было описано ещё два: M. b. ognevi Skalon (1950) – из высокогорий Юго-Западного Алтая; M. b. kastschenkoi Stroganov et Yudin (1956). – лесостепной сурок, обитающий по отрогам Салаирского кряжа и Колывань-Томской возвышенности (Скалон, 1950; Строганов, Юдин, 1956). К перечисленным подвидам Л.И. Галкина добавила кызылрайского M. b. aphanasievi Bashanov (1953), а тяньшанский подвид отелила от M. baibacina (Галкина, 1970).
Таким образом, цель нашей работы: ревизовать внутривидовую структуру M. baibacina. На основании анализа морфологических показателей сурков относящихся ко всем перечисленным формам мы считаем наиболее обоснованным деление M. baibacina на два подвида – M. b. baibacina и M. b. centralis. M. b. ognevi считаем синонимом M. b. baibacina, а форме M. b. kastschenkoi, вслед за Л.И. Галкиной (1962) и О.В. Брандлером (1999, 2003), придаем видовой статус.
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
^ ПРЕДВАРИТЕЛЬНЫЕ РЕЗУЛЬТАТЫ МОРФОЛОГИЧЕСКОГО И МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКОГО АНАЛИЗА СЕРОГО И СТЕПНОГО СУРКОВ (RODENTIA, SCIURIDAE) ИЗ ЗОНЫ КОНТАКТА
В КАЗАХСКОМ МЕЛКОСОПОЧНИКЕ
Preliminary results of the morphological and molecular genetic analysis of grey
and steppe marmots (Rodentia, Sciuridae) from the zone of their contact
in Kazakhskij low mountains
^ Тараненко Д.Е.1, Поляков А.В.2
Таrаnеnkо D.Е., Polyakov A.V.
1Институт систематики и экологии животных СО РАН, Россия, schturm@ngs.ru
2Институт цитологии и генетики СО РАН, Россия, polyakov@bionet.nsc.ru
Проблема систематического статуса представителей рода сурков (^ Marmota), относящихся к группе «bobak», инициировала обсуждение возможности межвидовой гибридизации серого (Marmota baibacina Kastschenko, 1899) и степного (M. bobak Müller, 1776) сурков в зоне перекрывания их ареалов (Галкина, 1962; Громов 1965; Капитонов, 1966). Долгое время этот вопрос остается не решенным из-за недостаточности исследованного материала и относительно высокой вариабельности отдельных морфологических признаков, используемых в анализе. Развитие современных методов исследования, таких как геометрическая морфометрия и молекулярно-генетический анализ позволяет использовать дополнительные возможности для развития данной темы.
Ареалы серого и степного сурков формируют узкую зону контакта на территории Казахского мелкосопочника (Карагандинская область, Казахстан). Именно из района этой зоны был произведен сбор представителей двух видов сурков. Сбор проводился из трансекта в направлении с востока на запад, начиная с западной границы ареала серого сурка, затем в зоне стыка ареалов серого и степного и, затем, в пределах восточной границы ареала степного сурка.
Результаты анализа габитуальных параметров и мест обитания позволяют однозначно идентифицировать видовую принадлежность особей из крайних точек трансекта. Выявление наличия взаимодействия представителей контактирующих популяций двух видов сурка и потока генов между ними проводится в настоящее время с помощью краниометрического анализа полученного материала методом геометрической морфометрии с привлечением выборок из удалённых географических популяций. Изучение характера распространения в исследуемом трансекте молекулярно-генетических маркеров позволит уточнить филогенетические взаимосвязи представителей контактирующих популяций и сделать заключение о возможности межвидовой гибридизации.
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
^ ГИМАЛАЙСКИЙ СУРОК MARMOTA HIMALAYANA, HODGSON (1841)
В АНТРОПОГЕННОМ ЛАНДШАФТЕ НЕПАЛА
The Himalayan marmot (Marmota himalayana Hodgson,1841)
in the аnthropogenic landscapes of Nepal
Улак А.1, Никольский А.А.2
Ulak A., Nikolskij A.A.
1Travel ways international, Nepal (Непал), atmarasi@hotmail.com
2Российский университет дружбы народов, Россия, bobak@list.ru
Материалом для данного сообщения послужили наблюдения, проведённые нами на территории Непала в 2003, 2004, 2006 и в 2009 гг. в регионах Манаслу, Нижний Мустанг, Соло Кхумбо. “Конфликт интересов” сурков и людей заключается в том, что те и другие используют для своих поселений одни и те же формы рельефа, населяя преимущественно аллювиальные террасы с мощным слоем мелкозема. Сурки используют мелкозем как субстрат, в толще которого устраивают норы. Для человека аллювиальные отложения удобны тем, что на них развиваются пригодные для земледелия почвы. Широкие выровненные террасы удобны так же и для выпаса скота. Основными формами антропогенного воздействия на сурков являются: 1) поселения человека и сельскохозяйственные угодья, которые приводят к уничтожению мест обитания и расчленению поселений сурков; 2) выпас скота, влияющий на кормовые условия сурков, и 3) неконтролируемый промысел. Все поселения гималайского сурка расположены на выпасах домашних животных, подверженных пастбищной дигрессии. Местное население регулярно добывает сурков петлями. Когда люди забрасывают поля и долго не обрабатывают их, сурки возвращаются на прежние участки обитания. Они могут оставаться на соседних с полями участках, и даже заселять заброшенные постройки. В 2006 г. в Нижнем Мустанге мы обнаружили многочисленные нежилые норы сурков. Некоторые из них содержали следы отлова животных петлями. Нижний Мустанг является южной окраиной огромного поселения сурков, центр которого находится в Верхнем Мустанге, на границе Тибета и Непала. В регионе Соло Кхумбо мы не обнаружили сурков, хотя в литературе есть указания на его обитание в национальном парка “Сагармата”, расположенном в данном регионе. Сурок в Непале не входит в категорию редких, или угрожаемых видов. Целесообразно вести регулярный мониторинг над численностью сурков в Непале. Необходимо установить категорию статуса гималайского сурка. Его промысел следует регулировать или полностью запретить. Серьёзным препятствием к организации охраны гималайского сурка является его слабая изученность, приобретая иногда трагикомические формы. Так, в 2008 г. на непали в Катманду вышла книга “Дикие млекопитающие Непала” под эгидой таких солидных организаций как Фонд охраны дикой природы (WWF) и Международный союз охраны природы (IUCN). Книга роскошно издана и подготовлена европейско-непальской командой из 20 авторов. Согласно этому изданию, Непал населяют два вида: байбак (Bobak Marmot, Marmota bobak) и гималайский сурок (Himalayan Marmot, M. himalayana). По мнению авторов издания, виды занимают практически один и тот же ареал. Книга адресована школьникам, студентам и преподавателям. Нетрудно представить, с какими трудностями столкнутся потенциальные читатели, искренне желающие познать природу родного края.
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
^ СОХРАНЕНИЕ ПОПУЛЯЦИИ ТАРБАГАНА (MARMOTA SIBIRICA SIBIRICA RADDE, 1862) В РЕГИОНАЛЬНОМ ЗАКАЗНИКЕ «ГОРНАЯ СТЕПЬ»
The protection of tarbagan (Marmota sibirica sibirica Radde, 1862) population
in the regional zakaznik «Mountain steppe»
^ Ушакова М.С.
Ushakоvа М.S.
Сохондинский государственный природный биосферный заповедник, Россия, sochondo@rambler.ru
В данной работе приводятся данные, полученные в результате наблюдений за состоянием популяции монгольского сурка, или тарбагана (Marmota sibirica sibirica Radde, 1862) на территории заказника «Горная степь» в 2005-2008 гг.
Государственный природный комплексный заказник регионального значения «Горная степь» находится на юге Восточного Забайкалья, примыкает к российско-монгольской границе (в пределах Кыринского района Забайкальского края). «Горная степь» входит в состав проектируемой единой трансграничной особо охраняемой территории «Истоки Амура». Региональный заказник «Горная степь» образован 9 декабря 2003 г. с целью восстановления и сохранения естественных экосистем Даурской горной степи, охраны и поддержания численности редких и исчезающих видов растений и животных. Со времени образования заказника в Кыринском районе взяты под государственную охрану практически все представители флоры и фауны юга Восточного Забайкалья.
Территория заказника «Горная степь» расположена от устья п. Галзутый до устья п. Нарын, включает в себя бассейны падей Нарын, Сухая, Глубокая, Большой и Малый Кожуртаны. Протяженность «Горной степи» вдоль государственной границы составляет 10 км, площадь этой ООПТ равна чуть более 5 тыс. га.
Начиная с 2004 г., с момента установления режима охраны, на территории заказника численность тарбагана стала возрастать. В 2006 г. она достигла значения в 160-180 особей, в 2007 г. - не менее 200 особей. Однако в 2008 г. было отмечено снижение численности (всего 112 особей). Вероятно, существенное сокращение численности монгольского сурка в «Горной степи» произошло в результате степных пожаров весны 2008 г.
Таким образом, в связи с охраняемым положением в государственном природном комплексном заказнике регионального значения «Горная степь» тарбаган находит на его территории наиболее благоприятные условия для жизни. Об этом свидетельствует возрастание численности этого сурка после образования заказника. На территории ООПТ «Горная степь» сосредоточено около 40% от численности всей популяции тарбагана на юге Восточного Забайкалья.
Ещё до создания заказника было прекращено животноводство, вследствие чего был устранен один из антропогенных факторов, влияющих на кормовые условия сурков. Сенокошение в настоящее время производится в основном жителями с. Гавань и в ограниченном количестве. Однако, одним из основных лимитирующих факторов, сдерживающим реально возможный рост численности популяции тарбагана, являются весенние степные пожары. Они подрывают кормовую базу монгольского сурка в период выхода его из зимней спячки, когда энергетический баланс сурка минимален.
В 2008 г. на территории «Горной степи» (п. Нарын, восточная граница заказника) был построен еще один кордон. Его наличие позволило организовать постоянный контроль практически за всей территорией заказника, в т.ч. контроль за выявлением и своевременным предупреждением пожаров, случаев браконьерства и т.п. Все это
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
способствует оптимизации режима охраны в заказнике, а значит и сохранению популяции тарбагана на данной площади.
^ ЗАКОНОМЕРНОСТИ РОСТА И РАЗВИТИЯ СУРКОВ (MARMOTA BOBAC MULL.) КЛЕТОЧНОГО РАЗВЕДЕНИЯ В ПОСТНАТАЛЬНОМ ОНТОГЕНЕЗЕ
The regularities of growth and development of the cage breeding marmots
in the postnatal ontogenesis
^ Федорова О.И.
Fеdоrоvа О.I.
Московская государственная академия ветеринарной медицины и биотехнологии
имени К.И. Скрябина, Россия, ox_fed@mail.ru
Литературные данные о развитии молодняка сурков в природе малочисленны. В основном они касаются периода выхода сурчат из нор, то есть возраста молодняка после 40-50 дней. У сурка Мензбира масса сурчат в этот период составляет 195-235 (в среднем 221) г, длина тела 195-235 (218) мм, задней ступни 48,4 -57,2 (53,7) мм, хвоста 53-70 (70) мм, уха 13,9 - 18,3 (16,6) мм (В.И. Машкин, 1993). По европейскому байбаку имеются данные о массе тела сурчат при выходе из нор – 240-260 г (В.И. Машкин, 1997). Длина новорожденных клеточных сурчат – 8 см (А.Ю. Максимов, 1993).
Работа по изучению постнатального онтогенеза сурков проводилась в ГПЗ «Пушкинский» на всем поголовье экспериментального стада сурков байбаков (Marmota bobac Mull).
Исследование формирования первичного опушения сурчат, родившихся от родителей, живущих в клетках, показало, что оно мало чем отличается от животных, обитающих на воле. Щенки родились голыми и имели розовую кожу. Уже на второй день кожа стала фиолетово-серой, появился подрост первичных волос. До 18 дней волосы на спине были темно-серого цвета, на брюшке золотистого. В 18-20 дней опушение на верхней части животных (шее, лопатках, хребте, боках, огузке) густое, золотистого цвета с черной рябью. Длина волос на боку 7-10 мм. Цвет кожи темно-синий. К месячному возрасту сурчата имели густой волосяной покров (высотой до 21 мм) золотистого цвета с темно-серой вуалью.
В возрасте 45 дней рост ювенальных волос у сурчат прекратился, и было зарегистрировано выпадение отдельных волос первичного опушения. Цвет кожи слегка синеватый. Строение корней волос соответствовало законченному росту волоса, т.е. корень и самая нижняя часть основания стержня чистые, светлые; серцевинный слой в корнях волос отсутствовал. Полное выпадение ювенальных волос завершилось к 80-90-дневному возрасту. Одновременно с выпадением первичных волос произошли закладка и рост волос вторичного волосяного покрова. В возрасте 65 дней подрост от 0,5 до 10 мм был зарегистрирован у всех сурчат. К 100-дневному возрасту высота волосяного покрова достигла 23,1 мм, при длине пуховых волос 14,9.
Для изучения роста сурков в постнатальный период всех полученных сурчат (32 головы) взвешивали со дня рождения до первой спячки, а также измеряли длину тела каждые 10 дней.
Измерения новорожденных сурчат показали, что при рождении длина тела их составляет в среднем 9 – 10 см (9,30,3 у самок, 9,70,2 у самцов), а живая масса 29-30 г (30,80,8 у самок, 29,11,0 у самцов). К месячному возрасту живая масса сурчат
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
увеличивается более чем в 10 раз и составляет у самок в среднем 383,6 г, а у самцов - 432 г. Длина тела увеличивается к этому возрасту в 2,5 раза. У самок она составляет в среднем 22,9 см, у самцов – 24,7 см.
За 100 дней (от 45 до 145-дневного возраста) длина тела самок увеличилась на 61,3%, самцов – на 60,7% . Больший интенсивный рост отмечен у сурчат по живой массе. У самок за 100 дней живая масса увеличилась в 5,2 раза, у самцов в 5,8 раза и составила к 145-дневному возрасту 4580 г у самок и 4642 г у самцов.
За время первой спячки длина тела у самцов увеличилась в среднем на 1,5 см, у самок на 1 см, а живая масса уменьшилась в среднем на 1032 и 1115 г, соответственно (Р >0,999). Ко второй спячке живая масса у самок сурков составила в среднем 6200 132 г, у самцов 6450148 г, а длина тела 53,4 и 55,2 0,5, соответственно.
Выводы:
Живая масса сурчат (родившихся в клеточных условиях) увеличилась от дня рождения до первой спячки более чем в 150 раз, а длина тела в 5 раз.
В период от рождения до спячки более интенсивный рост живой массы и длины тела у сурчат наблюдается в первые 45 дней.
На первом году жизни у сурчат по живой массе и длине тела не наблюдается полового диморфизма.
В процессе доместикации сурков заметно их превосходство по средней живой массе и длине тела над их сородичами в природе.
^ МОРФОЛОГИЯ РЕПРОДУКТИВНОЙ СИСТЕМЫ СУРКОВ MARMOTA BOBAK
The morphology of the reproductive system in Marmota bobak
Федосеева Г.А.1, Сунцова Н.А.2, Плотников И.А.3
Fedoseeva G.A., Suntsova N.A., Plotnikov I.A.
1ГНУ НИИ пушного звероводства и кролиководства, Россия, niipzk@.mail.ru
2ФГОУ ВПО ВГСХА, Россия,
3ГНУ Всероссийский НИИ охотничьего хозяйства и звероводства
Россельхозакадемии, Россия, bio.vniioz@mail.ru
В период убоя для исследования были взяты семенники у самцов и яичники у самок. По самцам было две группы животных: первая - самцы, от которых самки щенились, вторая - самцы, от которых самки не щенились.
Семенники самцов снаружи покрыты серозной оболочкой, под ней расположена соединительно-тканная белочная оболочка. Внутри семенника имеется соединительно- тканное средостение семенника. От него до белочной оболочки отходят перегородки, разделяющие семенник на множество долек. Паренхима состоит из извивающихся семенных трубочек. Трубочки соединены между собой межтубулярной рыхлой соединительной тканью. Строма содержит интерстициальные клетки или клетки Лейдига, размером 5х10 мкм. Вблизи средостения трубочки соединяются друг с другом и переходят в прямые семенные канатики, не участвующие в спермиогенезе. Белочная оболочка у сурков толщиной 0,1-0,4 мм, состоит из коллагеновых волокон с примесью эластических. Дольки семенника у сурков длинной конической формы, септулы между ними хорошо развиты. Извитые семенные канальцы диаметром 50 - 160 мкм. В извитом семенном канальце самцов, от которых самки щенились, спермиогенные клетки расположены между
X Международное совещание по суркам стран СНГ – 10-th International Meeting on Marmots of CIS countries
_______________________________________________________________________________________________________
фолликулярными клетками. На периферии канальца выявляются спермогонии, затем слой первичных и вторичных спермиоцитов, слой сперматид и слой спермиев.
У самцов, от которых самки не щенились, наблюдается атрофия сперматогенного эпителия. Дегенеративные процессы сильнее выражены в наиболее дифференцированных слоях сперматогенного эпителия – в формирующихся сперматозоидах и сперматидах. Набухшие клетки сливаются и в просвете семенного канальца выявляются семенные шары.
По самкам также было две группы животных: первая - самки, приносившие приплод, вторая - самки, не приносившие приплод.
Было проведено гистологическое исследование яичников у сурков. Яичник самок сурков овальной формы, сплюснутый. Имеет типичное гистологическое строение для млекопитающих. Состоит из коры и медуллы. Кора содержит паренхиматозные структуры органа – яичниковые фолликулы и желтые тела. В медулле или мозговой зоне расположены крупные кровеносные сосуды. Снаружи яичник покрыт зачатковым эпителием. Под ним находится слой, богатый коллагеновыми волокнами – белочная оболочка. Соединительная ткань коры – яичниковая строма заполняет пространство между паренхиматозными элементами. Особенности: зачатковый эпителий - однослойный кубический. В белочной оболочке волокна расположены слоисто и идут в радиальном и поперечном направлении. Толщина этого слоя у самки степного сурка составляет 15 –25 мкм. Волокна, направляющиеся от белочной оболочки к мелким фолликулам, перекрещиваются между собой под острым углом. Пространство между фолликулами и желтыми телами заполнено веретенообразными клетками.
У самок сурков, нормально размножавшихся, в коре находятся яичниковые фолликулы на разных стадиях развития. Первичные фолликулы образуют мелкие скопления. Количество первичных фолликулов на срезе яичника составило 60 – 350, вторичных 80 – 120. Между ними расположены желтые тела в разной стадии развития. Желтые тела самок сурков относятся к эпителиальному типу.
У не размножавшихся самок гистологическая картина иная. Зачатковый эпителий деформирован или отсутствует. Фолликулярная зона сужена. Количество первичных фолликулов снижено до 20 – 50 или отсутствуют. В единичных вторичных фолликулах выявлены дегенеративные процессы, они выражены в сморщивании или лизисе цитоплазмы. На месте атретических фолликулов (дегенеративных) отмечаются многочисленные зоны в виде неправильной формы колец. Паренхима яичника редуцирована. Между фолликулами – разросшаяся соединительная ткань. Кровеносные сосуды облитерированы или гиализированы.
Полученные результаты очень важны для изучения биологии размножения сурков.