
Профессор В. А. Шальнев Вопросы географии и краеведения
| Вид материала | Документы |
| Ставропольский государственный университет |
- В. М. учитель географии, биологии и краеведения, высшей категории, Ношенской оош., 36.1kb.
- Вопросы, вопросы. На все эти вопросы учащиеся находят ответы на уроках биологии и географии, 36.93kb.
- Региональный курс краеведения (7-8 кл.), в рамках которого изучается ряд проблем, 628.85kb.
- Географии и краеведения, 1771.64kb.
- Захарова Елена Викторовна, учитель начальных классов моу«сошуиоп №8» Формы и методы, 21.42kb.
- Школьного географического краеведения стр, 311.91kb.
- Методические рекомендации по методике организации и проведению учебно-тематических,, 400.41kb.
- Учебное пособие историко-культурные туристские ресурсы Северного Кавказа для студентов, 496.84kb.
- Учебное пособие историко-культурные туристские ресурсы Северного Кавказа для студентов, 671.13kb.
- Кутняхова Наталья Анатольевна, аспирантка кафедры литературы и методики преподавания, 75.18kb.
Литература
Глазовская М.А. Методологические основы типологии и методики исследования природных ландшафтов. М.: Изд-во Моск. Гос. Ун – та, 1964. - 230с.
Исаченко А.Г. Ландшафтоведение и физико-географическое районирование. – М.: Высшая школа, 1991.
Преображенский В.С. Феномен в географии. Раздумья географа // АН СССР Сер. Геогр. – 1979. - №4. – С. 20-27.
Т.В.Дегтярева, В.Н.Мамонтов
Ставропольский государственный университет
Геохимия ландшафтов музея-заповедника «Татарское городище»
Применение методов геохимии ландшафтов при анализе заповедных территорий позволяет получить данные о фоновом состоянии и функционировании ландшафтных систем. Природно-археологический комплекс музея-заповедника «Татарское городище» располагается на южном склоне Ставропольской горы и, примыкая к юго-западной окраине урбанизированной зоны г.Ставрополя, находится в зоне техногенного геохимического воздействия со стороны города. Заповедник входит в Верхнеегорлыкский водораздельный окультуренный ландшафт типичных лесостепей провинции лесостепных ландшафтов Ставропольского края (Шальнев, 1995). Территории присуща ярусность рельефа и в ландшафтном отношении выделяются две местности: структурно-денудационное останцовое плато и речные долины (Археологический и природный…, 1999).
Почвенно-геохимическое опробование проводилось по двум ландшафтным профилям, охватывающим типичные для данной территории урочища: плакоры окраин платообразных поверхностей выравнивания, крутые склоны останцовых возвышенностей, речные долины раннеакчагыльской эпохи формирования. В почвенных образцах с опробованием по всему почвенному профилю определялись щелочно-кислотные условия, содержание гумуса методом И.В.Тюрина, содержание кислоторастворимых форм металлов (Pb, Cd, Cu, Zn) методом вольтамперометрического анализа (ИВА) в лаборатории почвоведения и геохимии ландшафтов СГУ. При статистической обработке результатов анализов было рассчитано среднее содержание элементов в почвах морфологических единиц ландшафтов на данной территории, которые могут рассматриваться в качестве фоновых (табл.1).
Таблица 1
Некоторые параметры почв ландшафтов музея-заповедника
| Морфологические единицы ландшафтов | рH | Содержание гумуса, % | Содержание микроэлементов, мг/кг | |||
| Свинец | Кадмий | Медь | Цинк | |||
| Плакоры окраин поверхности выравнивания | 7,4 | 6,8 | 1,3 | 0,17 | 3,26 | 6 |
| Крутые склоны останцовых возвышенностей | 8,1 | 4,4 | 0.75 | 0,15 | 2,3 | 2,6 |
| Верхние части склонов верховьев речных долин | 7,7 | 6,5 | 1,08 | 0,24 | 1,96 | 3,3 |
| Нижняя часть склонов речных долин | 7,1 | 6,3 | 1,32 | 0,22 | 2,26 | 4,16 |
Плакоры окраин платообразных верхнесарматских поверхностей выравнивания отличаются элювиальными условиями миграции элементов. Сформированные здесь выщелоченные черноземы (Антыков, Стомарев, 1970) имеют черновато-серую окраску гумусовых горизонтов, тяжелосуглинистый состав, четко выраженную комковато-зернистую структуру, нейтральную реакцию (рH=7,4). Содержание гумуса высокое и составляет 7-7,3%, с глубиной гумуссированность падает постепенно. При радиальном распределении элементов характерна поверхностная аккумуляция свинца и меди, содержание цинка увеличивается к породе. Относительно почвообразующей породы почвы обеднены свинцом в 1,7 раза, кадмием в 4,5 раза, медью в 1,2 раза. В то же время цинка в почве содержится в 2 раза больше, чем в породе, что может быть следствием биогенного накопления или аэрогенного антропогенного поступления.
На крутых склонах останцовых поверхностей выравнивания с трансэлювиальными условиями миграции продуктов почвообразования сформированы смытые деградированные черноземы. Гумусовые горизонты серо-бурой окраски имеют слабощелочную реакцию (рH=8-8,3), содержание гумуса понижается до 4%, структура почвы более распылена, гранулометрический состав почв легкосуглинистый. Накопление в верхних горизонтах при радиальном распределении характерно для свинца, меди и цинка. В сравнении с породами в почвах рассеивается свинец (кларк рассеивания Кр=2,8), медь (Кр=1,3), кадмий (Кр=6) и концентрируется цинк (кларк концентрации Кк=1,26).
Почвы верхних частей склонов верховьев речных долин и балок представлены смытыми темносерыми лесными почвами, для которых характерна нейтральная реакция среды в верхних горизонтах (рH=7,6) с усилением слабощелочной реакции в нижних горизонтах (рH=7,9). Содержание гумуса на глубине 10 см – 7,8%, на глубине 50 см – 6,6%. При радиальном профильном распределении элементов их четкой биогенной аккумуляции нет, вероятно, за счет постоянного смыва гумусовых горизонтов почв. Для свинца и меди идет увеличение концентраций по профилю к породе.
В пределах нижних частей склонов речных долин второго и третьего порядков раннеакчагыльской эпохи формирования под буково-грабовыми лесами развиты темно-серые лесные почвы. Их отличает суглинистый гранулометрический состав, нейтральная реакция среды (рH=6,9-7,25), содержание гумуса 5,4-6,6%. Аккумулятивные условия миграции приводят к концентрации в почвах относительно пород всех рассматриваемых элементов: свинца (Кк=1,14), меди (Кк=1,02), цинка (Кк=1,2), кадмия (Кк=1,8).
Содержание элементов в почвах музея-заповедника зависит от почвообразующей породы (табл.2). Анализ распределения концентраций ТМ показал, что почвы, сформированные на известняках среднего сармата, содержат самые высокие значения цинка (6,85 мг/кг). Почвы, сформированные на песках среднего сармата, обеднены комплексом элементов; для них характерны самые низкие уровни содержания кадмия (0,21 мг/кг), свинца (0,35 мг/кг), цинка (2,7 мг/кг). Почвы, образованные на глинах среднего сармата, обогащены свинцом и медью, для которых выявлены максимальные уровни накопления (свинца – 1,2 мг/кг, меди – 1,9 мг/кг).
Таблица 2
Среднее содержание элементов в почвах, сформированных на различных почвообразующих породах, мг/кг
| Почвообразующая порода | Свинец | Кадмий | Медь | Цинк |
| Известняки среднего сармата | 0,65 | 0,25 | 1,75 | 6,85 |
| Пески среднего сармата | 0,35 | 0,21 | 1,8 | 2,7 |
| Пески и глины среднего сармата | 0,98 | 0,26 | 1,7 | 4,3 |
| Глины среднего сармата | 1,2 | 0,24 | 1,9 | 4,3 |
Существование ярусности ландшафтного пространства территории музея-заповедника позволяет провести латеральный анализ миграции элементов с помощью коэффициента местной концентрации (Км), представляющего отношение содержания элемента в почвах подчиненных фаций к его содержанию в почвах автономных фаций. Свинец выносится из почв склонов останцовых поверхностей выравнивания (Км=0,55) и из верхних частей склонов верховьев речных долин (Км=0,29). В почвах нижних частей склонов речных долин свинец накапливается (Км=1,9). Латеральная дифференциация кадмия также отличает транзитный вынос из почв склонов останцовых поверхностей выравнивания (Км=0,75) и аккумуляция в почвах нижних частей склонов речных долин (Км=2,7).
Медь и цинк относительно элювиальных позиций плакора выносятся из почв всех ярусов ландшафтной структуры территории заповедника. При этом от крутых склонов останцовых поверхностей выравнивания к нижним частям склонов речных долин идет уменьшение интенсивности выноса элементов: для меди Км 0,36 – 0,75; для цинка Км 0,03 -0,7. Таким образом, латеральное распределение меди и цинка происходит не в соответствии с ландшафтно-геохимическим сопряжением между автономными фациями плакоров платообразных поверхностей выравнивания и подчиненными фациями речных долин. Приуроченность максимальных концентраций меди и цинка к почвам плакоров с элювиальными условиями миграции может быть вызвано антропогенным аэрогенным переносом химических элементов с территории города Ставрополя.
Литература
Антыков А.Я., Стомарев А.Я. Почвы Ставропольского края и их плодородие. - Ставрополь: Кн. изд-во, 1970. - 413 с.
Археологический и природный музей-заповедник «Татарское городище» как природное наследие /Под ред. проф. В.А. Шальнева. – Ставрополь: Изд-во СГУ, 1999. – 100с.
Шальнев В.А. Ландшафты Ставропольского края. - Ставрополь: Изд-во СГУ, 1995.
В.А. Шальнев, Д.В. Олейникова
Ставропольский государственный университет
Пространственный анализ растительного покрова высокогорных ландшафтов северо-западного Кавказа
В условиях все возрастающего антропогенного воздействия происходят сокращения площадей естественных ландшафтов и биоэкосистем с их бесплатными средоформирующими и ресурсовоспроизводящими функциями, что диктует необходимость решения проблем сохранения ландшафтного и биологического разнообразия биосферы.
К числу таких направлений можно отнести биотику ландшафта, под которой понимается научное направление, изучающее территориальное распределение биоты и биомов в рамках геосистем регионального и локального уровней и их роль в функционировании последних на основе ландшафтной парадигмы (Лиховид и др., 2005). Это новое направление в ландшафтоведении, которое стало формироваться на кафедре физической географии Ставропольского госуниверситета с 2000 года в связи с тем, что исследование биотических компонентов ландшафта и в настоящее время остается слабо разработанным звеном теории ландшафтоведения. Более того, изучение этих вопросов происходит в рамках научного направления «Геофизика ландшафта», т.е. природе простейших форм движения материи.
Методологической основой данного направления является ландшафтный подход, где природный ландшафт является реальным объектом исследования и рассматривается в качестве фокуса территориальной организации компонентов биосферы с позиций холизма (целостности взаимодействия абиотического, биотического и биокосного вещества в одном пространстве). Именно биота является тем «метахондрием» в генотипе ландшафтных геосистем, который определяет сущность взаимодействия абиотических, биотических и биокосных компонентов на определенном отрезке существования геосистемы.
В концептуальной основе биотики ландшафта лежит идея о полисистемной модели ландшафта (Шальнев, 1999, 2004). Она предусматривает выделение в рамках теоретического моделирования ландшафта трех подсистем: компонентной, морфологической и биоэкосистемной (биоценотической). В рамках этой модели биотика ландшафта находит возможности своей реализации на всех трех уровнях анализа. Важным для биотики ландшафта является биоценотическая подсистема.
В биоценотической подсистеме (подсистеме биоэкосистемного типа) возможно применение пространственно-временного анализа по принципу монодуализма с функциональной первичностью времени, что проявляется в сезонной динамике биоты, биохимических процессов и круговоротов СЭИ. Имманентная сущность связей в такой модели строится по принципу экоцентрических прямых и обратных связей, а эмерджентные свойства проявляются в поддержании инвариантных свойств качества природной среды жизни биоты. «Хозяин» модели (биота) испытывает не только воздействие компонентов окружающей среды, но и сам влияет на эти компоненты и качество среды. Причина – биохимические процессы и биотические источники энергии, определяющие особенности биогеохимических круговоротов вещества в пределах биоэкосистемы. В таком качестве набор биоэкосистем (в ранге фаций) представляет собой «анатомию» и «физиологию» ландшафтной структуры, которая формирует и регулирует природную среду не только ландшафта, но и биосферы в целом.
В традиционных ландшафтных исследованиях и в настоящее время основное внимание уделяется растительному покрову с позиций компонентного анализа с использованием описательных методов. Внутриландшафтная дифференциация биотического компонента обычно рассматривается на примере изучения биогеоценозов.
Исследования проводились в высокогорьях Западного Кавказа методом ключевых ландшафтов, обеспеченных крупномасштабными ландшафтными картами в ранге урочищ. В среднегорьях и высокогорьях Бокового хребта была выбрана группа ландшафтов, занимавшая западные (Софийский), центральные (Хатипарский) и восточные (Гондарайско-Узункольский) районы Западного Кавказа и позволившая проводить сравнительный анализ на региональном уровне. Исследования, которые проводились в течение шестилетних полевых сезонов (2000-2006 гг.) позволили получить интересные данные и сделать предварительные выводы об особенностях пространственной дифференциации растительного покрова регионального и внутриландшафтного содержания.
Ландшафтный подход в изучении луговой растительности высокогорий Западного Кавказа позволил проследить ее вертикальную и горизонтальную внутриландшафтную дифференциацию и отметить некоторые закономерности этой дифференциации. Причиной вертикальной дифференциации является изменение гидротермических условий с высотой (климатогенных полей однородности) и, как следствие, формирование однородного видового состава в рамках доминантных и переходных геоботанических поясов. Сравнительная характеристика макроэкотопов (геоботанических поясов) Хатипарского ландшафта показана в таблице 1.
Таблица 1.
Доля ботанических групп (луговой растительности) в высотных геоботанических поясах Хатипарского ландшафта в % (Олейникова, 2005)
| Геоботанические пояса | Злаки и осоки | Разнотравье | Бобовые |
| Пояс экотона, границы леса | 53,0 | 44,0 | 3,0 |
| Субальпийский | 53,0 | 29,0 | 18,0 |
| Альпийский | 56,0 | 19,0 | 25,0 |
| Субнивальный | 47,0 | 24,0 | 29,0 |
Для флористического сравнения микротопов строился алгоритм максимального корреляционного пути с использованием коэффициента Жаккара. На его основании построены дендриты (рис. 1, 2, 3), содержащие корреляционные плеяды разного порядка по каждому ключевому участку высокогорного ландшафта, объединяющиеся в районы (Иванов, 1998).
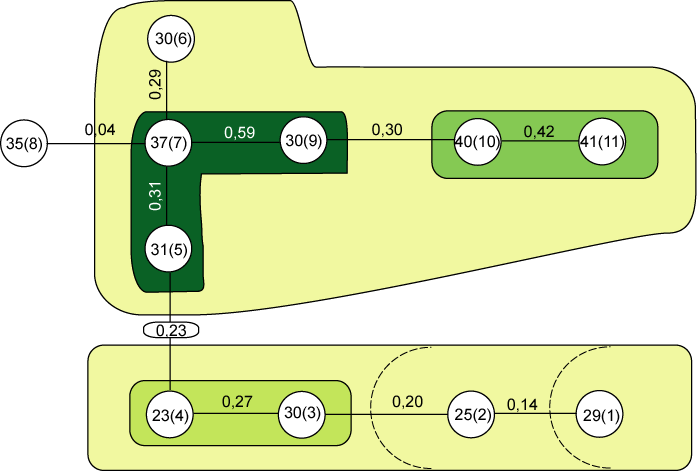
Рис. 1. Дендрит, построенный способом максимального корреляционного пути ключевого участка Хатипарского ландшафта (Оленикова, 2005)
0,27, 0,30…n – коэффициент сходства Жаккара
9(2), 23(4), 30(3)… n – номер урочища (номер пробной площадки)

Рис. 2. Дендрит, построенный способом максимального корреляционного пути ключевого участка Софийского ландшафта (Оленикова, 2005)
Р
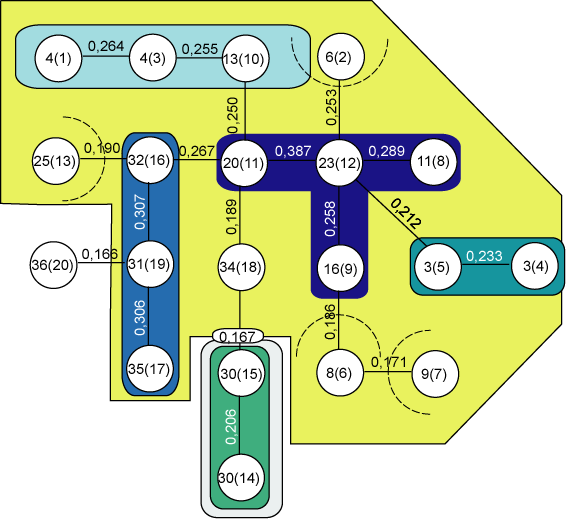
ис. 3. Дендрит, построенный способом максимального корреляционного пути Гондарайско-Узункольского ландшафта (Оленикова, 2005)
На уровне минимальной связи (0,04) в дендрите ключевого участка Хатипарского ландшафта (рис. 1) все урочища образуют общую корреляционную плеяду. При последовательном повышении уровня связи r в дендрите происходит отделение районов и выделение корреляционных плеяд более низких уровней. Первым от общей плеяды отделяются урочища альпийского пояса 35(8), где наблюдается большое видовое разнообразие. Затем следует расщепление общей плеяды на две: в первой последовательно отщепляется зона экотона верхней границы леса 29(1) и отделяется плеяда урочищ с субальпийским высокотравьем 25(2). Во второй - сначала отделяется плеяда урочищ субнивального пояса 40(10), 41(11) со специфическим растительным покровом, состоящим из видов, хорошо приспособленных к перепаду температур. Дальнейшее повышение уровня связи в дендрите приводит к выделению плеяды, состоящей из урочищ альпийского 30(9), 37(7) и субальпийского поясов 30(6), 31(5).
В дендрите ключевого участка высокогорного Софийского ландшафта при минимальной связи (рис. 2) первым отделяется урочище субнивального пояса 20(10) с наибольшей автономностью в растительном покрове. Затем образуются четыре плеяды, выделенные в два района. Первые две соответствуют урочищам, располагающимся в зоне экотона верхней границы леса 9б(3), 9а(4) и 9(2), 9а(5), 9а(1), причем уровень связи больше в плеяде урочищ, занимающих юго-восточную и восточную экспозиции. Дальше происходит отделение плеяд урочищ, расположенных в субальпийском 12(7), 14(6) и альпийском поясах 17(9), 19(8).
На уровне минимальной связи (0,166) в дендрите ключевого участка Гондарайско-Узункольского ландшафта (рис. 3) отделяется урочище субнивального пояса 36(20). При повышении уровня связи выделяются два района и плеяды урочищ с альпийскими коврами 32(16), 31(19), 35(17). Затем происходит отщепление урочища зоны экотона верхней границы леса 25(13), занимающего склоны восточной экспозиции, труднодоступные для животных. Дальше идет отщепление еще двух урочищ зоны экотона, но уже с явным нарушением состояния растительного покрова 6(2), 8(6). И последним отделяется урочище субальпийского пояса 9(7) с наиболее типичными субальпийскими лугами, находящимися в изолированном состоянии от прямого воздействия скота. Затем следует расщепление общей плеяды на плеяду урочищ, занимающих озерную четку 3(5), 3(4) и отличающихся большой антропогенной нарушенностью травянистого покрова. С последующим повышением уровня корреляционной связи выделяется плеяда урочищ субальпийского пояса западной экспозиции 4(1), 4(3), 13(10). Следующей отделяется плеяда урочищ альпийского пояса нижнего озера 30(15), 30(14). Последними выделяются урочища субальпийского пояса второго и третьего высокогорных цирков 20(11), 23(12), 11(8), 16(9).
При изучении растительных комплексов в рамках урочищ необходимо учитывать не только сходство фитоценозов, но и их автономность. С этой целью рассчитывался коэффициент доли автономных видов. Автономностью и формированием своей специфической растительности отличаются две переходные зоны: пояс экотона верхней границы леса и субнивальный переходный пояс у верхней границы ландшафта. Фактор природной разнородности приводит к дифференциации растительного покрова по экспозициям склона, а также приуроченности к более молодым моренным отложениям или более старым по возрасту коллювиальным отложениям на склонах. В результате на моренах найдена более автономная растительность, которая еще не обнаруживает тесную связь между собой, а лишь начинает образовывать разорванные сообщества. Растительный покров коллювиальных склонов более однороден по флористическому составу и отличается наиболее сформировавшимися сообществами.
Исследование травянистой растительности на ключевых участках позволило рассчитать опытным путем средний показатель коэффициента обилия видов на пробных площадках. Наибольшим видовым обилием отличается Гондарайско-Узункольский ландшафт, что во многом связано с разрастанием паскуальных сорняков за счет хорошо поедаемых скотом кормовых трав. К таким видам относятся опушенные растения: представители рода бурачок (Alyssum), некоторые желтушники (Erysimum), многие виды губоцветных, особенно такие, как чистец (Stachys), несъедобные для скота. Большое распространение вследствие своей пахучести и непоедаемости получают чебрец (Thymus), некоторые виды Ziziphora, Heracleum, Veratrum. На остальных участках ландшафта растительность развивается в естественных условиях и подвержена незначительному влиянию антропогенного фактора. В структуре растительных фитоценозов сорные виды растений подавлены или же постепенно вытесняются другими.
Для оценки антропогенной дигрессии ключевого участка Гондарайско-Узункольского ландшафта по видовому разнообразию травяного покрова зоны экотона верхней границы леса и субальпийского пояса применялся коэффициент степени дигрессии растительного покрова: Кс.д.= количество сорных видов/общее количество. Была принята следующая градация видов дигрессии: 0-0,1 – не нарушенный; 0,11-0,2 – слабо нарушенный; 0,21-0,3 – нарушенный; 0,31-0,5 – сильно нарушенный; более 0,5 – очень сильно нарушенный; 1- полностью разрушенный. В процессе обработки данных получены следующие показатели коэффициента степени антропогенной дигрессии. В поясе экотона верхней границы леса на склонах западной экспозиции данный коэффициент изменяется от 0,3 до 0,43. Склоны восточной экспозиции менее подвержены антропогенной трансформации, здесь коэффициент составляет 0,2. В местах водопоев и толок этот показатель увеличивается до 0,52. Выше, в субальпийском поясе, где идет непрерывный выпас скота, коэффициент на западных склонах цирка – 0,32, а на восточных изменяется от 0,3 до 0,4 в местах выгона. Увеличение абсолютной высоты над уровнем моря влечет за собой уменьшение показателя степени дигрессии от 0,27 до 0,15 в альпийском поясе. Таким образом, степень антропогенной дигрессии изменяется от слабонарушенного до очень сильно нарушенного травяного покрова в местах непосредственного ведения свободного пастбищного выпаса. В альпийском поясе дигрессия проявляется слабо.
В результате межвидового анализа растительного покрова урочищ выявлено, что литогенное поле разнородности, образующее различные формы мезорельефа высотных поясов ландшафта, определяет внутрипоясную дифференциацию растительного покрова в зависимости от местоположения (экспозиции и крутизны склона, характер горных пород и др.), и создает большую пестроту гидротермических «композиций» фитоценозов. В таких условиях в растительном покрове преобладают латеральные связи. Антропогенное вмешательство частично нарушает эти связи, усиливая автономность сообществ в пределах урочищ. Видовое разнообразие на ключевых участках ландшафтов во многом ограничивается сильным конкурентным воздействием со стороны основных эдификаторов и режимом частных нарушений (Олейникова, 2005).
При проведении регионального анализа растительного покрова можно сказать, что наименее автономна растительность Хатипарского ландшафта. Это во многом связано со сложившимися на протяжении многих лет тесных внутривидовых связей в рамках заповедности и ненарушенности естественных условий. Занимая центральное положение района исследования, ландшафт отличается более благоприятными климатическими условиями и достаточно однородным рельефом без резких перепадов высот.
Софийский ландшафт, занимая западный ключевой участок, отличается повышенным увлажнением и растительность здесь находится в стадии восстановления после антропогенных нагрузок. Формирующийся специфический растительный покров, отличается значительной долей автономности в двух зонах экотона – на границе леса и субнивальном поясе.
Гондарайско-Узункольский ландшафт, занимающий восточный ключевой участок, более автономен в растительном покрове по сравнению с остальными изучаемыми ландшафтами. Наиболее интенсивно здесь протекают процессы ксерофитизации, что отличает растительный покров по видовому составу от других ландшафтов. На автономность зоны экотона верхней границы леса оказывает нивелирующее воздействие выпас скота, в результате чего доля автономных видов здесь значительна, однако меньше чем в субальпийском поясе. Субальпийский пояс в свою очередь отличается разнообразным рельефом, представленным чередующимися высокогорными цирками. В результате формируется специфическая растительность, отличающаяся значительной автономностью. Альпийский пояс испытывает наименьшее антропогенное влияние, однако однородность в растительном покрове во многом зависит от дефицита тепла и короткого вегетационного периода. Субнивальный пояс отличается наибольшими отметками высоты над уровнем моря, которые сказываются на формировании специфических низкорослых сообществ растений приуроченных только к этому району местообитания.