
Dxm faq ответы на часто задаваемые вопросы о декстрометорфане (dxm)
| Вид материала | Документы |
- Ответы на вопросы, 90.99kb.
- Налоговая инспекция информирует, 47.91kb.
- Есн и страховые взносы (внебюджетные фонды). Ответы на наиболее часто задаваемые налогоплательщиками, 58.71kb.
- Отчетными периодами признаются первый квартал, полугодие и девять месяцев календарного, 78.47kb.
- 182 1 03 01000 01 0000 110 Вопросы исчисления и уплаты ндс, в том числе оформления, 312.98kb.
- 3 Приборы EcoQuest. Ответы на часто задаваемые вопросы. Вопросов больше не будет, 491.44kb.
- Ответы на часто задаваемые вопросы, 53.91kb.
- 2 наиболее часто задаваемые вопросы, 856.03kb.
- Содержание Часто задаваемые вопросы, 2264.71kb.
- Программа похудения и оздоровления Наиболее часто задаваемые вопросы Пускай Пища станет, 3239.1kb.
9.5. Фармакокинетика: Как метаболизируется DXM
DXM, как соль гидробромида, быстро абсорбируется из желудочно-кишечного тракта; в течение 30 минут он может войти в кроветок. Соединение полистирекса предназначено для продолжительной абсорбции, и его полный переход в кровь может занять 6-8 часов.
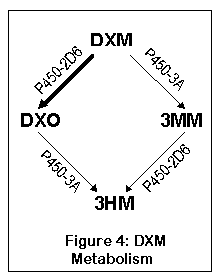
DXM метаболизируется двумя путями, оба из них приводят к одному - 3-гидроксиморфинану (3ГМ, 3HM). Первый путь исходит из DXM к DXO (декстрорфан), а затем к 3HM; второй из DXM к 3-метоксиморфинану (3ММ), а затем к 3HM. Большая часть DXM (до 90%) метаболизуется через DXO у нормальных индивидуумов.
DXM преобразуется в DXO печеночным ферментом, называемым цитохром P450-2D6 (дебрисохин 4-гидроксилаза, debrisoquine 4-hydroxylase). Примерно у 7% белых и 0,5% азиатов версия этого фермента работает крайне (до 70 раз медленнее) неэффективно и не может метаболизировать DXM в DXO должным образом. От 0.5 до 2% популяции имеют многочисленные копии гена P450, что приводит к очень быстрому метаболизму DXM в DXO (155). После превращения в DXO, ферменты P450-3A4 и P450-3A5 перобразовывают DXO в 3-гидроксиморфинан.
Другой путь ведет сначала к 3-метоксиморфинану (посредством ферментов P450-3A4 и P450-3A5), а уже затем к 3-гидроксиморфинану. Большинство людей не метаболизуют основное количество DXM таким образом, хотя люди с недостатком нормального P450-2D6 будут превращать существенную часть DXM в 3ММ. А 3ММ вероятно не психоактивен, что означает - 5-10% людей с недостатком фермента 2D6 ощутят меньший эффект от DXM (или более точно, не испытают эффектов DXO).
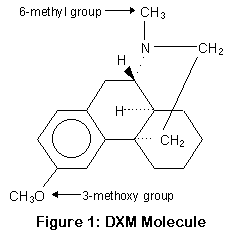
P450-2D6 работает, убирая метокси группу и заменяя ее гидрооксилом (OH) (или точнее, отрезая метил от кислорода); этот процесс называется О-деметиляцией. P450-3A4 и 3A5 заменяют метильную группу водородом (H); это процесс N-деметиляции. Положение метильной и метокси групп показано на рисунке.
9.5.1. Факторы, влияющие на метаболизм DXM
Как говорилось ранее, у некоторых людей отсутствует нормальный фермент P450-2D6. У остальной популяции этот фермент может быть ингибирован несколькими факторами. Многие лекарства ингибируют P450-2D6, особенно флюокситин (ProzacTM). Неполный список лекарств, ингибирующих фермент P450-2D6, дан в Разделе 15.1.
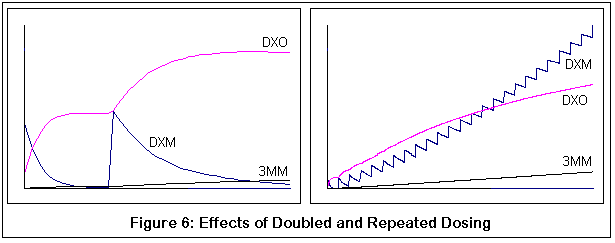
Сам по себе DXM, разумеется, будет конкурировать с другими веществами за P450-2D6 и, что важно, это будет делать и 3-метоксиморфинан (3ММ) (17). Фактически, 3ММ имеет большее сродство к ферменту P450-2D6, чем DXM. Это может объяснить тот факт, что вторая доза DXM "в догонку" производит эффект отличный от первой. Конкуренция за фермент P450-2D6 уменьшит количество DXM, превращающегося в DXO у второй дозы.
Этот график получен путем компьютерной симуляции метаболизма DXM:
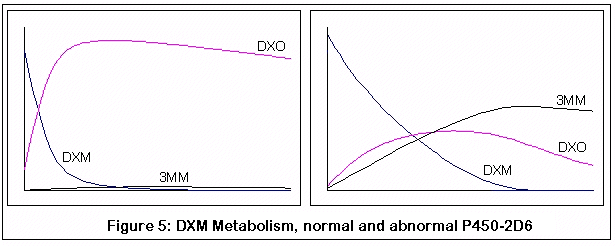
Первая пара графиков представляет метаболизм DXM у нормального индивидуума (слева) и индивидуума без нормального фермента P450-2D6 (справа). Заметьте быстрое и почти полное превращение DXM в DXO у нормального индивидуума, по сравнению с менее эффективным и более медленным превращением у человека с недостатком P450-2D6.
Вторая пара демонстрирует возможные результаты приема дополнительных доз ("в догонку"). Оба графика соответствуют нормальному варианту фермента P450-2D6. Отмечу, что вторая доза DXM не превращается в DXO также быстро (поэтому меньший наклон). Правый график показывает многочисленные дозы, и выгибание вверх метаболитической кривой DXM становится все более явным с каждой дозой.
Между прочим, это все качественные симуляции, а не количественные. Я пытался придерживаться известных значений КМ и VMAX для подходящих реакций [?], но симуляции были дискретным процессом (если честно, мои познания в уравнениях довольно слабы и требуют от меня титанических усилий). Я сравнил мои результаты с обнаруженными скудными данными, сравнения оказались разумными, но, опять же, я могу быть абсолютно неправым. Цель этих графиков - продемонстрировать относительные эффекты изменений в ферментной активности (из-за генетических изменений и конкурентного ингибирования 3-метоксиморфинаном), надеюсь, что этого оказалось достаточно.
У меня нет информации о том, что случается с самим 3-гидроксморфинаном. Он может выделяться напрямую почками, или подлежит дальнейшему метаболизму.
10. Нейрофармакология DXM
10.1. А что такое рецептор? (основы нейрофармакологии)
10.1.1. Структура нервной клетки
У всех клеток существует градиент напряжения между внешней средой и внутренним содержимым клетки; если бы вы измерили это напряжение, то обнаружили бы, что потенциал внутри клетки чуть больше –100 мВ по сравнению с внешней средой. В большинстве клеток этот заряд - следствие переноса ионов в и из клетки, и ни для чего такого не используется. Тем не мене, мускульные и нервные клетки нашли ему применение.
У мускульных и нервных клеток (нейронов) есть "возбудимые" мембраны - заряд мембраны может измениться, а это изменение применяется в специальных целях. В нейронах понижение этого заряда используется для передачи сигнала.
В мозге человека находится 10 миллиардов нервных клеток, и все они делают одну и туже работу: передают сигналы со своих входов (дендритов) на выходы (аксоны). Нейрон немного похож на дерево, только вместо корней и веток у него есть дендриты и аксоны. Грубо говоря, дендриты принимают сигналы, а аксоны - передают (помните, сигнал обозначается изменением потенциала клеточной мембраны).
В конце каждой веточки аксона находится синаптическое утолщение, маленькая, похожая на бутон структура, используемая для посылки сигнала к следующей нервной клетке [не всегда нервной] (об этом позже). Дендриты, наоборот, принимают сигналы и состоят из многих маленьких шишечек, каждая из них принимает сигнал от другого нейрона. Один нейрон может принимать сигналы от тысяч других нейронов.
Некоторые сигналы могут возбуждать мембрану клетки; другие - тормозить возбуждение. Если сумма возбуждающих сигналов минус сумма тормозящих сигналов, полученных дендритами, достаточно велика, то сигнал достигает тела клетки (сомы). Как только достаточно сильный сигнал достигает сомы, он тут же посылается по аксонам и передается дальше.
Это основа индивидуального функционирования нейронов. Тем не менее, это не объясняет, как один нейрон посылает сигнал другому (или, наоборот, как они принимают сигналы). Очень небольшое количество нейронов соединяются друг с другом на физическом уровне, и сигналы просто проходят из клетки в клетку как сигналы от одного транзистора к другому в микросхеме. Однако, большинство нейронов не касаются друг друга; вместо этого, они взаимодействуют с химическими веществами, называемыми нейротрансмиттерами [(нейро)медиаторами] и принимают их с помощью структур, называемых нейрорецепторами.