
Академия протагора в. А. Ивашко принципы эволюции человека – человечества
| Вид материала | Книга |
СодержаниеГармонизатором выступил нуклеопротеид – рибосома. |
- Происхождение человека, 109.18kb.
- Окружной методический центр восточного окружного управления образования лаборатория, 1613.77kb.
- Глобализация: понятие, этапы, противоречия, оценки, 325.76kb.
- Некоторые особенности сознания, 227.68kb.
- В. Н. учитель высшей категории, отличник просвещения РФ. 19 февраля 2010 года Задачи, 251.23kb.
- Методическое пособие к курсу "теория эволюции" Научный центр эволюции земли и человека, 4157.76kb.
- Методическое пособие к курсу "теория эволюции" Научный центр эволюции земли и человека, 4157.69kb.
- Эволюция человека, 22.68kb.
- Программы, залаженные в молекуле, 11344.58kb.
- Программы, залаженные в молекуле, 11345kb.
и АТФ – главный аккумулятор энергии любой клетки,
и еще 10 млн. самых разнообразных органических соединений как естественных, так и искусственных.
Белки, жиры, углеводы, ДНК – РНК и АТФ мы называем биогенами. Базовыми составляющими биогенов есть органогены: углерод, водород, азот и кислород, которые в сумме составляют почти 99% массы человеческого тела. На все остальные 20 химических элементов, входящих постоянно в метаболические процессы, проходящие в нашем теле, приходится всего 1%-1,5%. Здесь важна не столько точность цифр (у разных авторов они значительно разнятся) сколько неизменность безусловной доминации 4 органогенов. Все четыре органогена относятся к десятке самых легких элементов. Три из них являются активными участниками реакции горения, а азот – ее активным противником.
Теперь обратим внимание на стык атмосферы, гидросферы и литосферы. Доминантами являются:
в атмосфере азот 75%, кислород 23%
в гидросфере водород 11%, кислород 89%
в литосфере кремний 29%, кислород 49%
в биосфере водород 50%, кислород 25%
Формально доминации углерода нет даже в биосфере, но это из-за того, что все бионты более чем наполовину состоят из воды. Так тело человека включает:
вода – 65%
белки – 19%
жиры – 12%
углеводы – 1%
другие
(
 в виде золы) – 3%.
в виде золы) – 3%.итого – 100%
Отсюда ясно, почему в биосфере углерод составляет 18% по массе, а в теле человека – 11%. В то же время в атмосфере, гидросфере, литосфере вместе взятых углерод не занимает и 0,1%. Следовательно, доминация углерода в биосфере беспрецедентно неопровержима.
Однако из этих сравнений четко видно и другое. Литосфера представлена кристаллической формой SO2, гидросфера – водой Н2О, а биосфера удивительной коллоидной формой.
Еще в середине ХIХ в. английский химик Т. Грэм предложил разделить все вещества на коллоиды и кристаллоиды. К коллоидам отнесли дисперсные системы, т.е. растворы с взвешенными частицами:
твердыми – суспензия
жидкими – эмульсия,
в свою очередь эмульсии разделили на гидрофильные (мыло) и гидрофобные (жиры). Однако уже в 1869 г. профессор Киевского университета И. Г. Борщов доказал, что возможно существование одного и того же вещества как в коллоидной, так и в кристаллоидной формах. О твердо- , жидко- и газокристаллических формах вещества мы уже говорили. Следовательно, и коллоидная и жидкокристаллическая форма вполне могут совместиться в определенных условиях. Несомненно, что жидкокристаллическая форма хронотопно уже является коллоидом, но вот эволюционность коллоида в жидкокристаллическую форму далеко не так очевидна.
Споры о том, почему углерод, а не стоящий сразу ниже него в таблице Менделеева кремний стал органогеном беспредметны по одной простой причине: энергия связи Si – Si в 2 раза меньше, чем у С – С, поэтому кремниевые цепочки ограничены максимум 8 атомами (Si8H16) и кремний неспособен в отличие от углерода образовывать двойные и тройные связи. Так что в атоморганизационном отношении углерод – безусловный лидер органической природы. К тому же поразительна изомерность углерода: если 8 атомов углерода в октане (С8Н18) дают 18 различных вариантов их расположения в цепи, то соединение с 40 атомами углерода уже могут дать 60 триллионов изомеров. Все живое на Земле построено всего из 20 разных остатков аминокислот, которые могут составить 2 432 902 008 178 640 000 изомеров. И это при том, что аминокислот уже имеются полторы сотни и открываются новые.
По традиции белок считается биогеном №1. В 1836 г. Г. Мулдер открыл, что основу всех белков составляет протеин (C40H62N10O12). В начале ХХ в. Э. Фишер взялся за анализ и синтез белка. И установил, что белок последовательно расщепляется на пептоны, которые далее расщепляются на пептиды, которые далее расщепляются на аминокислоты. А в 1907 г. он осуществил и первый синтез полипептида из 18 аминокислотных остатков, т.е. доказал возможность не только развертки, но и свертки белка в принципе.
Глицин – одна из простейших аминокислот – это производное уксусной кислоты, в которой один из атомов водорода заменен на остаток аммиака (группа – NH2). В этом элементарном школьном примере четко виден переход неорганического в органическое.
Итак, белки – это высокомолекулярные органические соединения, состоящие из остатков аминокислот с молекулярной массой от 6,5 тыс. до 32 млн. Уже известно много сотен белков и полторы сотни составляющих их аминокислот. Но все бионты состоят всего лишь из 20 аминокислот, которые при полимеризации отдают предпочтение какой-либо другой. И эта выборочность, предпочтимость - сочетаемость есть один из главных факторов того, что из 150 известных уже аминокислот основу жизни составляют только 20 дружественных (другие появляются лишь изредка в специфических условиях у низших животных), которые и дают в клетках высших животных и человека 3000 различных белков. Все белки в бионтах имеют спиральную форму (Л. Полинг) и левонаправленны (Л. Пастер).
Жиры химики относят к группе липидов и липоидов, которые представляют собой сложные эфиры глицерина и карбоновых кислот. Нам здесь важны только жирные кислоты, ибо в организме пищевые жиры распадаются на глицерин и жирные кислоты. Животные жиры содержат пальмитиновую и стеариновую кислоты, а растительные жиры (масла) содержат олеиновую и линолевую кислоты. Уже известно почти полторы тысячи различных жиров и масел в природе. Нерастворимость жиров в воде и стала причиной того, что они стали основой мембраны (оболочки) клетки. А в процессе внутриорганизменной пересинтезации (ресинтезе) участвует фосфорная кислота и эти фосфатиды становятся еще и аккумуляторами энергии. Но все абиогенные жирные кислоты имеют равное количество четности и нечетности атомов углерода, а жирные кислоты бионтов имеют только четное количество углеродных атомов. К липоидам по причине их жирорастворимости химики отнесли и витамины А, Д, Е, К.
Первым в 1854 г. синтезировал жиры Марселен Бертло. И сейчас их синтезируют в огромных количествах в технических целях. Синтез пищевых жиров и масел до сих пор остается проблемой, но вовсе не технической, а медико-биологической: не навредить бы.
Углеводы химики называют сахаридами. Базовый сахарид – глюкоза (С6Н12О6) дает много изомеров и более сложных соединений, которые при разложении все приходят к исходной форме глюкозы. Синтез глюкозы из СО2 и Н2О за счет энергии Солнца – это уникальное запасание энергии впрок путем фотосинтетического свертывания. Далее у глюкозы два пути:
1. служить исходным аккумулятором энергии и выделять ее по мере надобности путем окисления;
2. служить исходным строительным материалом для построения молекул большинства других необходимых организму соединений, т.е. еще более упаковываться энергетически. Именно поэтому в растениях сахариды составляет 50% древесины.
Иная картина у животных. Чтобы стать аккумулятором энергии, сахаридам надо было преобразоваться в нуклеотиды, которые включают:
- азотистые основания:
а) производные пиримидина:
цитозин, тимин, урацил;
б) производные пурина:
аденин, гуанин;
- сахара – пентозы:
рибоза, дезоксирибоза;
- фосфаты.
Самым главным из базовых хранителей энергии стала аденозинтрифосфорная кислота АТФ. АТФ – это комплекс аденина, рибозы и трех остатков фосфорной кислоты, которые могут последовательно отщепляться (АТФ↑→АДФ↑→АМФ) освобождая каждый раз 11000 калорий, необходимых организму. Эта разрядка аккумулятора внутренней химической энергии восполняется путем окисления глюкозы, освобожденная энергия которой идет на дозарядку. Таким образом синтез АТФ у растений и животных качественно различен: собственно упаковыванием энергии Солнца занимаются растения, а животные лишь преобразуют ее, точнее паразитируют за счет растений.
Следует напомнить здесь и о реакции брожения, т.е. разложения той же самой глюкозы не путем окисления, а под действием определенных ферментов. Специалисты утверждают, что окисление энергетически эффективнее брожения в 10-50 раз как по скорости, так и количественно.
В 1929 г. К. Леман открыл АТФ, выделив ее из живой ткани. В 1948 г. А. Тодд синтезировал АТФ. Но до полного эволюционного понимания энергетической сущности бионта еще очень далеко.
Нуклеиновые кислоты были открыты в 1871 г. Ф. Мишером Правда сам автор назвал их нуклеином. Переименовал нуклеин в нуклеиновые кислоты в 1889 г. ученик Ф. Мишера Р. Альтман. Много позднее оказалось, что и это название не совсем точное, ибо собственно нуклеиновой (nucleus - ядро) является только ДНК, а РНК находится в цитоплазме. Но уже так свыклись с названием РНК что решили оставить все как есть, хотя и гносеологически ущербно. В 1944 г. О. Эвери доказал экспериментально, что ДНК и гены – это одно и то же. Сформировался первый стык/шов органической химии, биохимии и генетики. В 1953 г.о Ф. Крик и Д. Уотсон создали первую модель ДНК в виде двойной спирали. Ударная волна этого открытия перевернула не только мир научный, но и цивилизованное общественное сознание. Мир вперился и впился в дотоле неизвестные даже большинству ученых аббревиатуры ДНК – РНК. Гигантски расширился фронт работ в этой сфере. В 1968 г. А. Корнберг синтезировал в пробирке первый активный вирус. В 1972 г. Пол Берг впервые ввел искусственные гибридные молекулы ДНК в клетку – хозяина и изменил ее генотип. Родилась генная инженерия. В 1973 г. уже был создан ген для синтеза гемоглобина. И т.д., и т.п. Едва Сэнгероле, Гилберт и Макэм разработали метод определения последовательности нуклеотидов ДНК, что теоретически давало возможность привязать функциональные свойства генома к определенным структурам, как началась эпохальная работа над расшифровкой генома человека. Теперь, видимо, мучаются над вопросом, как бы все добытое подороже продать. Ведь вся высокая философско-педагогическая евгеника оказалась всего лишь специфической рекомбинационной структурой из пусть и высокомолекулярных, но все-таки тривиально химических соединений. А мир застыл в недоумении, то ли радоваться сему, то ли плакать. Однако вернемся к прозе химического мира на его границе с миром биологическим.
Итак, ДНК – это полимер сахаров дезоксирибозы, соединенной фосфатными связями с азотистыми основаниями: пуринами (аденин, гуанин) и пиримидинами (цитозин, тимин), которые образуют двойную спирать так, что аденин обязательно связан с тимином, а гуанин с цитозином.
РНК – это модификат ДНК, т.е. полимер сахаров рибозы, соединенной фосфатными связями с азотистыми основаниями: пуринами (аденин, гуанин) и пиримидинами (цитозин, урацил).
Родство ДНК и РНК очевидно, они отличаются только сахаром – пентозой (дезоксирибоза и рибоза) и одним из пиримидиновых оснований (тимин и урацил). Но если в ДНК тимин связан строго с аденином, то в РНК уже урацил строго связан с аденином. Любопытно, что именно аденин, связанный с рибозой в аденозин, и тремя остатками фосфорной кислоты образует АТФ. Сей факт явно указывает на какие-то эволюционные основания. Ведь и главенство ДНК – это только в полноценной ядерной клетке. Специалисты предполагают не без оснований что эволюционно первой была РНК. Значит не РНК есть модификат ДНК, а ДНК есть модификат РНК, где рибоза потеряла одну оксигруппу (–ОН) и стала дезоксирибозой, а урацил заменил тимин, т.е. тот же урацил но с метильной добавкой (–СН3). К тому же РНК много короче ДНК и, главное, имеют не двойную спираль, а одиночную, но может образовывать и двойную. Наконец, оказалось, что просто РНК не существует: имеются информационные РНК, транспортные РНК и рибосомные РНК.
При этом транспортных РНК ровно столько, сколько аминокислот, т.е. 20. Значит каждая транспортная РНК должна не только узнавать свою аминокислоту, но и последовательность ее поставки к рибосоме. Такая себе логистика на биомолекулярном заводе, производящем разнообразные белки-блоки из одних и тех же 20 аминокислот. Думал ли А. М. Бутлеров в 1861 г., когда впервые синтезировал сахар из обыкновенного ядовитого формальдегида, что сахара-сахариды окажутся в самом центре скандальной проблемы сущности жизни. А ведь вся эта хитроумнейшая логистика еще далеко не жизнь, а обычные последовательно-цепные реакции органохимического мира в специфических физических условиях. РНК в зависимости от силы ионных связей и температуры раствора могут существовать в виде нитей, палочек, клубков, а вот ДНК в силу своей великости предпочитает только тугоклубочный тип существования. И чем длина молекулы ДНК больше, тем клубок туже. Сама длина обусловлена числом включенных нуклеотидов. Так, у бактерии их 2 млн. пар, у знаменитой мушки дрозофилы 200 млн. пар, а у человека 3 млрд. пар. Грубо можно подсчитать, что якобы дрозофила в 100 раз сложнее устроена, чем бактерия, а человек всего лишь в 15 раз сложнее, чем дрозофила и в 1500 раз, чем бактерия, тогда как по длине молекул ДНК все эти цифры больше, т.е. 15 и 15000.
Понятно, что эта математика как-то связана с качественными соотношениями, но явно не столь грубо, ибо по сути мы понятия не имеем, а что собственно считаем: подсистемы систем или функции, которые к тому же могут быть разными в зависимости от структуроположенности.
Итак, в мире химическом избирательность взаимодействий проявляется еще четче и глубже по иерархии:
мир гравитационный,
мир электромагнитный,
мир вещественный:
- минеральных соединений,
- углеродных соединений,
- биохимических соединений.
Вроде бы вполне системно-логично, но только если смотреть на иерархию хронотопно, ибо так есть сейчас. Но эволюционно имеет место быть гносеогическая заноза логики перескока углеродных соединений в биохимические с их уникальным генеалогическим статусом. Ведь именно на этом стыке должен был возникнуть качественно иной от хемореактора биореактор, уникально низкоэнергетический.
Мир биогенный
При анализе мира биогенного строго системно-логически неизбежно упираемся в триединство проблемы жизни:
- гносеологическую: что такое жизнь?, которую должна изучать виталогия;
- онтологическую: проблема реального геономического биогенеза;
- технолого-теоретическую: моделирование живого, т.е. бионта.
И сваливать их в кучу, как это обычно делается и философами-методологами и биологами-методологами, совершенно недопустимо.
Виталогия – сугубо философская дисциплина о сущности жизни и возможностей ее существования в иной, чем биологическая, форме, что совершенно не противоречит принципам философии. Я по крайней мере не нашел ни одного системно-логического запрета на вероятность ее существования. А на шутки стохастики надо относиться с пониманием: не обнаружили до сих пор, то можем обнаружить в будущем. Ведь и теоретическую вероятность появления жизни на Земле специалисти оценивают далеко не оптимистически: 10-255 по Г. Кастлеру или 10-800 по Л.А. Блюменфельду. А она-то есть уже более 4 млрд. лет. И тот факт, что вы сейчас читаете эту книгу, говорит о том, что земная жизнь сумела развиться до такой сверхсложной системы как Человек-Человечество.
Вторая проблема виталогии – наш неизбывный антропоморфизм, породивший вечных, но человекоподобных богов, а от них и богоподобных людей с бесподобными гносеологически-мифологическими (культурологическими) и чисто литературно-фантастическими вариациями.
Третья проблема виталогии напрямую связана и вытекает из идеи испоконвечных богов, но в более наукообразной форме – идея панспермии, ведущая свое начало от Анаксагора.
Четвертая проблема виталогии была поставлена уже самой гносеологией: в чем сущность живого? А так как у нас под рукой ничего кроме биологической формы жизни нет, то и ответить на этот вопрос можно только отталкиваясь от ее антипода, т.е. неживого. А это весьма и весьма непросто, в чем вы сами убедитесь в ходе дальнейшего анализа, ибо придется научиться различать продукт углеродных соединений и продукт биохимический, т.е. косное и живое, неживое, мертвое (системно-функциональное, системное, но не функциональное, несистемное и не функциональное).
Итак, виталогия в основе своей пока что целиком и полностью зависит от результатов максимально возможно строгого системно-логического анализа именно мира биогенного, т.е. того, что понимается как теоретическая биология. Но беда в том, что теоретической биологии пока что не существует. Есть отдельные наметки, подходы, но они базируются на абсолютно разных основаниях и уже поэтому не могут интегрироваться, совместиться в цельносистемное обозрение.
Именно поэтому проблема биогенеза фактически решается без какого-либо действенного участия биологов-теоретиков биохимиками и биофизиками с редкими набегами других специалистов, иногда весьма далеких от понимания сути самой проблемы. Я не говорю уже о том ущербе, который наносят и ученым-исследователям и широкой общественности скандально-шумные малограмотные масс-медийные спекуляции по этой теме. Да и в самой проблеме наметился гносеологически опасный разрыв между теоретической реконструкцией биогенеза и его физико-химическим (по сути технологическим) моделированием с доминацией последнего. В первом случае необходимо реставрировать саму систему, породившую жизнь, а во втором – сотворить копию уже существующего продукта, не суть важно какими методами. Боюсь, даже нанотехнологии не объединят эти два направления, ибо задача первого – объяснить процесс биогенеза, а второго – его результат. И как было показано в предыдущей главе успехи моделирования результатов угрожающе велики. И пока что невозможно сказать, что здесь есть подгонка под результат, а что соответствует логике реального биогенеза. Свою роль сыграло и то, что моделирование результатов фактически ведет свое начало со случайного открытия Ф. Вёлера, а моделирование процесса биогенеза началось лишь сто лет спустя со знаменитого доклада А.И. Опарина 3 мая 1924г.
Биогенез
Строго системно-логически проблема биогенеза предполагает решение трех взаимопереплетенных проблем:
- геономии,
- геохемогении,
- цитогении.
Геономия в рамках теории образования Солнечной системы обязана прежде всего дать строго системно-логический анализ формирования и эволюционного взаимодействия геономического триединства: Солнце-Земля-Луна, которое и предопределило сам геобиогенез, предопределяло, предопределяет и будет предопределять эволюцию всего живого на Земле, включая и Человека-Человечество при всем его технотехнологическом современном самомнении. Однако ответить на вопросы: где? когда? как? и почему? образовалась гелиосистема и геономическая триада в ней, уже никогда не удастся сколько-нибудь доказательно, ибо для этого нужна не просто теоретическая модель образования Метагалактики (хотя бы и в виде Большого взрыва), а много более прозаическая и точная карта ее, восстановить которую просто невозможно ни ретроспективно, ни ретроперспективно в силу непрерывной изменяемости всего и вся. К тому же мы понятия не имеем о возрасте гелиосистемы, что окончательно рушит попытки определения былого хронотопа даже на контурной карте Метагалактики. Имеющиеся в литературе ссылки на якобы достаточно точные данные о возрасте Солнца вовсе не говорят что-либо внятное о возрасте самой гелиосистемы, т.е. о строгости взаимосвязи формирования звезд и их планетных систем. Маргинальность Солнечной системы обусловила огромные трудности в формировании планетологии, ибо планет как таковых и тем более планетных систем типа нашей оказывается весьма и весьма незначительное количество в сравнении со сгустками плазмы звезд. Еще большую путаницу вносит наличие гигантских “газопылевых” образований, природа которых еще более темна. Единственное, что уже вроде бы общепризнано, это четкая спиралевидность галактик как тенденция. Однако, что именно выражает эта спиралевидность: момент закручивания или момент раскручивания, определять надо в каждом конкретном случае отдельно.
На базе этих весьма зыбких представлений и делаются попытки выдвижения гипотез о горячем и холодном происхождении Солнечной системы. Но никто не анализирует при этом качественное различие физико-энергетического и химико-атомарного миров. Мы ведь по сути понятия не имеем о структуре иных планет кроме Солнечной системы. Более того, мы не имеем даже четкого определения планеты (история с Плутоном тому яркое подтверждение). Шарообразность – первый признак планеты – явно указывает на спиралевидный тип ее образования, т.е. более согласуется с холодной, газопылевой теорией их происхождения. Отсюда и обязательное вращение планеты вокруг своей оси как продолжение спиралевидного закручивания ядерного вещественного сгустка в момент происхождения. Но остается вопросом, мог ли быть/стать ядром такого образования энергоплазменный сгусток, использовавший принцип снежного кома? Этот принцип работает в обоих случаях спиралевидного происхождения. Однако из него четко и строго системно-логически должно следовать, что масса планет такого типа должна непрерывно расти как функция гравитации самой себя по законам небесной механики Ньютона. Прекращение осевого вращения переводит планету из активной формы в пассивную, а значит и зависимую во много большей степени. Луна скорее всего и является такой планетой. И строго системно-логически должна терять свою массу в геологических масштабах времени. Однако остается темной проблема схода Луны со своей орбиты и ее захват Землей и т.д. и т.п. Явный диссонанс в идиллию гипотезы холодного происхождения Солнечной системы вносят само Солнце и каверзно неправильное направление вращения Венеры и Урана вокруг своих осей, прямо противоположное относительно остальных планет гелиосистемы. Таким образом, все указывает пока что на уникальность Солнечной системы как по ее современному положению в Метагалактике, так и по способу образования (скорее всего смешанному). Для нас же в данном случае это означает отсутствие сколько-нибудь твердой гносеологической опоры на геономическую триаду при анализе проблемы биогенеза. Отсюда и неизбежность использования принципа актуализма, получившего распространение сначала в геологии, а затем и в других науках.
Значит нам ничего не остается как только, хотя бы в общих чертах, определить роль геономических факторов.
Земля обеспечивает:
- вращение вокруг своей оси с запада на восток с некоторой скоростью, эволюционные параметры которой пока плохо изучены;
- гравитацию некоторой силы с темной эволюционной изменяемостью;
- некоторый наклон оси вращения относительно Луны и Солнца, эволюционная изменяемость которого (наклона) далеко не ясна;
- магматическую деятельность некоторого интрапланетарного реактора с периодическими слабо предсказуемыми вулканическими образованиями/извержениями и землетрясениями;
- дрейф коры, обусловленный плаванием на магме с далеко не ясными эволюционными характеристиками;
- уникальное для космических тел сохранение по длительности триединства поверхностного стыка лито – , гидро – и атмо – сфер, а значит и единоцельности планеты в целом.
Луна обеспечивает относительно Земли:
- гравитацию некоторой силы с эволюционно темной изменяемостью, но четкой актуально приливно-отливной ритмикой земной коры от магматического основания до стратосферы;
- отражение солнечного света.
Солнце обеспечивает относительно Земли:
- гравитацию некоторой силы с эволюционно темной изменяемостью;
- в новолунья и полнолунья усиливает приливно-отливную ритмику магмы, земной коры и т.д., со всеми вытекающими последствиями;
- мощное электромагнитное облучение, обуславливающее непрерывный кругооборот веществ и энергии на поверхности планеты и прилегающих сферах в их бесчисленных и многообразных превращениях и взаимодействиях, однако эволюционно характеристики этого облучения пока что изучены слабо.
Понятно, что при таком количестве темных мест говорить о каком-либо ретроперспективном прогнозе не приходится. Меня спасает только сам факт существования жизни на Земле. Другое дело: а земная ли она по сути?
Геохемогения должна бы изучаться геохимиками, но геохимия сама еще осень молода, а фронт работ огромный. Тем не менее успехи теоретической геохимии очевидны. Иное дело, что до карбонэргической (органической) геохимии у них еще руки не дошли. Да и чисто методологические трудности различения минералогенности и биогенности требуют своего принципиально четкого разрешения, что в условиях уже существования биосферы далеко не так просто.
Тот факт, что некоторые карбонэргические соединения обнаружены и далеко за пределами Земли, говорит о том, что процесс свертывания химических элементов во все более упакованные структуры носит в химическом мире всеобщий характер. Загвоздка только в том, что там нет четырехвалентного углерода, дающего возможность создавать как простенькие мономеры, так и поистине гигантские полимеры. Но именно и только этот четырехвалентный углерод присутствует в биохимических структурах:
а) моносахаридах,
б) аминокислотах,
в) нуклеотидах.
Значит именно геономический хронотоп позволил за какой-то геологически короткий промежуток времени проявить именно четырехвалентному углероду свои гениальные организаторские способности. Таким образом, под геохемогенезом мы здесь понимаем не собственно хемогенез, ибо Земля уже была химическим телом а именно карбонэргический хемогенез из того, что уже было: водорода, кислорода, азота, фосфора, кальция и т.д. как поотдельно, так и через соединения метана, углекислого газа, воды, аммиака, формальдегидов и т.д.
Моносахариди Cn H2n On ведут свое начало из чисто минеральной уксусной кислоты (С2 Н4 О2), но обрели иную стереохимическую структуру, а став изомерами, обрели и новые качественно отличные от своей прародительницы свойства. Реагируя между собой, они стали образовывать цепочки углеводов. Главное место среди них несомненно принадлежит глюкозе (С6 Н12 О6), которая в форме крахмала или гликогена накапливаются в организме бионта и служит ему по мере необходимости едва ли не единственным топливом при производстве энергии в процессе непрерывной жизнедеятельности.
Аминокислоты – это все та же уксусная кислота, в основе которой один/два атома водорода заменены на остаток аммиака (-NH2). И в результате получаем абсурдный с минеральных позиций продукт – гермафродит, ибо в одной молекуле имеем два антагонистических элемента: группа -NH2 есть носитель основных (щелочных) свойств, а группа –СООН есть носитель кислотных свойств. Отсюда и идет гигантская сочетаемость аминокислотных остатков между собой до сотен тысяч в одной белковой молекуле. При этом энерговещественная свертываемость идет строго по линии: аминокислоты→пептиды→пептоны→белки. В свого очерель белки достаточно легко образуют сложные белки: нуклеопротеиды, хромопротеиды, металлопротеиды и т.д. Это и обусловило центральную роль белков в биогенезе. Ряду белков присущ автокатализ, а сократительные возможности некоторых белков легли в основу активной пульсирующее – двигательной, а на ее основе и в дополнение еще и чувствительно – двигательной функции бионтов. О доминации левосторонней хиральности у белков мы уже говорили. А вот о доминации 20 аминокислот из 200 уже известных следует сказать особо, но не с биохимических, а с гносеологических позиций. Школярская абсолютизация двадцатки одних и тех же аминокислот во всем живом не соответствует действительности, ибо не было одноактности творения живого, а был мучительно долгий процесс его эволюции, который продолжается и сейчас. И отбор на сочетаемость/выживаемость аминокислотных остатков в белках особенно нагляден для низших форм организации живого, прежде всего одноклеточных. И здесь биохимики обнаруживали, обнаруживают и еще будут обнаруживать множество сочетаний вовсе не характерных для канонической двадцатки, характерной для человека. Дидактические упрощения не должны смазывать сам процесс эволюции живого. А она (эволюция) неизбежно идет, ибо процесс изменяемости всего и вся непрерывен. И никакие догматики от биологии не в силах его остановить.
Нуклеотиды – это весьма гармоничное и многофункциональное сочетание трех составляющих:
Азотистое основание: + сахара-пентозы: + фосфаты:
- аденин, - рибоза - моно –
- гуанин, - дезоксирибоза - ди –
- тимин, - три –
- урацил,
- цитозин,
В нуклеиновых кислотах тон задают сахара-пентозы, поэтому и кислоты называются по ним:
рибонуклеиновая, (РНК)
дезоксирибонуклеиновая (ДНК),
и правая хиральность РНК и ДНК обусловлена ими, и азотистые основания выстраиваются строго по их указаниям:
пурины: пиримидины:
РНК: аденин – урацил
гуанин – цитозин
ДНК: аденин – тимин
гуанин – цитозин
ДНК – это та же РНК, но лишенная одной гидроксильной группы, отсюда и в ее названии префикс дезокси. А урацил – это тот же тимин, но с добавлением метильной группы – СН3. Как видим, поиск – отбор оптимальных сочетаний шел ступенчато. И скорее всего именно от РНК к ДНК, ибо:
- РНК обычно односпиральна,
- рибосомы построены как РНК,
- АТФ включает рибозу…
И тот факт, что у ряда бактерий встречается двуспиральная РНК указивает на отборочные процессы. Победила устойчиво двуспиральная ДНК, оказавшаяся наиболее подходящей для моноядерных клеток на этапе фундаментального эволюционного преобразования прокариотов в эукариоты.
Но опять-таки школярская ясность не должна затемнять эволюционную составляющую. Гносеологически некорректно опираться исключительно на каноническую ДНК (аденин-тимин, гуанин-цитозин), ибо есть и РНК с урацилом. А в ДНК ряда высших растений и многоклеточных животных обнаружен 5 – метилцитозин, а в ДНК ряда бактерий 6 – метиладенин. А ДНК краба вроде бы состоит вообще только из пары: аденин – тимин. И таких примеров множество. Но это задачи эволюционной генетики.
Нуклеоды обеспечивают не только наследственную функцию памяти формы, но и энергообеспечение. И опять-таки аденозинтрифосфорная кислота всего лишь эволюционный доминант, но не абсолют, ибо и другие азотистые основания выполняют подобные функции у ряда до сих пор успешно существующих живых существ. Универсальность АТФ как энергоаккумулятора поразительна, но и поиск-отбор шел мучительно долго. Правда и эта история еще не написана.
Гносеологически важно подчеркнуть, что все химические элементы, входящие во все моно – и полимеры карбонэргической природы всего живого на нашей планете уже были в наличии и 5 млрд. лет назад. Формально это проблема геономическая, но в проблеме биогенеза все тесно взаимопереплетено, что дает возможность реанимации в масс-медиа совершенно алогичных спекуляций на тему обязательного привнесения чего-то неземного в акт творения жизни как отголосок помеси креационизма и панспермии. Причина тому – одноактность, единая и в креационизме, и в панспермии, и в биогенезе. Но это только по форме, а не по сути. Одноактность креационизма и панспермии строго системно-логически есть неизбежно строго круговая цикличность, а одноактность биогенезная – строго спиральная цикличность. Значит различие в степени самой возможности развития: одно по кругу, другое по спирали. Одноактность биогенеза четко выражена в знаменитом принципе Ф. Реди – Л. Пастера: Omne vivum ex vivo. В 1885г. Р. Вирхов сделал принципиально важное уточнение: Omnis cellula ex cellula, сведя тем самым саму категорию “жизнь” к чисто биологической категории “клетка”. Оставляем принцип Ф. Реди – Л. Пастера виталогии и обратимся ко много более близкому проблеме биогенеза принципу Р. Вирхова.
Цитогения – завершающая и много более сложная стадия биогенеза. Иногда даже путают цитогенез с биогенезом под влиянием принципа Р. Вирхова, но это гносеологически некорректно. Биогенез действительно начался с формирования протоклетки но на этом он и завершился, ибо с этого момента уже только клетка могла породить другую клетку и ничто иное. Но протоклетка дала начало до сих пор продолжающемуся процессу цитогении, тогда как биогенез был действительно одноактен и ни одной другой, кроме протоклетки, формы жизни не изобрел. Таким образом, цитогения есть единственная эволюционная форма жизни на Земле, т.е. принцип организации специфической гипоформы на базе уникального стохастического отбора комплекса высокополимерных карбонэргических структур/образований разной степени устойчивости как в отношении факторов внутренних связей, так и в отношении факторов внешней среды.
Цитология ведет свое начало с 1665 г.. Родоначальником ее стал Р. Гук, который первым в сорокократный микроскоп собственного изготовления на срезе пробки увидел, описал и предложил сам термин “клетка”. Эмпирика бурных микроскопических исследований не особо различала цитологию и микробиологию. Лишь в 1838-1839 гг. ботаник М. Шлейден и зоолог Т. Шванн сформулировали клеточную теорию всего живого. В 1845 г. Зибольд окончательно доказал одноклеточное строение всех простейших и назвал их Protozoa, А Р. Вирхов в 1859 г. пришел к выводу, что “клетка есть действительно последний морфологический элемент всех живых тел, и мы не имеем права искать жизнедеятельности вне её”. На основе этого вывода уже через несколько лет Р. Эберхард замахнулся на виталогическое обобщение выдвинув принцип: Omne vivum ab aeternitate e cellula.
Как видим вольно или невольно имела (и до сих пор встречается) гносеологическая и методологическая путаница части и целого, корпоративной, организменной субсистемы и автономного организма-системы, а отсюда и некоторого корпоративного набора функций и автономного поведения. Таким образом, мы здесь снова сталкиваемся с имплицитным, подспудным неразличением целенаправленности, целеполагаемости и целеположенности. Ведь поведением обладает только целеполагающая автономная система-целостный организм, тогда как клетка в многоклеточном организме выполняет функции целеположеной субсистемы, хотя и способна в своих пределах выполнять некоторые целенаправленные функционально действия. А объединяет все клетки от протоклеток до клеток нашего организма только поразительный феномен слитости формы и содержания в виде строгого структурно-системного образования из специфических высокополимерных соединений, которые создают специфическую же способность к единоцельному действию:
→трофика→рост→редупликация→
В основу этого структурно-системного образования положен принцип элементарного многослойного физико-химического фильтра:
Рис. 9
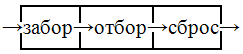
А так как это автономная г/г форма, то и забор и сброс происходит через некоторую граничную оболочку, отделяющую тело от внешнего мира и выступающего в роли фильтра-сита, т.е. имеющего некоторое количество входов и выходов, которые могут как совпадать, т.е. выполнять попеременно функции то входа, то выхода, так и не совпадать, т.е. выполнять свои функции автономно. Но если выборочность забора очевидна из самого принципа фильтра, то выборочность отбора и последующего сброса далеко не очевидна. Отбор происходит по химическому закону соединительных объемов Гей-Люссака (1808г), т.е. внутри г/г формы происходит специфическое свертывание некоторой части забранного извне, а значит и уплотнение вещественно-энергетического образования, создавая возможность следующего забора некоторой порции извне, что явно демонстрирует действие принципа разноплотности. Строго системно-логически забор легко коррелирует с трофикой, а отбор с последующим вещественно энергетическим уплотнением – с ростом. А вот для корреляции сброса с редупликацией явно необходимо некоторое дополнительное условие – ограничитель роста. Таким ограничителем могла быть только сама оболочка в виде ее способности к растяжению, а значит и неспособности к адекватному соразмерному росту. И ответ на этот вопрос надо искать на более глубоком уровне.
Итак, физически протоклетка была явно сфероидной формы, а значит и пульсировала в силу принципа разноплотности во все стороны одинаково своими входами/выходами, поли – или монофункциональный характер деятельности которых требует дополнительных изысканий, ибо он осложняется потенциально возможной безотходностью жизнедеятельности протоклетки с одноступенчатым типом отбора с последующим уплотнением. А это возможно только при изобилии уже готовых требующихся вещественно-энергетических структур во внешней среде.
Где могло возникнуть такое уникальное хронотопное изобилие при учете таких теоретически крайне низких показателях вероятности самого факта цитогенеза даже по Кестлеру, есть главная проблема геобиогенеза. Хоть как-то повысить эти показатели можно уникальным же сочетанием геономических, геохемогенических и геофизических факторов. Наиболее близким этому точечному хронотопу является гипотеза гейзера, где частота чисто механической сталкиваемости дополняется тепловыми, грозовыми, воздушными и инсоляционными воздействиями, которые содействовали проявлению карбонэргических связей. И формированию белковых микросфер, коацерватов. Это полностью соответствует и аксиоме гидро – , лито – , атмосферного стыка как безусловного места биогенеза. Мелководные лагуны стали местом эволюции уже образовавшихся протоклеток. Гносеологически важно здесь постоянно иметь в виду, что речь идет о макромолекулярном уровне протоклетки, т.е. ее чрезвычайно малой габаритности, доступной только очень сильным микроскопам. И речь идет исключительно о принципах организации микросферы/протоклетки в ее биофизическом и биохимическом единстве.
Забор сквозь оболочку из уплотняющихся сетей макромолекул протеина мог производиться исключительно путем электропиноцитоза. Значит и поступление газов было уже включено в пиноцитоз. Обмен ионами протоклетки со средой выполнял связующую роль электрокатализатора и элекроингибитора внутренних процессов относительно внешних, а значит и сами внутренние процессы неизбежно связаны между собой электрохимически. Оболочка автономизировала фактически некоторую часть той же водной среды, что была снаружи. Но это ограничение пространства неизбежно ведет к резкому увеличению взаимодействий тех соединений, которые оказались внутри. Эти взаимодействия неизбежно вели к некоторой структуризации путем свертываемости дружественных образований, их роста – накопления. К ограниченному пространству добавились каталитические способности ряда белков, ускоряющих химические взаимодействия в клетке в десятки и сотни тысяч раз. Но сами ферменты и коферменты в своей активности обусловлены температурой, кислотностью (рН среды), концентрацией. В каждой клетке одновременно происходит до 10 тысяч химических реакций. Таким образом стохастика точечного геономического гидро–лито–атмосферного хронотопа достаточно строго системно-логически объяснима, ибо все было в наличии миллионы лет: аминокислотные остатки, нуклеотиды, источники энергии.
Ограниченное макромолекулярное пространство (коацерватная капля, микросфера) стала основой своеобразного карбонэгического мегаполиса-крепости, выполняющего функции судностроительного дока, т.е. создания копии самого себя.
Гносеологически здесь мы имеем дело с самоорганизацией высокомолекулярных соединений:
аминокислот
и
нуклеотидов,
которые выступают в качестве строительных кирпичей-мономеров, а результатом такой самоорганизации являются:
белки
и
нуклеиновые кислоты,
которые выступают уже в качестве цельных блоков-полимеров.
Сам принцип самоорганизации есть по сути процесс вещественно-энергетической свертываемости начинающийся в химическом мире с формирования атомов тоже при весьма редких сочетаниях множества факторов, прежде всего температуры и давления. Аналогична самоорганизация и кристаллов, но с появлением четко выраженной памяти формы (фундамент ионной решетки). Правда, кристалл самоорганизуется наслоением извне. Гидрогенный характер протоклетки обусловил ее жидкокристаллическую природу, и тем более самым казалось бы коренным образом должен был все поменять, но факт наличия и твердокристаллических форм пребывания вирусов, т.е. РНК/ДНК как способ самосохранения для переживания неблагоприятных условий среды обитания указывает на явное физико-энергетическое сродство памяти кристаллов и памяти РНК/ДНК, как некоторой последовательности операций, но…
И здесь мы сталкиваемся с двумя толкованиями последовательности:
- последовательность как некоторая цепочка;
- последовательность как строгое чередование.
Чтобы разобраться с этим, вернемся к анализу формирования белков и нуклеиновых кислот. Выше мы уже говорили об электрохимической сродственности аминокислот между собой и нуклеотидов между собой.
В
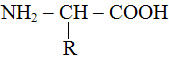
се аминокислоты имеют единую основу
а различаются они друг от друга именно химической группой – R. Аминная группа NH2, имеющая основные свойства, может образовывать пептидные связи (CO – NH) с карбоксильной группой СООН, имеющей кислотные свойства, т.е. остатки аминокислот могут позволить себе любые сочетания любой длины. Теоретически у новоприбывшего в микросферу аминокислотного остатка имеются 4 варианта существования:
- свободное состояние, что в силу сродства маловероятно;
- стать организатором нового полимера, что тоже маловероятно, ибо надо достаточно долго быть самостоятельным;
- присоединиться к уже существующему полимеру пептидной связью и тем самым раствориться в нем;
- присоединиться к уже существующему полимеру боковой водородной связью и положить начало подобию матричного копирования.
Именно подобию, ибо водородные связи в белках неспецифичны, а значит и гарантировать не только количественно, но и качественно соответствие копии образцу невозможно. Значит матричное копирование требует учета не только количества мономеров в полимере (это просто стохастическая цепочка - последовательность), а и их качественных характеристик (последовательности соответствующей качественному составу мономеров в полимере). Значит биогенез и цитогенез (а затем и цитогения) – это непрерывная гармонизация количества и качества, формы и содержания. Количество и форма регулируются оболочкой, а качество и содержание тем, что находится внутри клетки.
Гармонизатором выступил нуклеопротеид – рибосома.
Нуклеотиды обычно смотрятся случайными квартирантами в белковом мегаполисе-крепости, явными инородцами и в количественном и в качественном отношениях, а их поразительная сочетаемость в РНК/ДНК до сих пор не получила должного объяснения. Стохастически такую сочетаемость объяснить невозможно, ибо строгая упорядоченность является зеркальным отражением именно аминокислот в белках, а потом уже и самой себя. Значит вначале на уровне протоклетки нуклеотиды образовали две родственные цепочки мономеров из пуринов и пиримидинов, различающихся только тем, что пиримидины имеют одно углеродно-азотистое кольцо, а пурины – два. Все нуклеотиды связываются в одну цепочку-полимер фосфодиэфирными связями, т.е. кончиками остатков фосфорной кислоты. А строгая комплементарность пар: аденин – урацил/тимин,
цитозин – гуанин, оказалась обусловленной только тем, что все другие при физиологических температурах крайне неустойчивы. Именно это каноническое постоянство только и могло стреотипизировать такое же постоянство и в белковой среде. А вот как это произошло, остается темным. Генетики работают над тем, как функционирует ДНК как данность. А нас здесь волнует вопрос как возникла сама ДНК не в структурно-количественном, а в структурно-качественном отношении именно как информационной системы. У прокариотов доминируют не ДНК, а РНК. Следовательно и эволюционно РНК первичны. Но как возникли сами РНК как информационные системы? Ведь чтобы знать все белки клетки, надо их как минимум измерить. И уже доказано, что каждой аминокислоте соответствует триплет-кодон цепочки нуклеотидов. Число возможных сочетаний из 4 нуклеотидов по 3 равно 64. Сам факт трехкратной избыточности системы кодирования относительно якобы строго 20 базовых аминокислот уже вызывает вопросы. Вполне возможно, что 20 базовых есть результат эволюционного отбора, а на этапе протоклетки их было гораздо больше.
Но генетики описывают процесс транскрипции и трансляции, однако эволюционно транскрипции строго системно-логически должна предшествовать прескрипция, т.к. прежде чем переписать, надо сначала записать. Таким образом необходимо объяснить что и как:
- измеряет количественно и качественно;
- сводит эти результаты в некоторый эталон.
Д
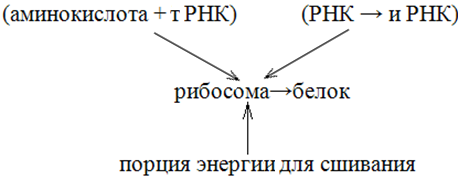
елать это могли только нуклеопротеиды как медиаторы в виде рибосом, удивительно однородных по величине, строению и химическому составу от бактерий до человека. Транспортных РНК ровно столько, сколько аминокислот, а информационных РНК ровно столько, сколько белков. Получаем цепочку технологий – сцепок:
Рис. 10
Итак, самоорганизация присуща и нуклеотидам в РНК и аминокислотам в белках, но только нуклеопротеиды обладают способностью объединить эти два параллельных и совершенно независимых процесса в некоторую систему, т.е. системно-логическое соответствие структурной последовательности путем взаимодействия. И неизбежно возникает вопрос, что чему задавало последовательность? Если структура самообразовавшихся белков служила моделью-образцом первичного копирования нуклеопротеидами, то затем эта нуклеопротеидная структура могла:
- стать основой формирования базовой РНК либо присоединиться к уже существующей;
- могла развалиться на некоторые части, став основой:
а) рибосомы (м РНК),
б) и РНК,
в) т РНК.
Первый вариант достаточно строго системно-логически объясняет, откуда РНК могла знать структуру белка, т.е. сам феномен прескрипции.
Второй вариант объясняет сродственную вариативность элементов системного синтеза.
Однако генез феномена триплетности кодирования остается темным. Но тайна сия явно не в белках или РНК, а в нуклеопротеидах.
Если структура самообразовавшейся цепочки РНК служила моделью-образцом первичного копирования нуклеопротеидами, то появление новых белков ставит проблему их встраивания – взаимодействия с уже существующими.
Отсюда следует, что нельзя исключать и третий вариант формирования РНК – смешанный, т.е. белковомодельные структуры и модельнобелковые структуры образуют единобазовую РНК.
Таким образом, суть жизнедеятельности состоит в системно-образующей деятельности нуклеопротеидов в рамках полузамкнутого пространства микросферы протоклетки, т.е. без медиаторной функции комплементарных нуклеопротеидов любые по сложности белки и РНК/ДНК остаются не более чем карбонэргическими структурами, а собственно биохимическими только в триедином комплексе непрерывного функционирования: аминокислоты – нуклеопротеиды – нуклеотиды. А феномен прескрипции обусловливает процесс эволюции.
Эти выводы полностью согласуются с аналогичной непрерывной изменчивостью атомов, но свернутость атомов обусловлена внутренними связями при открытости структуры, тогда как протоклетка способна функционировать только как ограниченное макромолекулярно от внешнего мира некоторое внутреннее, автономное пространство. Свертываемость идет по линии: (ион) – атом – молекула – макромолекула. И именно ионы передают протоклетке электромагнитную зарядность оболочной структуры, регулирующей все взаимоотношения мира внутреннего с миром внешним и наоборот. Вещественно-энергетическая свернутость неизбежно обусловлена некоторым температурным спектром и спектром давления, которые являются прямыми трансформами разноплотности – энергии. Геобиогенезная устойчивость обоих спектров обусловила первую биофизическую закономерность: физиологическую температуру – оптимум биохимического развития. Гидрогенный характер формирования протоклетки при всех эволюционных коллоидальных изысках обусловил ее жесткую зависимость от воды, но в то же время на первых порах нивелировал в силу физических свойств воды в данном спектре температур от колебаний внешнего давления, что позволило сформироваться механизму общения внутреннего мира с внешним путем регуляции давления внутреннего, т.е. феномену электропиноцитоза, базирующегося на двух формах диффузии: диализ и осмос (диффузия – один из физико-химических трансформов той же разноплотности). Сама белковая оболочка была долгое время до образования протоклетки и некоторое время после продуктом самообразования белков из аминокислот. Строго многослойные протеино-липидные оболочки – это уже продукт эволюции протоклетки, постепенного усиления авторитаризма, автократизма РНК с последующим революционным скачком РНК→ДНК и образованием в биохимическом мегаполисе – крепости, клетке, административного замка в виде ядра обособленного типа, т.е. превращение прокариота в эукариота. Подавляющее большинство белков стали целиком зависеть от администрации клетки, ибо именно она (РНК/ДНК) давала задания своим мастерам – нуклеопротеидам. Однако часть белков в отличие от четкого конвейера матричного копирования на рибосоме продолжала создаваться кустарно-ремесленным путем, т.е. ферментами. Строго системно-логически это может означать только то, что количество кустарно-ферментных белков крайне ограничено, требуются они не постоянно, а время от времени. В то же время вариативность такого типа белкогенеза много выше, отзывчивость тоже, а это весьма важно при разного рода ЧП на клеточном уровне. Значит и здесь имеем дело не с недочетами администрирования из замка-ядра, а с весьма важной стратегически функциональной комплементарностью производства массового и эксклюзивного. Однако называть это некоей целесообразностью было бы некорректно, ибо это не есть результат принятия решения некоей целеполагающей системы, а всего лишь чисто энергетическое ограничение, ибо единая валюта биомира – биохимическая энергия – весьма и весьма дорога. Таким образом все белки клетки четко делятся на матричные и ферментные, а значит и на левосторонние и нейтральные относительно поляризации, по их происхождению. Это несомненно как-то связано и выражается в классификации белков по их функционально доминантным свойствам:
- пластические (самостроительные и иностроительные);
- сократительные (обеспечившие затем все двигательные функции);
- регулятивные (ускоряющие и замедляющие ход реакций в виде катализаторов и ингибиторов в их сложнейшей внутренней иерархии и взаимодействии);
- консервативные (способные хранить необходимые вещества прозапас);
- агрессивно-разрушительные (для многих других веществ и даже самой клетки);
- сигнальные (на основе осцилляции, обратимой денатурации или необратимой денатурации, которые упреждающе включают механизмы других химических реакций защитного типа);
- энергетические (но только на самый крайний случай из-за их важных предыдущих шести свойств).
Но, если все аминокислоты имеют единую основу, то значит все специфические свойства белков-полимеров обусловлены количеством и последовательностью их радикальных групп, т.е. даже одна новая радикальная группа способна придать и/или лишить какого-то или каких-то свойств.
По сути протобионт представлял собой белковый населенный пункт, проходящий этап становления стойбища-привала множества туристов-мигрантов разных национальностей, слабо или совсем незнающих языков друг друга. Только количественно эти на нацгруппы: разные: от одной до сотен и тысяч. Кто-то уходит, а кто-то приходит, но остающиеся обязаны как-то привыкать друг к другу. А привыкать – это прежде всего узнавать, идентифицировать, ибо от этого зависит, что делать самому:
- уничтожить, обезвредить, т.е. разложить или связать;
- нейтрально пройти мимо;
- завязать знакомство для последующего возможного сотрудничества, взаимопомощи, созидания.
Заметим, что в химии высокомолекулярных соединений реакции происходят не совсем так, а то и совсем не так, как в минеральной химии. И это касается не только доминации эндотермики над экзотермикой (в неорганической химии доминируют реакции с выделением тепла, а здесь с поглощением тепла, что имеет кардинальное значение для функционирования протобионта и его последующих модификатов). На химическом уровне таких знаков всего два: электрический (+,0,-) и формообразующий (отсюда важность учета стереометрии молекул).
На неорганическом уровне всегда используется метод уничтожения путем ковровой бомбардировки самопокрытием, а для высокомолекулярных соединений более важен точечный удар, ибо, убрав только одну аминокислоту, мы лишаем вредного для нас свойства весь белок-полимер, что много выгоднее чисто энергетически.
Нейтральность, безразличие, индифферентность фактически проявляется одинаково в обеих химиях. Но во взаимодействиях высокомолекулярных соединений тысячекратно возрастает лицемерие, коварство, подлость и опять-таки из-за одного-двух автоматических добавлений к полимеру. Пораздельно они безобидны, а одна добавка вдруг превращает белок в монстра-убийцу.
Лояльность, приязнь и даже любовь через временный союз тоже не редкость, а некоторые только и делают, что сводятся-разводятся. Понятно, что дело не в чувствах, а в энергетическом самосохранении. Итак, многомиллионолетний этап формирования протобионта – это непрерывная притирка белковых полимеров друг к другу в рамках ограниченного пространства, которое неизбежно многократно увеличивает частоту взаимодействий, а значит и отбор. Именно ограниченное пространство сузило влияние стохастики до некоторого подобия упорядоченности. Однако до более-менее приемлемого порядка было бы невозможно по законам стохастики химического мира прийти, чтобы преобразовать их в законы стохастики мира биогенного, без 1) создания более прочных границ и правил входа-выхода, т.е. упорядочения взаимоотношений с изменчивым внешним миром необходимо устойчивого мира внутреннего, и 2) упорядочения самого мира внутреннего по принципу “чужие тут не ходят”, ибо без принятия такого принципа в белковом мегаполисе невозможна была бы эффективная идентификация одних белков другими, а значит и сам порядок. А чтобы белки были узнаваемо своими, выход был только один: производить их на месте, т.е. в самом же белковом городе. Как только формирование полимера-полипептида заканчивается, он сначала скручивается в спираль вдоль своей оси, а затем еще и спираль складывается в максимально компактную форму комочка. И это имеет свой смысл в деле самосохранения белка как самостоятельного гражданина белкового города, ибо не позволяет ни присоединиться, ни отсоединиться даже одному аминокислотному остатку в цепи полимера (чем это чревато, мы уже говорили выше). Кроме того компактизация и вторичная (спираль), и третичная (комочек) много эффективнее защищают от разрушительных сил как внутри, так и в особенности вне клетки (в частности, от радиации). Наконец, компактизация – это и неизбежное следствие перенаселения в клетке, т.е. феномен гармонизирующих сил мутуализма: взаимополезные уступки ради мирного взаимовыгодного сосуществования. И особой умности во всем этом белку совсем не нужно, ибо те, кто так не делал, просто гибли. Для внутриклеточного сосуществования традиции всегда священны и оправданно священны, ибо, только следуя этим традициям, жизнь как биологический феномен и смогла сохраниться едва ли не 5 млрд. лет. В закрытости, т.е. эгоизме, своекорыстии клетки кроется феномен ее выживаемости даже при экстремальных (в условиях Земли) температурных гидро – , лито – и атмосферных режимах. В науке это называется достижением и поддержанием гомеостаза. И возможно это только путем непрерывного вещественно-энергетического обмена внутреннего мира клетки с внешним через оболочные регуляторы (входы/выходы). Мы описали схематически вещественные процессы, теперь надо так же систематизировать и процессы собственно энергетические.
Гравитационная энергия явно нивелировалась водной средой, в которой и работал биореактор, порождая протоклетки.
Электромагнитная энергия, базовая для химического мира в целом, уже была задействована в процесс и результат создания протоклетки в виде ионного калий-натриевого насоса оболочки и т.п.
Диффузионная энергия разноплотностей (диализ, осмос) тоже проявляется на оболочном уровне.
Тепловая энергия пронизывает всю жизнедеятельность, получив специальное название “физиологические температуры” как базовой характеристики комфортного гомеостаза. Обусловлена эта жесткая зависимость живого от температуры спецификой биохимических (карбонэргических) реакций. Термоэволюция живого изучена и описана еще весьма и весьма слабо от отдельных биохимических реакций прокариотов до человека, но уже ясно, что зависимость от внешней тепловой энергии вряд ли бы позволила жизни продвинуться вперед сколько-нибудь далеко в ее эволюции. Упорядочивание тысяч непрерывных одновременно последовательно-паралельных биохимических реакций в клетке в строго целенаправленную систему требует автономии от состояния внешнего изменчивого мира в виде некоторых вещественно-энергетических запасов. Делать вещественные запасы даже для протоклетки особой трудности не представляло, ибо она сама уже была вещественной. Другое дело – запасы энергетические, ибо они требуют специального механизма связывания-развязывания, свертывания-развертывания, которые в минеральной химии требуют высоких температур и давлений. И вновь поразительно экономное решение биогенеза: трифосфатные комплексы тех же нуклеотидов что входят в состав РНК/ДНК. Однако эволюция по каким-то причинам отдала предпочтение аденозинтрифосфату (АТФ). Это и есть главный и вечный биоаккумулятор, совершающий разрядку и зарядку: АТФ
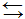 АДФАМФ путем последовательного разрушения и восстановления макроэргических связей, дающих (каждая) энергии ровно на сшивку двух аминокислот или наоборот, на их расшивку. Подзарядка осуществляется обычно углеводами. Но их-то биореактор почему-то вырабатывал много меньше, чем это необходимо было все увеличивающемуся количеству протоклеток. Наступил энергетический кризис. Шли напряженные поиски неуглеводных источников питания и были достигнуты определенные успехи, но в особо специфических условиях, препятствующих глобализации жизни. Правда, и эти достижения (хемосинтез) вошли составной частью в вещественно-энергетический кругооборот биосферы. Но генеральное решение энергетического кризиса пришло с формированием фотосинтеза, который стал поставлять углеводы (сахара в виде крахмала) практически в неограниченном количестве. АТФ вновь заработали на полную мощность и жизнь совершила качественный скачок от протоклетки к моноциту. Для этого ей пришлось энергетически переориентироваться на Солнце. Закончилась эра геобиогенеза. Началась эра гелиоцитогении.
АДФАМФ путем последовательного разрушения и восстановления макроэргических связей, дающих (каждая) энергии ровно на сшивку двух аминокислот или наоборот, на их расшивку. Подзарядка осуществляется обычно углеводами. Но их-то биореактор почему-то вырабатывал много меньше, чем это необходимо было все увеличивающемуся количеству протоклеток. Наступил энергетический кризис. Шли напряженные поиски неуглеводных источников питания и были достигнуты определенные успехи, но в особо специфических условиях, препятствующих глобализации жизни. Правда, и эти достижения (хемосинтез) вошли составной частью в вещественно-энергетический кругооборот биосферы. Но генеральное решение энергетического кризиса пришло с формированием фотосинтеза, который стал поставлять углеводы (сахара в виде крахмала) практически в неограниченном количестве. АТФ вновь заработали на полную мощность и жизнь совершила качественный скачок от протоклетки к моноциту. Для этого ей пришлось энергетически переориентироваться на Солнце. Закончилась эра геобиогенеза. Началась эра гелиоцитогении.Теперь, когда мы уяснили принципы физико-химических процессов забора – отбора, т.е. биофизико-биохимических трофики-роста, пришла пора проанализировать конечную стадию функционирования протоклетки: сброс/редупликация. Ведь геобиореактор на протяжении всей своей активности сам порождал множество микросфер. А вот живое стало порождать живое, точнее, по Вирхову, клетка клетку только с обретением способности к редупликации.
То, что исполнителями деления клеток на две равные части являются сократительные белки, сомнений уже не вызывает. Они же выполняют главную функцию при сбросе из клетки всего ненужного, а то и вредного. Темным остается вопрос, что заставляет их так поступать, ибо кроме как сокращаться они ничего более не умеют. Правда они могут работать в ансамбле плавно-последовательно, либо взрывно-одновременно, а это значит обладать способностью некоторого сочувствования ансамблевой работы, т.е. владеть реакцией на натяжение. Сугубо сгибательный принцип действия сократительных белков требует неизбежно их специфической архитектоники в клетке в виде последовательно-параллельных цепочек зеркального типа, могущих обеспечить пульсацию “сокращение-распрямление”. Доминация цитологических исследований на эукариотах подталкивает к мысли, что редупликация регулируется ядром. Но это мало доказательно для многоядерных прокариотов и тем более для протоклетки. Счетный принцип “мономер-полимер” позволяет предположить, что причиной деления на две части является достижение некоторого критического объема, включающего автоматически сократительные белки, такой себе кварковый принцип на макромолекулярном уровне. Но это еще нужно доказать, что экватор не успевает за ростом микро – сферы и срабатывает в обратом направлении. Ведь как бы ми ни антропоморфизировали все процессы в клетке, решений там принимать некому, а весь механизм строится исключительно на строгой системно-логической упарядоченности, последовательности.
Однако конец эпохи протоклетки мы ведем не с затухания геобиореактора, а с момента обретения первого жгутика, дополнившего соматомоторику целенаправленной локомоторикой, т.е. дополнение пульсации собственно движением, в пространстве. Жгутик позволил не подчиняться косной физико-химической среде, т.е. много эффективнее реализовать возможность самосохранения. Но возможность самосохранения неизбежно предполагает наличие самой способности самосохранения. А это уже качественно новая характеристика сугубо биогенного мира: система базовых потребностей на базе самосохранения. Таким образом, принцип сугубо косного физико-химического фильтра в ходе биогенеза трансформировался в биофизико-биохимический автомат, т.е.
→забор→отбор→сброс→
→трофика→рост→редупликация→
А жгутик завершил формирование полноценного бионта уже в таком виде:
Модель бионта
(система базовых потребностей)
Р
ис. 11
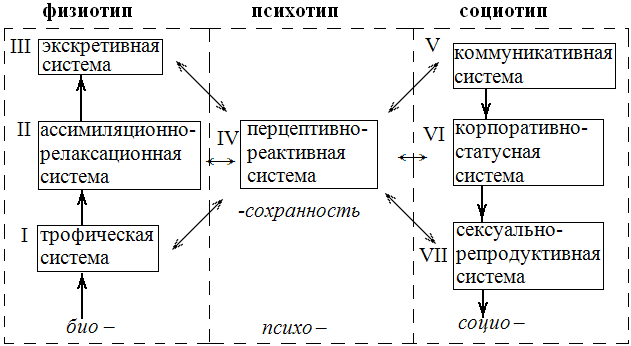
Обоснованию того, как организменно элементарная (но вещественно-энергетически архисложная по системно-логической упорядоченности) структура-автомат “→ трофика → рост → редупликация →” стохастически эволюционно непрерывно совершенствует свою “Систему базовых потребностей” вплоть до полного абсурда самоотрицания, самопожертвования и будет посвящено все дальнейшее изложение/описание мира биогенного и мира гуманогенного. Однако прежде всего нам придется проанализировать именно эволюцию бионта как физиотипа, т.к. завершился геобиогенез, но продолжается геоцитогения.
Гносеологически геоцитогения четко делится на пять этапов:
I доклеточный (протоцит),
II одноклеточный (моноцит),
III многоодноклеточный (ассоциоцит),
IV одномногоклеточный (полицит),
V многомногоклеточный (социоцит).
Протоцит мы уже описали, хотя и в грубо контурном виде. Биогенный мир обрел триединую единицу анализа – бионта в его физиотипической, психотипической, социотипической ипостасях.