
Особенности развития трутней в пчелиных семьях apis mellifera L. На территории краснодарского края >06. 02. 10 частная зоотехния, технология производства продуктов животноводства
| Вид материала | Автореферат диссертации |
- Программа вступительного экзамена в аспирантуру по специальной дисциплине 06. 02., 48.04kb.
- Продуктивные и физиологические показатели импортного мясного скота в условиях краснодарского, 399.9kb.
- Продуктивные качества красной степной и чёрно-пёстрой пород при совершенствовании, 504.38kb.
- Программа минимум кандидатского экзамена по специальности 06. 02. 04 «Частная зоотехния,, 206.32kb.
- Обоснование технологии производства молока и молочных продуктов в условиях введения, 915.78kb.
- Интенсификация воспроизводства и повышение продуктивности свиней с использованием биотехнологических, 1068.55kb.
- Продуктивность, некоторые биологические особенности бычков абердин-ангусской и симментальской, 399.25kb.
- Пути повышения эффективности производства молока при интенсификации животноводства, 662.32kb.
- Интенсификация производства говядины в мясном скотоводстве 06. 02. 10 частная зоотехния,, 864.33kb.
- Программа-минимум кандидатского экзамена по специальности, 71.9kb.
На правах рукописи
Абрамчук Алексей Васильевич
ОСОБЕННОСТИ РАЗВИТИЯ ТРУТНЕЙ В ПЧЕЛИНЫХ СЕМЬЯХ
APIS MELLIFERA L. НА ТЕРРИТОРИИ КРАСНОДАРСКОГО КРАЯ
06.02.10 – частная зоотехния, технология производства
продуктов животноводства
Автореферат диссертации на соискание ученой степени
кандидата сельскохозяйственных наук
Дивово 2010
Работа выполнена в Кубанском государственном университете
Научный руководитель: доктор биологических наук, доцент
Морева Лариса Яковлевна
тор биологических Ярошенко В.А.
Официальные оппоненты: доктор сельскохозяйственных наук, доцент
Пустовой Виталий Филиппович
кандидат сельскохозяйственных наук,
доцент Редькова Лидия Анатольевна
Ведущая организация: ГНУ Научно-исследовательский институт пчеловодства Российской академии сельскохозяйственных наук
Защита состоится «30» ноября 2010 года в 9 часов на заседании диссертационного совета Д 006.018.01 при ГНУ Всероссийском научно-исследовательском институте коневодства по адресу: 391105, Рязанская область, Рыбновский район, пос. Дивово, п.о. Институт коневодства.
С диссертацией можно ознакомиться в научной библиотеке ГНУ ВНИИ коневодства.
Автореферат разослан 2010 г.
Ученый секретарь
диссертационного совета Готлиб М.М.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Медоносные пчелы (Apis mellifera mellifera L.) занимают обширный ареал, включающий зоны с холодным, умеренным и жарким климатом. Пчелиная семья представляет собой сложную целостную биологическую и хозяйственную единицу. Особенности экологических условий влияют на характер проявления тех или иных признаков у рабочих пчел и трутней. Трутни являются временными особями пчелиной семьи, что связано с ограниченной их функцией, направленной только на спаривание с маткой. Многие вопросы биологии трутней в теплом климате Краснодарского края изучены недостаточно.
Отсутствие государственного контроля над сохранностью генетических ресурсов и антропогенное влияние приводят к определенной трансформации и метизации трутней южных популяций (Кривцов, 2000). Несмотря на прогресс в исследовании медоносных пчел (Apis mellifera mellifera L.), морфофункциональная межпопуляционная изменчивость и структура популяций до настоящего времени остаются недостаточно изученными и, прежде всего, плохо изучены популяции южных регионов. Еще более немногочисленны данные о морфологии трутней в чистопородных и помесных семьях серой горной кавказской породы. Недостаточны сведения о температурной зависимости изменения строения крыльев трутней. Это является актуальным для понимания возможных изменений функциональных свойств их летательных аппаратов, играющих важную роль при спаривании с маткой, а, следовательно, успешность передачи генетической информации. Все это сказывается на ведении селекционной и племенной работы в пчеловодстве. В связи с этим определенный интерес представляет изучение особенностей выращивания трутневого расплода пчелиными семьями в конкретных условиях Краснодарского края.
Актуальным является использование перспективного сырьевого источника, а именно – гомогената трутневого расплода. Весьма малоизученным является влияние комплексного воздействия гомогената трутневого расплода – биларпродукт на молодняк сельскохозяйственных животных в качестве биологически активной добавки.
Цель и задачи исследования.
Целью работы является изучение особенностей развития чистопородных и помесных трутней в теплом климате Краснодарского края. Для достижения намеченной цели необходимо было решить следующие задачи:
- Установить влияние микроклимата гнезда на развитие и жизнедеятельность трутней в пчелиных семьях территории Краснодарского края;
- Изучить сезонную динамику развития трутней и рабочих пчел в пчелиных семьях;
- Проследить динамику выращивания трутней пчелиными семьями с разновозрастными матками в ульях разной конструкции;
- Выявить возрастную изменчивость чистопородных и помесных трутней;
- Изучить морфометрические показатели экстерьера и аномалий жилкования крыльев трутней чистопородных и помесных пчел серой горной кавказской породы;
- Показать возможность использования трутневого расплода в качестве биологически активной кормовой добавки.
Научная новизна. Впервые в условиях теплого климата Краснодарского края установлены закономерности развития трутней в чистопородных и помесных семьях пчел серой горной кавказской породы. Установлено влияние температуры на формирование экстерьерных признаков развивающихся трутней в чистопородных и помесных семьях серой горной кавказской породы пчел (Apis mellifera caucasica Gorb.), детально описаны особенности их развития.
Дана сравнительная характеристика морфологических и морфометрических признаков чистопородных и помесных трутней медоносных пчел. Установлено, что помесные трутни подвержены большим изменениям морфометрических признаков по сравнению с чистопородными, а также изучено изменение количества крыловых зацепок, влияющие на силу сцепления крыльев во время полета. Впервые для юга России изучены аномалии в жилковании крыльев трутней чистопородных и помесных пчел серой горной кавказской породы. Показана возможность использования гомогената трутневого расплода в качестве биологически активной кормовой добавки.
Теоретическая и практическая значимость работы. Экспериментально установленные сведения параметров внутригнездового микроклимата в зоне выращивания трутневого и пчелиного расплода на территории Краснодарского края в ульях различной конструкции, позволяют регулировать сроки появления трутней на пасеках. Сведения о сезонной и температурной изменчивости размеров крыльев, их жилкования и аномалий жилкования как чистопородных, так и помесных трутней серых горных кавказских пчел важны для понимания фенотипической изменчивости трутней и ведения селекционной работы в пчеловодстве.
Испытана эффективность использования трутневого расплода в страусоводстве, в результате проводимых опытов с 2009 года на страусиной ферме ООО «С.Е.В.» применяют гомогенат для подкормки всех отродившихся страусят.
Материал исследований используется при чтении курсов лекций: "Животноводство", "Физиология насекомых", "Экология насекомых", "Основы пчеловодства и опыление энтомофильных культур" в Кубанском государственном университете на биологическом факультете и при преподавании пчеловодства в институте начального и среднего профессионального образования.
Основные положения, выносимые на защиту:
1. Особенности сезонного выращивания чистопородных и помесных трутней на территории Краснодарского края при различных погодных условиях, в ульях различной конструкции и с разновозрастными матками.
2. Сезонная изменчивость морфометрических признаков и аномалий жилкования крыльев чистопородных и помесных трутней серой горной кавказской пчелы.
3. Возможность использования гомогената трутневого расплода в качестве биологически активной кормовой добавки.
Апробация работы Результаты исследования доложены на межреспубликанских научно-практических конференциях «Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий», проводимых Кубанским государственным университетом (Краснодар, 2009-2010 гг.); XIII съезде Русского энтомологического общества, (Краснодар, 2007); конференции посвященной 100-летию со дня рождения Г.Ф. Таранова (Рыбное, 2007); Международной конференции "Пчеловодство-XXI. Темная пчела (Apis mellifera mellifera L.) в России" (Москва, 2008); Международных научно-практических конференциях по пчеловодству (Новосибирск, 2008, 2009); Научно-практической конференции «Основные направления развития пчеловодства на современном этапе» (Рыбное, 2009).
Публикации. По материалам диссертации опубликовано 16 работ, в том числе 3 в изданиях, рекомендованных ВАК Минобрнауки России.
Объем и структура диссертации. Диссертационная работа изложена на 125 страницах печатного текста, включает 21 таблица, 29 рисунка. Состоит из введения, 4 глав, выводов, списка литературы. Список использованной литературы включает 135 источников, из них 18 на иностранном языке.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Научно-методические аспекты изучения трутней
В главе приведен обзор многочисленных литературных источников и фондовых материалов по теме исследования. Среди них особо отмечены работы, затрагивающие вопросы исследования трутней пчелиной семьи (Алпатов, 1948; Тряско, 1955; Таранов, 1970, 1973; Кривцов, 1993; Лебедев, 1998; Бурмистрова, 1999).
Глава 2. Материал и методы исследования
В основу работы положены данные, полученные автором в процессе лабораторных и пасечных исследований в 2005 по 2010 гг. Материалом для исследований послужили чистопородные и помесные пчелиные семьи серой горной кавказской породы, подверженные влиянию местных абиотических факторов. Работа выполнялась на учебной пасеке АПИ-лаборатории Кубанского государственного университета и экспериментальной пасеке ООО "Павловский мед". На всех экспериментальных участках были выделены пчелиные семьи разной силы. Температурный режим гнезда пчелиных семей определяли по методике предложенной Е.К. Еськовым (1991) с помощью термодатчиков типа ММТ-4, изготовленных в НИИ пчеловодства для определения температуры в разных точках улья. При организации и проведении работ руководствовались Методическими указаниями к постановке экспериментов в пчеловодстве (М. Россельхозакадемия, 2000).
Отбор материала для морфометрических исследований проводили каждый месяц. При этом на сотах с расплодом на выходе ставили колпачок и на следующий день проводили отбор молодых трутней, которых взвешивали и фиксировали в 70% этиловом спирте. Морфометрические исследования проводили по методике, предложенной В.В. Алпатовым (1948). Препараты изготавливали по методике
Г.Д. Билаш и Н.И. Кривцова (1985). Морфометрические признаки измеряли с помощью окуляр-микрометра стереоскопического микроскопа МБС-10. Анализировали 11 морфометрических признаков: длину хоботка, длину переднего левого крыла, длину переднего правого крыла, ширину переднего левого крыла, ширину переднего правого крыла, длину заднего левого крыла, длину заднего правого крыла, ширину заднего левого крыла, ширину заднего правого крыла, число зацепок на заднем левом крыле, число зацепок на заднем правом крыле.
Количество расплода определяли рамкой-сеткой с квадратами 5×5 см, охватывающими по 80 ячеек трутневого расплода.
Полученные данные подвергнуты статистической обработке методом вариационной статистики с проверкой достоверности результатов с помощью критерия Стьюдента с использованием программы «Statistica 6.0».
Глава 3. Природно-климатическая характеристика Краснодарского края
В главе на основании литературных данных приводится краткая характеристика природных условий территории исследования.
Глава 4. Особенности развития трутней в пчелиных семьях Аpis mellifera L. на территории Краснодарского края
4.1 Влияние температурного фактора на развитие и жизнедеятельность трутней
Срок появления трутней зависит от состояния пчелиной семьи, а также от внешней температуры, оказывающей влияние на их развитие. От ее сезонных изменений во многом зависит количество и размещение трутней в семье, но численность их в несколько раз ниже численности пчел. Первый трутневый расплод появляться в феврале и обычно локализуется в зоне пчелиного расплода единичными экземплярами, поэтому температурные условия их развития одинаковые. В марте, несмотря на значительные перепады внешних температур, количество трутневого расплода увеличивается до 88−120 ячеек в отдельных пчелиных семьях. В апреле трутневый расплод располагается на сотах выше пчелиного, в ячейках, где раньше был мед, при таком расположении трутневых личинок пчелам легче поддерживать оптимальную температуру 35−36 0С. Максимальное его количество в этот период составляет 400 ячеек.
Температура воздуха в апреле не опускалась ниже +12 0С, а максимальная достигает +19 0С. Такие положительные температуры воздуха способствуют меньшей затрате энергии пчелами на поддержание оптимальной температуры (+35−36,5 0С) в зоне пчелиного и трутневого расплода. Для развития трутневого расплода необходима среднесуточная температура воздуха не менее + 15 0С, обычно такая температура характерна для II декады апреля. На территории Краснодарского края очень часто отмечаются весенние похолодания, которые приходятся на I декаду мая, снижая количество как пчелиного, так и трутневого расплода в пчелиных семьях. Часто во время весенних похолоданий пчелы начинают изгонять трутней.
Учитывая, что внутригнездовая температура подвержена влиянию внешней, в летний период с установившейся жаркой погодой в гнездах семей у расплода происходит повышение температуры соответственно с повышением внешней. Так при повышении внешней температуры до +35−40 0С в зоне трутневого расплода она колебалась в пределах +36−37,2 0С. При экстремально высокой внешней +42−44 0С, как это было в 2010 году, и невозможности создать внутри гнезда оптимальные температурные условия, трутни отрождались с морщинистыми и редуцированными крыльями или пчелы проявляли каннибализм, освобождаясь от трутневого расплода и сохраняя пчелиный.
4.2 Особенности роста и развития стаз в пчелиных семьях
Пик развития трутневого расплода в центральных, северных регионах, а также на юге России совпадает при условии, если лето прохладное, он приходится на II декаду июля (Морева, Абрамчук, 2008).
Многолетние наблюдения на исследуемой территории дали возможность выявить закономерности их развития в жаркие и экстремально жаркие летние периоды. Если в северных регионах развитие происходит по одновершинной кривой, то развитие трутней в южных регионах, имеется иные закономерности. Максимальное количество трутневого расплода в пчелиных семьях на территории Краснодарского края отмечено в III декаде марта, а в очень теплые зимы при колебании средней температуры от минус 1 до 1 оС в отдельных пчелиных семьях трутни могут появляться во II декаде февраля – I декаде марта.
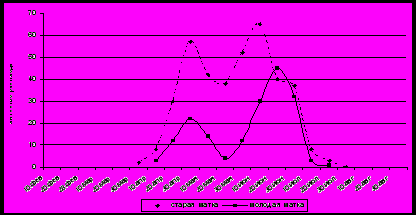
Результаты наших исследований по развитию трутневого расплода показали, что динамика его развития имеет вид двухвершинной кривой (рис. 1).

Рисунок 1. Динамика выращивания расплода пчелиными семьями на территории Краснодарского края.
Первый пик массового появления трутней в III декаде апреля − I декаде мая обычно совпадает с цветением плодовых. Уже в конце мая наступает незначительный спад в развитии трутней. Это связано с тем, что в период цветения белой акации погодные условия сильно меняются, чаще всего идут дожди, резко снижается температура до +10−15оС. В этих условиях для пчел наступает первый медосборный период с акации. Пчелы изгоняют значительное количество трутней из семей, и матка сокращает откладку трутневых яиц (рис. 1). Обычно после окончания цветения акации начинается второй пик роста трутневого расплода и максимум приходится на II декаду июня, на период цветения подсолнечника. В это время отмечается и роевое состояние отдельных пчелиных семей.
На территории Краснодарского края до 90 % трутневого расплода пчелы выращивают за 120 дней (с конца апреля до конца июля) больше всего в I−II декаде мая, во II декаде июня. На исследуемой территории в начале марта и в начале августа трутней выращивают только отдельные пчелиные семьи. Теплые зимы 2003 и 2009 года на территории края характеризовались отсутствием отрицательных температур, а дневные иногда поднимались до +20°С, это обусловило нетипичное поведение пчел в отдельных семьях. Январские оттепели активизировали пчел, а необходимая сумма положительных температур, которая наблюдалась во II декаде января и послужила тому, что пчелы заставили матку отложить яйца и из них вывели новую матку. Повторный осмотр, проведенный 12.03.04, показал наличие в улье открытого расплода, но трутневого, и 24.03.04 появлялись первые вышедшие трутни. Молодая матка откладывала трутневые яйца из-за отсутствия в ее семяприемниках мужского семени. Первый засев матки-трутовки находится в центре клуба и зачастую бывает правильной, т.е. сплошной засев на 50−80 ячеек. Лишь по «горбатой» печатке можно определить наличие в семье матки-трутовки.
Учитывая, что в Краснодарском крае в пчелиных семьях есть тенденция ранней закладки маточников, мы решили исследовать подвижность сперматозоидов у трутней, появившихся 24.03.04 г., с этой целью отбирали трутней в возрасте 8 и 12 дней. В первые дни жизни трутни незрелы в половом отношении, что было выявлено при исследовании спермы, взятых из семенных пузырьков. Было установлено, что сперматозоиды подвижны и у трутней возраста 8 дней и возраста 12 дней, но у трутней в возрасте 12 дней количество сперматозоидов значительно больше и составило 4,5−5 млн. в 1 мм3 и они обладают большей активностью. В это время сперма лучше поддается извлечению из половых органов трутней и наиболее пригодна для осеменения маток. Наблюдения за спермой под микроскопом показали, что сперматозоиды сохраняют подвижность в течение 35−40 мин. при температуре +25оС. Без покровного стекла сперматозоиды теряют активность и становятся неподвижными уже через 3–5 мин. Ранневесенние трутни для осеменения маток должны быть в возрасте 13–15 дней, когда они наиболее активны и хорошо эякулируют; в этот период у них максимальное накопление в семенных пузырьках жизнеспособной спермы.
Иногда из зимовки некоторые пчелиные семьи могут выйти без матки и из открытого расплода подставленного пчеловодом из другой семьи, пчелиная семья выводит новую матку. К моменту выхода ее во II и III декадах апреля будут трутни способные спариваться с маткой. Конечно, по всем признакам они не будут лучшего качества, чем летние, но выведенные апрельские матки будут оплодотворенные, а семьи пчел получат раньше развитие, чем, если бы трутни появились в конце апреля как обычно в нашем крае.
Таким образом, ставший уже закономерностью, теплый температурный режим зимы в центральной части Краснодарского края, ведет к изменениям жизнедеятельности в пчелиных семьях, что заслуживает более детального изучения.
4.2.1 Сезонное выращивание трутней в помесных и чистопородных пчелиных семьях серой горной кавказской породы
Нами установлено, что в помесных семьях трутни развиваются раньше, чем в чистопородных, их пчелиные семьи раньше вступают в роевое состояние. Роевой период у них начинается в мае и продолжался 2–3 недели – до III декады июня. В семьях чистопородных пчел случаев роения с молодой маткой, не наблюдалось. Только в конце июня чистопородные пчелиные семьи входили в роевое состояние.. На равнинной территории Краснодарского края, на которой преобладают помесные пчелы, образовавшиеся в результате спаривания кавказских маток с трутнями украинской степной, карпатской и других пород, сохраняют склонность выращивать много трутневого расплода и, как следствие, являются более ройливыми, чем серые горные кавказские чистопородные. Мы сравнили количество трутневого расплода выращиваемого в течение года чистопородными и помесными пчелиными семьями. Ранней весной в марте помесные пчелы выращивали в 9 раз больше трутней, чем чистопородные, майские в 7,6 раза, а июльские в 3 раза.
Максимальное количество трутневого расплода в семьях чистопородных и помесных пчел отмечено в I-II декадах июня. Первое появление трутней в 2006 и 2009 гг. отмечено в помесных пчелиных семьях во III декаде марта, а в чистопородных – во II декаде апреля (рис. 2).
чистопородные
помесные
Рисунок 2. Выращивание трутневого расплода в помесных и чистопородных пчелиных семьях.
В конце III декады апреля отмечен первый пик появления большого количества трутней, но чистопородные пчелы выращивали в 3 раза меньше трутней, чем помесные. Второй пик приходится у помесных на II декаду июня, а у чистопородных на III декаду июня. Наблюдения показали, что количество трутней в отдельных семьях составляло 3500 ячеек печатного расплода, но откладка трутневого расплода у помесных семей оставалась в 1,5 раза выше.
По окончании медосбора пчелы в семьях принимают меры к экономному расходованию собранных кормовых запасов: они изгоняют из ульев ненужных теперь трутней. Матки значительно снижают, а затем и вовсе прекращают откладку яиц. Наблюдения показали, что в III декаде августа 2005 г. как в чистопородных серых горных кавказских, так и в помесных семьях отмечалось активное изгнание трутней из ульев. В единичных семьях серых горных кавказских пчел трутни встречались в сентябре-октябре. А в одной семье трутни были замечены в III декаде октября и в начале ноября. Стремление оставлять трутней в зиму или вывод их ранней весной говорит о том, что матки этих семей, скорее всего, будут сменяться пчелами в новом сезоне.
4.2.2 Динамика выращивания трутневого расплода в пчелиных семьях с разновозрастными матками в ульях различной конструкции
Весенние и летние похолодания, нередко наблюдающиеся в условиях Краснодарского края, тормозят наращивание расплода в семье медоносных пчел. Колебания температуры при смене дня и ночи, приводит к тому, что в условиях близких к оптимальному находится только центр расплодного гнезда. Так как трутневой расплод находится чаще всего в периферической части сотов, следует учесть тип сота, в ячейках которого происходит развитие трутней. В течение ряда лет мы наблюдали откладку трутневых яиц матками различного возраста в ульях различной конструкции: на рутовскую, дадановскую и украинскую рамку.
Все основные показатели пчелиных семей (количество пчел, кормовых запасов, качество и количество сотов) на начало опытов были практически одинаковыми. Для подопытных пчелиных семей были созданы одинаковые условия содержания и ухода.
| Рисунок 3. Динамика выращивания трутневого расплода пчелиными семьями в ульях на рутовскую рамку. 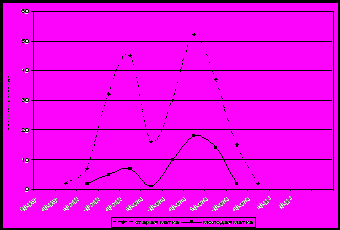 Рисунок 4. Динамика выращивания трутневого расплода пчелиными семьями в ульях на Дадана-Блатта. Рисунок 4. Динамика выращивания трутневого расплода пчелиными семьями в ульях на Дадана-Блатта.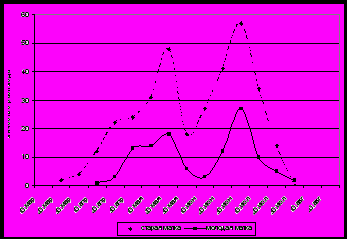 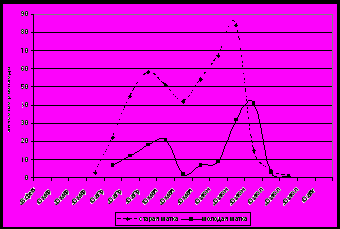 Рисунок 5. Динамика выращивания трутневого расплода пчелиными семьями в ульях новой конструкц. Рисунок 5. Динамика выращивания трутневого расплода пчелиными семьями в ульях новой конструкц. | Семьи со старыми матками откладывают неоплодотворенных яиц (трутневых) больше, чем матки первого года жизни, они и позже начинают откладывают трутневые яйца. Обычно первое появление трутней на данной территории происходит в ульях на рутовскую рамку и ко II декаде апреля составляет более 800 ячеек. Следовательно, расплодная часть в улье занимает значительный объем. Рутовские рамки, имеющие габариты 435 × 230 мм способствуют тому, что весенний, расплодный участок клуба занимает значительную часть сотов, поэтому в нижней его части матки откладывают неоплодотворенные яйца (рис. 3). Старые матки обычно уже во II декаде марта откладывают неоплодотворенные яйца, а молодые – на месяц позже. В ульях на дадановскую рамку трутневой расплод отмечен в начале апреля небольшими участками, а у маток первого года жизни он появляется во второй декаде апреля (400−640 ячеек) (рис. 4). Как и в ульях рутовской конструкции, старые матки в ульях системы Дадана-Блатта откладывают трутневые яйца на декаду раньше. В ульях на украинскую рамку, нашей конструкции, появление трутневого расплода в семьях со старыми матками отмечается в I декаде апреля, а с молодыми − в I декаде мая (рис. 5). |
У молодых маток, в ульях нашей конструкции на украинскую рамку, количество трутневого расплода в начале мая не превышало 480 ячеек, а у маток старших возрастов − 3760 ячеек.
При содержании пчел в этих ульях не встречаются матки старше двух лет, так как они всегда сменяются молодыми, выведенными в вспомогательных отделах улья. Старшего возраста сохранялись только чистопородные матки, для получения гибридов первого поколения, сохраняющие свои хозяйственно-полезные признаки.
На юге трутни появляются значительно раньше, чем в центральных и северных регионах, улей нашей конструкции может сдерживать появление большого количества их, а для содержания и разведения медоносных пчел в условиях теплого климата нашего края требуется своя специфическая технология, адаптированная к особенностям семьи и в полной мере соответствующая местным условиям. С этой целью на платформах с ульями нашей конструкции ставились ульи на рутовскую рамку для получения ранних трутней.
4.2.3 Трутневый расплод в годовом цикле пчелиных семей содержащихся ульях различной конструкции
Проведенные исследования по годовому циклу развития трутневого расплода в ульях различной конструкции подтвердили, что в ульях на рутовскую рамку они появляются на месяц раньше в I декаде февраля (рис. 6).
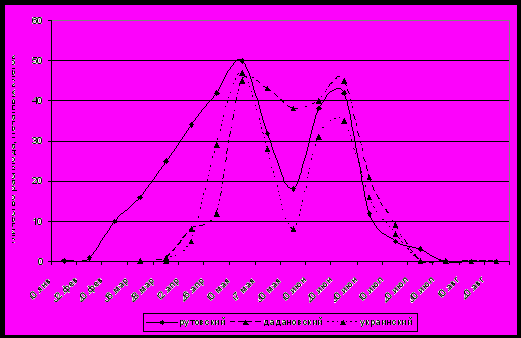
Рисунок 6. Динамика развития трутневого расплода в пчелиных семьях в 2010 году.
Динамика развития трутневого расплода в пчелиных семьях в годы с экстремально высокими наружными температурами (42−44оС) происходило изгнание трутней из ульев всех конструкции и отсутствие их в августе.
4.2.4 Возрастная изменчивость некоторых признаков трутней
Трутни, как и матки, участвуют в образовании генетической структуры семьи. В период исследования мы выявляли оптимальный возраст трутней, пригодных для осеменения, определяя количество и концентрацию спермы на разных этапах развития, а также их активность. Изучение вели с 1-го по 25-й день жизни трутней по пяти возрастным группам. На выходе трутней метили лаком разного цвета, чтобы определить их возраст. Затем определяли их массу (табл. 1).
Таблица 1 - Размеры и масса трутней различных генераций и породностей
| Генерация трутней | Длина тела (мм) | Масса (мг) | |||||
| однодневные | 25-дневные | ||||||
| | чис. | пом. | чис. | пом. | чис. | пом. | |
| Ранневесенние единичные (февраль) | 14−15 | 13−14 | 219±2,6 | 192±1,8 | 182±1,4 | 187±1,6 | |
| Ранневесенние (апрель) | 16−20 | 14−19 | 267±3,1 | 245±2,8 | 204±2,2 | 193±1,9 | |
| Весенние (май) | 16–19 | 17−20 | 272±3,4 | 277±2,6 | 212 ± 2,8 | 230±2,2 | |
| Летние (июнь I декада) | 15−18 | 14−17 | 258±2,3 | 218±1,9 | 206±2,08 | 198±2,0 | |
| Летние (июль II декада) | 17−19 | 18−19 | 265±3,6 | 271±3,2 | 242±2,6 | 251±2,7 | |
| Летние (август II декада) | 19−19 | 17−18 | 263±3,4 | 259±2,4 | 245±2,1 | 234±1,8 | |
По размерам тела ранневесенние трутни отличаются от весенних и были самыми меньшими 13–15 мм, так как они развивались в пчелиных ячейках. Весенние трутни (16−20 мм) развивались в единичных трутневых ячейках, а летние - 17−19 мм и развивались в трутневых ячейках на нижнем краю сотов, поэтому размеры тела их больше на 3–6 мм (табл. 1). По весу трутни, вышедшие в марте, также весили на 2,5−4,5 мг меньше чем летние трутни. Показано, что от диаметра ячеек, в которых развивались трутни, зависит их масса, размеры тела и его придатков. Так массы тела в среднем уменьшилась в 1,5 раза. Вероятно, от размера ячеек зависит трофическое обеспечение развивающихся в них личинок. Пчелы, ориентируясь по величине ячеек, снабжают личинок кормом тем обильнее, чем больше ячейки. Величина ячейки служит фактором, детерминирующим трофическую активность пчел-кормилиц.
Наблюдения показали, что лёт более тяжелых трутней был активнее, но число их заметно уменьшалось. Это дает нам возможность полагать, что тяжеловесные трутни раньше созревают и активнее принимают участие в спаривании с матками, это же отмечает И.И. Хруст (1984). Оценивая качество трутней по этим признакам можно предположить, что крупные трутни с более развитой мускулатурой будут более сильными, и первыми будут участвовать в процессе осеменения маток.
Возрастные изменения массы тела трутней представлены на рис. 7, 8. Определение массы трутней различных генераций чистопородных и помесных пчел показало, что однодневные трутни весят больше, чем 25-дневные, т.е. от рождения идет снижение массы тела трутней.
Вариабельность массы тела помесных трутней больше, чем чистопородных, и с возрастом приводит к изменению массы тела от рождения до половой зрелости, что наглядно отображено графически.
Как отмечает Н.Д. Шмелёва (1975), уменьшение массы трутней, так же, как и неплодных маток, вероятно, имеет глубокий биологический смысл, своего рода приспособленность к спариванию в воздухе, т.е. самому главному инстинкту – продлению рода.
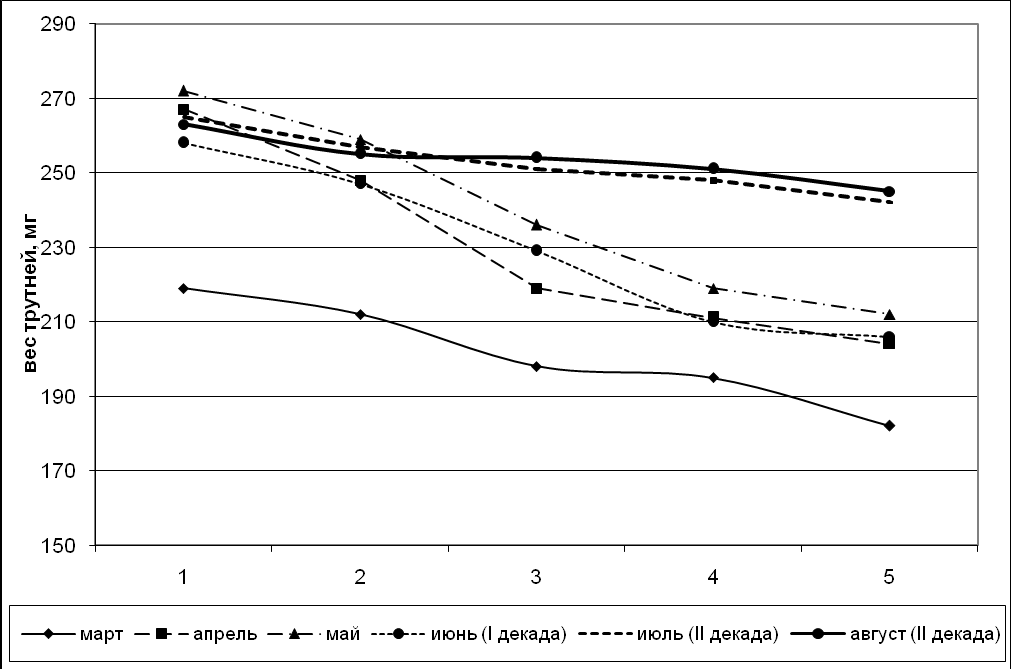
Рисунок 7. График возрастного изменения массы чистопородных трутней.
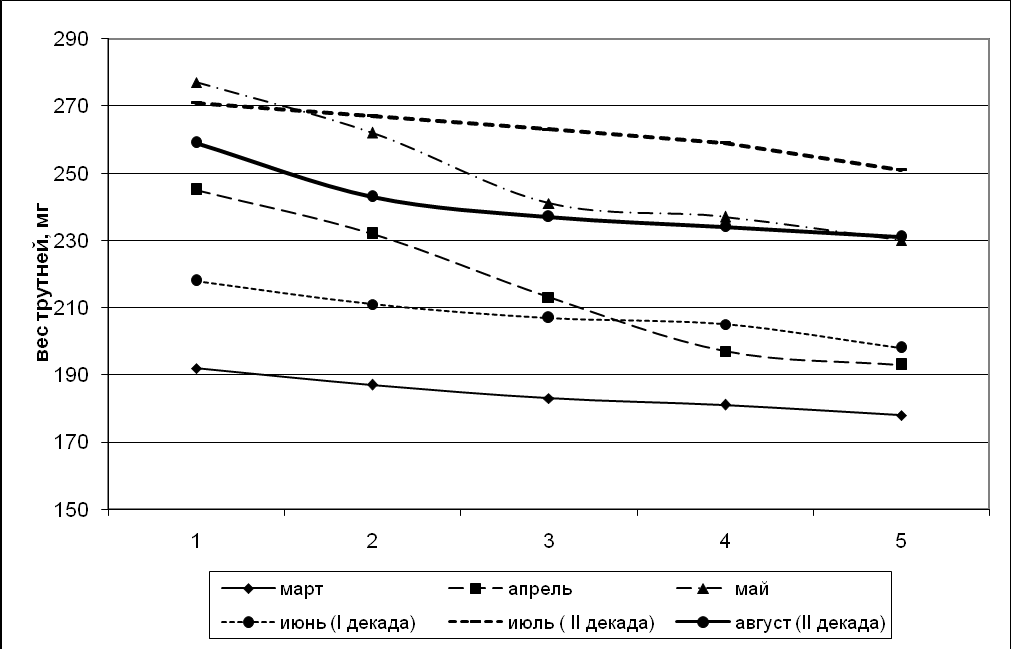
Рисунок 8. График возрастного изменения массы помесных трутней.
Следовательно, появление трутней в ранневесенний период способствует раннему осеменению маток.
4.3 Морфометрические признаки трутней в чистопородных и помесных семьях серой горной кавказской породы
Учитывая немногочисленные сведения по изменчивости морфометрических признаков трутней, мы в своих исследованиях поставили задачу тщательно изучить некоторые экстерьерные и интерьерные признаки трутней помесных и чистопородных пчелиных семей серой горной кавказской пчелы. Одиним из главных морфометрических признаков является длина хоботка. Он изменяется и у трутней, не только в зависимости от породы, популяции, физико-географических факторов, но и по периодам активного сезона. Сезонную изменчивость хоботка можно рассматривать как определенную реакцию организма на изменение таких внешних факторов, как климат и условия питания. У весенних трутней длина хоботка варьирует от 3,4−3,2 мм у чистопородных и 3,7–3,6 мм у помесных. У особей летней генерации при повышении температуры до 36−38 0С происходит уменьшение длины хоботка как у чистопородных так и помесных трутней серой горной кавказской породы составляет в среднем 0,2−0,3 мм. С повышением внешней температуры до 40−45 0С, длина хоботка трутней увеличивается. Размеры хоботка у чистопородных варьирует от 3,7 до 4,0 мм, у помесных – от 3,5 до 3,8 мм (табл. 2).
Таблица 2 - Сезонные изменения длины хоботка чистопородных и помесных трутней
| | Чистопородные | Помесные |
| Апрель | 3,412± 0,022 | 3,777± 0,032 |
| Май | 3,174± 0,061 | 3,612± 0,014 |
| Июнь | 3,772± 0,031 | 3,518± 0,026 |
| Июль | 3,861± 0,032 | 3,845± 0,046 |
| Август | 3,870±0,038 | 3,411 ±0,027 |
Наибольших размеров большое крыло трутней достигает в начале летнего период в июне (11,8340,037 мм) и в сентябре (11,940±0,032 мм). С повышением температуры повышается тенденция к уменьшению крыльев как у чистопородных, так и помесных трутней. Минимальных размеров они достигают в июле, когда отмечены высокие температуры окружающей среды и соответственно в ульях и ранней весной, когда трутни чаще всего выращиваются в пчелиных ячейках сотов.
Если в июле пчелы не в состоянии поддерживать оптимальную температуру в расположении расплода и она будет превышать +38 °С, а трутней пчелы, как правило, выращивают по краям расплодного гнезда, где температура повышается значительно выше чем в центре у молодых трутней, закончивших развитие, крылья остаются в зачаточном состоянии развития на крайней границе верхнего температурного оптимума 37–37,2 °С, длина оказывалась минимальной.
Такие же сезонные различия характерны и для малых крыльев. По их длине чистопородные и помесные трутни имеют незначительные отличия на 0,055 мм, значительные их отличия в июне 0,515 мм, а ширина у малого крыла у помесных трутней имеет значительные отличия от 0,157 до 0,534 мм.
Максимальные их размеры отмечены весной в апреле и составляют в среднем 8,125±0,018 мм у помесных и 7,516±0,021 мм у чистопородных, а минимальные − в июле 8,00±0,04 мм, а у чистопородных наблюдается тенденция к повышению, при внешней температуре, выходящей за пределы оптимальной.
По данным ряда авторов (Радченко, 1980 и др.), количество зацепок на малом крыле не подвержено сезонной изменчивости. Е.К. Еськов (1990, 1992) отмечает, что количество зацепок на крыльях пчел одной и той же расы может варьировать в 1,5 раза.
По нашим данным, имеет место максимальное количество крыловых зацепок у трутней, которые развиваются при оптимальной температуре. Отклонение от нее к верхней границе выше оптимальной отражается на уменьшении размеров малого крыла, а следовательно и к уменьшению краевых зацепок.
Количество крыловых зацепок у трутней уменьшается с повышением температуры до +37 С в зоне расплода, от 24 до 17 штук. Укорочение длины крыльев в июле приводит и к снижению количества зацепок. Весной и осенью размеры малого крыла, и число зацепок увеличивается. Анализируемый показатель отличался большим диапазоном изменчивости у трутней весенне-летней генерации, чем у трутней выведенных в апреле − мае по количеству зацепок. Их количество варьировало весной апрель − май от 22 до 23,5 штук. В июне – июле: от 19,5 до 23 штук. В августе от 21,8 до 23,6 штук.
Сезонная изменчивость крыльев к осени и весной выражается в увеличении их площади и укрепления сцепления. Последнее как раз и достигается за счет увеличения количества зацепок. Следовательно, происходит усиление крылового аппарата, так необходимого в связи с увеличением размеров крыльев и массы тела трутней. Это происходит к началу завершения их репродукции, особенно в роевой период у помесных в конце мая и количество зацепок составляет 24,6 штук, а у чистопородных 23,5 штук.
Среднее значение количества зацепок на левом крыле больше чем на правом. В среднем на одну зацепку различаются левые и правые крылья помесных трутней, а у чистопородных трутней количество зацепок практически не имеет различий. Проведенные нами морфометрические исследования показали, что у чистопородных трутней количество зацепок варьирует от 20 до 23, а у помесных от 18 до 24.
Для определения достоверности различий генерации трутней за май, июнь и июль нами был проведен однофакторный дисперсионный анализ морфометрических признаков табл. 3.
Трутни различных генераций достоверно отличаются по следующим признакам: ширина правого малого крыла, ширина левого большого крыла, ширина левого малого крыла, ширина правого большого крыла, длина тела, ширина тела.
Таблица 3 − Дисперсионный анализ морфометрических признаков трутней серой горной кавказской породы территории Краснодарского края за весенне-летний период
| Морфометрические признаки | Сум.квад эффект | Ср.квад. эффект | Сред.кв. ошибки | Ср.квад. ошибки | F | p<0,5 | Дост. |
| Длина тела | 78,06 | 39,03 | 85,60 | 0,82 | 47,88 | 0,00 | * |
| Ширина тела | 3,69 | 1,85 | 9,58 | 0,09 | 20,23 | 0,00 | * |
| Длина левого большого крыла | 4,77 | 2,39 | 261,88 | 2,49 | 0,96 | 0,39 | |
| Ширина левого большого крыла | 1,53 | 0,76 | 16,55 | 0,16 | 4,84 | 0,01 | * |
| Длина левого малого крала | 1,00 | 0,50 | 29,53 | 0,28 | 1,78 | 0,17 | |
| Ширина левого малого крыла | 1,91 | 0,95 | 12,42 | 0,12 | 8,07 | 0,00 | * |
| Длина правого большого крыла | 5,17 | 2,58 | 100,96 | 0,96 | 2,69 | 0,07 | |
| Ширина правого большого крыла | 1,30 | 0,65 | 17,83 | 0,17 | 3,83 | 0,02 | * |
| Длина правого малого крала | 1,34 | 0,67 | 32,66 | 0,31 | 2,16 | 0,12 | |
| Ширина правого малого крыла | 0,90 | 0,45 | 14,50 | 0,14 | 3,25 | 0,04 | * |
| Зацепки левого крыла | 22,86 | 11,43 | 750,14 | 7,14 | 1,60 | 0,21 | |
| Зацепки правого крыла | 35,20 | 17,60 | 694,90 | 6,62 | 2,66 | 0,07 | |