
«Белки. Нуклеиновые кислоты»
| Вид материала | Исследование |
Содержание3.1.2 Лекция по теме «Белки. Нуклеиновые кислоты» |
- Лекция Нуклеиновые кислоты. Атф нуклеиновые кислоты, 119.07kb.
- Нуклеиновые кислоты, 296.24kb.
- Нуклеиновые кислоты, 122.08kb.
- Современные методы исследования бас выделение и анализ бас, 65.85kb.
- Белки составляют основу жизнедеятельности всех организмов, известных на нашей планете, 482.65kb.
- Современные методы исследования бас выделение и анализ бас, 182.61kb.
- «Нуклеиновые кислоты», 105.98kb.
- Урок 5 (9). Нуклеиновые кислоты и их роль в жизнедеятельности клетки. Атф и другие, 199.62kb.
- Нуклеиновые кислоты и атф. Автор(ы):, 111.28kb.
- Имд «Амины, аминокислоты, белки», 40.89kb.
3.1.2 Лекция по теме «Белки. Нуклеиновые кислоты»
План лекции
- Белки.
- Понятие о белках.
- Состав белков. Пептидная связь.
- Свойства белков. Разнообразие белков и их роли в живых организмах.
- Ферменты-биокатализаторы. Механизм действия ферментов.
- Понятие о белках.
- Нуклеиновые кислоты.
- Дезоксирибонуклеиновая и рибонуклеиновая кислоты, их состав, свойства и функции.
- Принцип комплементарности.
- Дезоксирибонуклеиновая и рибонуклеиновая кислоты, их состав, свойства и функции.
- Биосинтез белка.
1. Среди биологически важных полимеров видное место занимают белки и нуклеиновые кислоты, входящие в состав живой клетки и играющие особую роль при возникновении и развитии живых организмов.
В состав белков входят 20 протеиногенных аминокислот, которые кодиpyютcя генетичеcким кодом и постоянно oбнapyживaютcя в белкax.
Главными структурными единицами белков и пептидов являются остатки аминокислот, связанные карбоксамидной пептидной связью между б-карбоксильной и б-аминогруппой.
- NH – C=O
Каждый белок характеризуется специфичной аминокислотной последовательностью.
При выяснении структуры белков необходимо установить не только число и природу остатков, но также и порядок их чередования в макромолекуле. Для решения этой задачи производят последовательное отщепление аминокислот с того или другого конца полимерной молекулы с последующей идентификацией их. В методе Эдмана, например, белок обрабатывают раствором фенилизотиоцианата в пиридине и полученный продукт присоединения — раствором НС1 в нитрометане. При этом концевой остаток отщепляется в виде соответствующего фенилтиогидантоина без изменения остальной части макромолекулы:
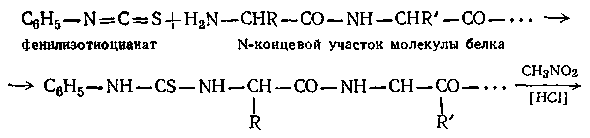
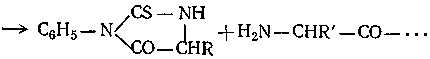
Фенилтиогидантоин
Щелочной гидролиз фенилтиогидантоина приводит к образованию свободной аминокислоты, которая идентифицируется методами бумажной хроматографии:
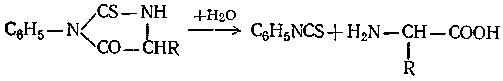
Теоретически, повторяя этот процесс многократно, можно отщеплять поочередно все остатки первоначальной белковой молекулы, установив тем самым их взаимное расположение в макромолекуле. Практически, однако, ввиду сложности задачи она была полностью решена только для некоторых белков несложного строения, как, например, инсулин. При этом выяснилось, что в размещении остатков аминокислот по цепи макромолекулы у биологически активных белков отсутствует та регулярность, которая нередко встречается у других полимеров. В то же время у каждого вида белка наблюдается строго определенная последовательность аминокислотных звеньев.
В белковой молекуле некоторые группы, не участвующие в образовании пептидной связи, остаются свободными или используются для создания мостиков между линейными цепями. Благодаря наличию свободных ионогенных кислых или основных групп белки являются полиамфолитами.
Непосредственное образование пептидной связи из групп СООН и аминогруппы, как показывает термодинамический расчет, должно протекать с увеличением свободной энергии системы. Следовательно, синтез белка из аминокислот может произойти только в том случае, если он сопровождается другими процессами, протекающими с уменьшением свободной энергии. В клетках живых организмов такими процессами являются окисление и гликолиз (биохимический распад молекулы глюкозы на 2 молекулы пировиноградной кислоты); энергия, освобождающаяся при этом, в значительной степени концентрируется в виде пирофосфатных связей молекул аденозилтрифосфорной кислоты (АТФ):
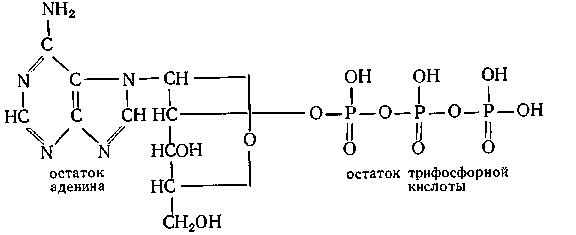
АТФ в реакциях схематически изображается так:

Аналогичные обозначения применяются для соответствующих монофосфорной (АМФ) и дифосфорной кислот (АДФ):
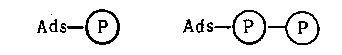
Использование энергии пирофосфатных связей в простейшем случае можно представить как результат образования промежуточного смешанного ангидрида аминокислоты (АК) и АМФ, который более реакционноспособен, чем сама аминокислота:
Ads – ОРО(ОН) – ОРО(ОН) – ОРО(ОН)2 + НООС – CHR – NH2
АТФ АК
Ads – OPO(OH)OOC – CHR – NH2 + H2P2О7
смешанный ангидрид АМФ-АК
Характерной особенностью биологически активных белков является легкость, с которой они изменяются под влиянием тепла, ферментов кислот и различных органических соединений. При этом происходит денатурация белка с полной утратой его биологической активности. Денатурация меняет специфическую пространственную конформацию макромолекулы, но не сопровождается гидролизом ковалентных связей. В живых организмах эта конформация возникает в результате взаимодействия боковых ответвлений полипептидных цепей, являясь термодинамически неравновесной; во время денатурации белок переходит в равновесную денатурированную форму. При достаточно сильном воздействии ферментов, тепла и различных химических агентов может произойти расщепление макромолекулы на отдельные аминокислоты вследствие гидролиза по пептидным связям.
Молекулярный вес различных белков — от десятка тысяч до нескольких миллионов. В состав живых организмов входит несколько видов белков. При использовании белков в качестве пищи организм перерабатывает их в другие, характерные для него белки. Благодаря наличию реакционноспособных групп в макромолекуле белок часто находится в клетках не в свободном состоянии, а в виде протеидов, т. е. комплексов с другими низкомолекулярными или высокомолекулярными веществами. К таким протеидам относятся нуклеопротеиды, хромопротеиды и др.
По химическому составу белки делятся на простые, состоящие только из аминокислотных остатков, и сложные. Сложные белки могут включать ионы металла (металлопротеины, или металлопротеиды), пигмент (хромопротеины, или хромопротеиды), нуклеиновыми кислотами (нуклепротеины), а также ковалентно связывать остаток фосфорной кислоты (фосфопротеины), углевода (гликопротеины) или НК (геномы некоторых вирусов). Состав аминокислот, образующих белки, выражается общей формулой:
 ,
,в которых радикал может содержать различные функциональные группы (R= - SH, OH,- COOH, -NH2) и кольца. -Аминокислоты в белках ковалентно соединены между собой пептидными связями:

Белковая молекула может состоять из одной или нескольких полипептидных цепей, содержащих от 2-3 десятков до нескольких сотен аминокислотных остатков каждая.
Образование пептидных связей происходит в результате взаимодействия карбоксила одной аминокислоты с аминогруппой другой. При этом из 2 -аминокислот образуются пептиды с выделением одной молекулы воды:

Из трех аминокислот образуются трипептиды, из большого числа аминокислот – полипептиды.
Функции, выполняемые белками, распределяются примерно следующим образом.
Структурообразующие функции. Структурные белки отвечают за поддержание формы и стабильности клеток и тканей. В качестве примера структурного белка – коллаген. К структурным белкам можно отнести также гистоны, функцией которых является организация укладки ДНК в хроматине. Структурные единицы хроматина, нуклеосомы, состоят из октамерного комплекса гистонов, на который навита молекула ДНК (DNA).
Транспортные функции. Наиболее известным транспортным белком является гемоглобин эритроцитов (слева внизу), ответственный за перенос кислорода и диоксида углерода между легкими и тканями. В плазме крови содержатся множество других белков, выполняющих транспортные функции. Так, преальбумин переносит гормоны щитовидной железы — тироксин и трииодтиронин. Ионные каналы и другие интегральные мембранные белки осуществляют транспорт ионов и метаболитов через биомембраны.
Защитные функции. Иммунная система защищает организм от возбудителей болезней и чужеродных веществ. Например, иммуноглобулин G, который на эритроцитах образует комплекс с мембранными гликолипидами.
Регуляторные функции. В биохимических сигнальных цепях белки осуществляют функции сигнальных веществ (гормонов) и гормональных рецепторов. В качестве примера здесь представлен комплекс гормона роста соматотропина с соответствующим рецептором. При этом экстрацеллюлярные домены двух молекул рецептора связывают одну молекулу гормона. Связывание с рецептором активирует цитоплазматические домены комплекса и тем самым обеспечивает дальнейшую передачу сигнала. В регуляции обмена веществ и процессов дифференцировки принимают решающее участие ДНК-ассоцированиые белки (факторы транскрипции). Особенно детально изучено строение и функции белков-активаторов катаболизма и других бактериальных факторов транскрипции.
Катализ. Среди 2000 известных белков наиболее многочисленную группу составляют ферменты. Самые низкомолекулярные из них имеют мол. массу 10-15 кДа. Белки среднего размера, как, например, приведенная на схеме алкогольдегидрогеназа, имеют мол.массу 100-200 кДа. Молекулярная масса высокомолекулярных ферментов, к которым относится глутаминсинтетаза, построенная из 12 мономеров, могут достигать 500 кДа.
Двигательные функции. Взаимодействие актина с миозином ответственно за мышечное сокращение и другие формы биологической подвижности. Гексамер миозина длиной 150 нм — один из наиболее крупных белков. Нитевидный актин (F-актин) образуется путем полимеризации относительно небольших молекул глобулярного актина (G-актин). Процессом сокращения управляют ассоциированный с F-актином тропомиозин и другие регуляторные белки.
Запасные функции. В растениях содержатся запасные белки, являющиеся ценными пищевыми веществами. В организмах животных мышечные белки служат резервными питательными веществами, которые мобилизуются при крайней необходимости.
В настоящее время различают первичную, вторичную и третичную структуры белковой молекулы.
Первичная структура белка – его химическая структура, т.е. последовательность чередования аминокислотных остатков в полипептидной цепи данного белка.
Вторичная структура белка – форма полипептидной цепи в пространстве. Установлено, что полипептидные цепи природных белков находятся в скрученном состоянии – в виде спирали. Спиральная структура удерживается водородными связями, возникающими между группами СО и NH аминокислотных остатков соседних витков спирали. Подобная вторичная структура получила название -спирали. Водородные связи в ней направлены параллельно длинной оси спирали (-спирали чередуются с аморфными частями). Такое представление является общепризнанным. Вытянутые полипептидные цепи имеет лишь небольшое число белков, например, белок натурального шелка – фиброин, вязкая сиропообразная жидкость, затвердевающая на воздухе в прочную нерастворимую нить.
Третичная структура белка – реальная трехмерная конфигурация, которая принимает в пространстве закрученная спираль полипептидной цепи. В простейших случаях третичную структуру можно представить как спираль, которая в свою очередь свернута спиралью. У такой структуры в пространстве имеются выступы и впадины с обращенными наружу функциональными группами. Третичной структурой объясняется специфичность белковой молекулы, ее биологическая активность. Определяющими факторами образования и удержания третичной структуры белков являются связи между боковыми радикалами аминокислотных остатков (дисульфидные мостики атомов серы, солевые мостики из аминогруппы и карбоксила, водородные мостики)
Физические и химические свойства белков
Строением белков объясняются их весьма разнообразные свойства. Они имеют разную растворимость: некоторые растворяются в воде, другие – в разбавленных растворах нейтральных солей, а некоторые совсем не обладают свойством растворимости (например, белки покровных тканей). При растворении белков в воде образуется своеобразная молекулярно-дисперсная система (раствор высокомолекулярного вещества). Некоторые белки могут быть выделены в виде кристаллов (белок гемоглобина крови).
Химические свойства
1) Подобно аминокислотам, белки проявляют амфотерные свойства. При действии щелочей белок реагирует в форме аниона – соединяется с катионом щелочи, образуя соль альбуминат:

2) При действии же кислот он становится катионом, образуя синтонин:

Если в молекуле белка преобладают карбоксильные группы, то он проявляет свойства кислот, если же преобладают аминогруппы, - свойства оснований. Присутствие белка можно обнаружить рядом цветных реакций. Эти реакции свойственны составным частям белка – аминокислотам или образуемым ими группировкам.
3) Биуретовая реакция (реакция на наличие пептидных связей). Биурет образуется при нагревании мочевины с отщеплением от нее аммиака:

4) Нингидриновая реакция.
Эта реакция характерна для аминогруппы в -положении. Белки с нингидрином дают синее или фиолетовое окрашивание:

5) Ксантопротеиновая реакция.
Эта реакция характерна для бензольного ядра циклических аминокислот. При действии крепкой азотной кислоты на эти аминокислоты происходит нитрование кольца с образованием нитросоединений желтого цвета [48, 49].

2. К белковым веществам относятся ферменты, или энзимы, выполняющие в живом организме функцию катализаторов высокой селективности и при очень мягких условиях. Это избирательное действие обусловлено комплиментарностью структур реагирующего субстрата и фермента — тем, что заряд или выступающая группа на поверхности одного из них отвечает противоположному заряду или полости у другого (принцип «ключа к замку» — см. рис. 12). Вследствие этого молекулы фермента и субстрата настолько сближаются, что резко возрастает эффективность межмолекулярных сил, противостоящих тенденции молекулярно-кинетического движения разъединить взаимодействующие частицы, происходит специфическая адсорбция (образование фермент-субстратного комплекса). Те же силы могут играть существенную роль в самом возникновении структурного соответствия между субстратом и ферментом.
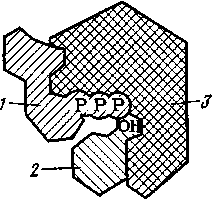
Рис. 12. Схематическое изображение фермент-субстратного комплекса, участвующего в реакции фосфорнокислого остатка АТФ с группой ОН глюкозы: 1 — молекула АТФ; 2—молекула глюкозы; 3 — фермент
Так как дальнейшая реакция протекает в пределах комплекса, в котором молекулы реагирующих веществ благоприятно ориентированы, резко возрастает число эффективных столкновений и, следовательно, скорость реакции. В простейшем случае гидролиза производных кислот под действием биокатализаторов, содержащих только одну активную группу (В :), роль фермента можно схематически изобразить следующим образом:
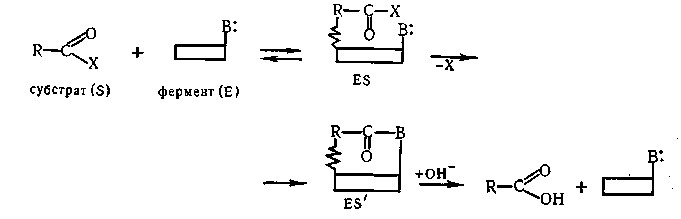
При наличии в ферменте (например, б-химотрипсин) двух групп противоположной электрохимической природы (—О- и —NH+) они могут участвовать в согласованной электрофильно-нуклеофильной атаке на субстрат, что значительно облегчит разрыв соответствующей связи и снизит энергию активации реакции («кооперативный эффект»). Вероятный механизм гидролиза сложного эфира ароматической кислоты под влиянием б-химотрипсина:
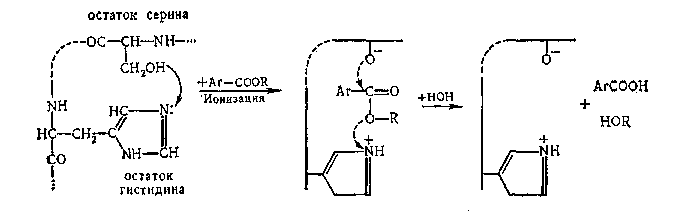
Действие ферментов очень чувствительно к конформации макромолекулы и надмолекулярной структуре его. Например, если в результате денатурации и выпрямления цепи б-химотрипсина активные группы значительно удалятся друг от друга, то «кооперативный эффект» исчезает и скорость гидролиза падает в миллион раз.
Большая активность ферментов обусловлена высоким значением предэкспонента А (эффект ориентации) в уравнении Аррениуса и низкой энергии активации («кооперативный эффект»).
3. Нуклеиновые кислоты, которые делятся на дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК) кислоты, были открыты в клеточном ядре (nucleus — ядро); РНК встречается также и в других частях клетки. Обе кислоты являются линейными полимерами, молекулярная цепь которых состоит из чередующихся остатков фосфорной кислоты и нуклеозидов. Нуклеозиды построены из звеньев циклической формы D-рибозы (РНК) или D-дезоксирибозы (ДНК) и остатков различных гетероциклических оснований, способных к попарному взаимодействию с образованием водородных мостиков. У РНК такие основания – аденин (А), цитозин (Ц), гуанин (Г) и урацил, у ДНК – аденин, цитозин, гуанин и тимин (Т):
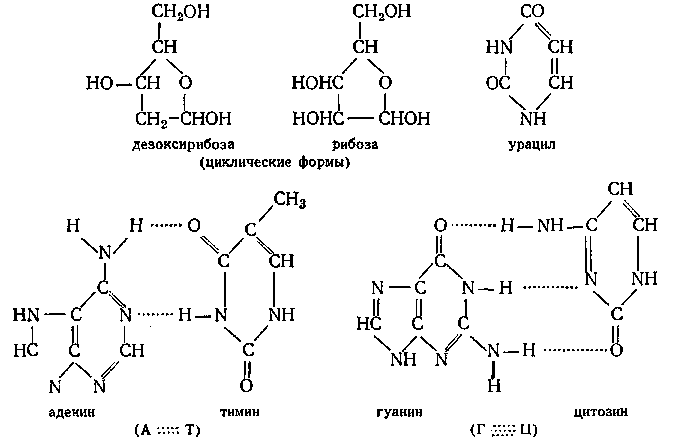
Как видно из формул, пара Г — Ц может образовать три водородных мостика, а пара А — Т — только два.
Так как рибоза (дезоксирибоза) соединяется с органическими основаниями за счет аминного водорода и глюкозидного гидроксила, нуклеозиды должны быть отнесены к N-глюкозидам, где роль агликона выполняют упомянутые основания
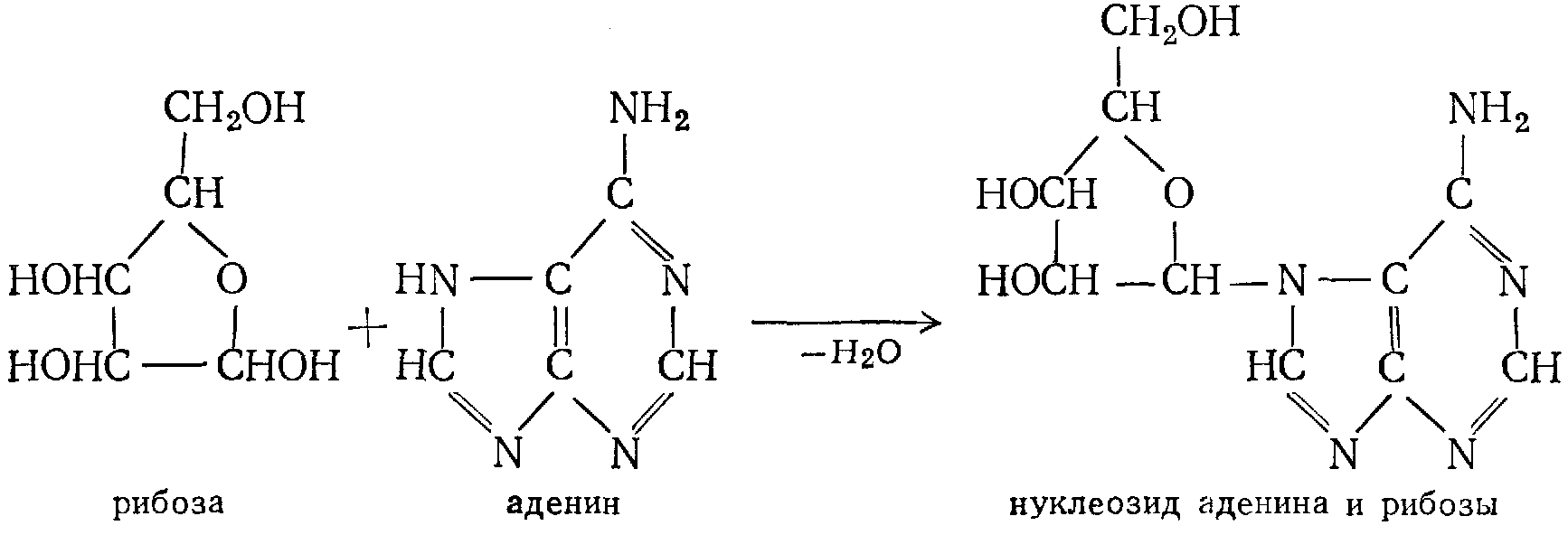
РНК реагирует с гидроксильными группами углеводов; при гидролизе РНК наряду с рибозой и соответствующими основаниями образуется фосфорная кислота. Аналогичные результаты – при гидролизе ДНК.
Если условно обозначить основания через
 , то строение ДНК можно изобразить следующим образом:
, то строение ДНК можно изобразить следующим образом:
или схематически

ДНК имеет молекулярный вес порядка 107 и в условиях клетки нерастворима. Существует несколько разновидностей РНК, среди которых наиболее активными в биосинтезе белка являются:
- растворимая, или транспортная, РНК (т-РНК) с Мr около 25 000;
- информационная РНК (и-РНК) с молекулярным весом порядка 6105-106.
ДНК является материальным носителем наследственности и входит в состав генов, из которых состоят хромосомы клетки. Макромолекулы ДНК связаны между собой попарно при помощи водородных мостиков в виде двойной спирали постоянного диаметра (рис. 13). При этом остатки гетероциклических оснований, находящиеся в боковой цепи, упакованы в середине спирали, как стопка монет. Аналогичную структуру имеет РНК.
Для обеспечения наибольшей устойчивости этой структуры необходимо, чтобы число водородных связей было максимально возможным. Это достигается тем, что соблюдается определенное соответствие в расположении остатков оснований одной спирали по отношению к остаткам другой: тиминовые группы располагаются напротив адениновых, цитозиновые напротив гуаниновых и т. д. («узнают» друг друга).
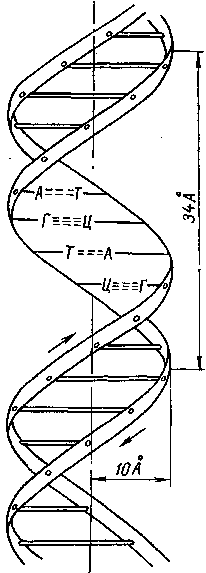
Рис. 13. Двойная спираль ДНК (по Крику и Уотсону)
Кроме того, только при выполнении этого условия будет обеспечено экспериментально доказанное постоянство суммарных размеров, боковых групп и неизменность диаметра двойной спирали на всем ее протяжении. В этой взаимной обусловленности порядка чередования звеньев в обеих цепях заключается принцип комплементарности, благодаря чему каждая цепь определяет структуру другой, являясь как бы ее репликой.
Спирализация приводит к возникновению так называемой «вторичной» структуры ДНК; при изгибании спирали появляется «третичная» структура и т. д. Возникновение изогнутой спирали, обусловлено наличием в спирали неупорядоченных гибких участков, где действие водородных связей почему-либо ослаблено. Однако двойная спираль — там, где она сохранилась — является достаточно жестким образованием и, следовательно, обладает небольшим числом степеней свободы. Поэтому она стремится разделиться на одиночные цепи (длина сегмента примерно в 50 раз больше, чем у гибких полимерных цепей), способные принять более вероятное состояние свернутого клубка; такой переход «спираль — клубок» сопровождается возрастанием энтропии системы, являющимся движущей силой этого процесса, и действительно имеет место при плавлении кристаллов ДНК (около 80°С). Аналогичный процесс разрушения водородных мостиков и биспиральной структуры, но без свертывания цепей в клубок, наблюдается во время подкисления или подщелачивания растворов ДНК. При этом на каждой макромолекуле возникают одноименные заряды (в результате присоединения протонов к аминогруппам или усиления диссоциации остатков фосфорной кислоты), вызывающие взаимное отталкивание цепей.
Если двойная спираль находится в растворе, содержащем большое количество различных нуклеотидов, т. е. нуклеозидов, химически связанных с фосфорной кислотой, то в результате замены водородных связей между цепями такими же связями их с нуклеотидами произойдет сорбция последних и одновременное разделение спирали на две самостоятельные макромолекулы (рис. 14). При этом нуклеотиды отбираются так, чтобы соблюдался принцип комплементарности; во время дальнейшей поликонденсации, протекающей под влиянием ферментов, на каждой цепи ДНК вырастает другая, являющаяся ее репликой. Этот процесс редупликации (копирования) макромолекул приводит к возникновению новых «дочерних» двойных спиралей.
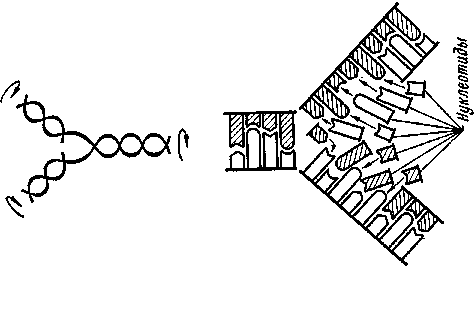
Рис. 14. Схема редупликации ДНК
Двойная спираль ДНК не только является матрицей воспроизведения самой себя, она также передает информацию, «записанную» в ее структуре, нуклеотидам, участвующим в синтезе РНК, тем самым предопределяет порядок их расположения в образующейся макромолекуле. Информационная РНК, на каждой цепи которой запечатлена структура молекулы определенного белка, играет ту же роль при образовании полипептидной цепи. В этом случае процесс осложняется тем, что в и-РНК имеется всего 4 нуклеотидных звена, а в белке – 20 аминокислотных. Поэтому для фиксации положения каждого аминокислотного остатка требуется не меньше трех нуклеотидных. Иначе говоря, химическое строение макромолекулы белка кодируется структурой и-РНК подобно тому, как кодируется текст с помощью азбуки Морзе.
С другой стороны, отсутствие структурного соответствия между суммарными размерами трех нуклеотидных звеньев (21 А) и величиной одного аминокислотного (3,6 А) исключает образование из них промежуточного комплекса, без которого невозможен синтез полипептидной цепи на матрице и-РНК. По мнению Крика, промежуточный комплекс все же получается, но более сложным путем, с участием т-РНК (адаптор). При этом один конец макромолекулы т-РНК избирательно образует лабильную связь с остатком определенной аминокислоты («узнает» ее) за счет реакции АМФ — АК с группой ОН рибозы:
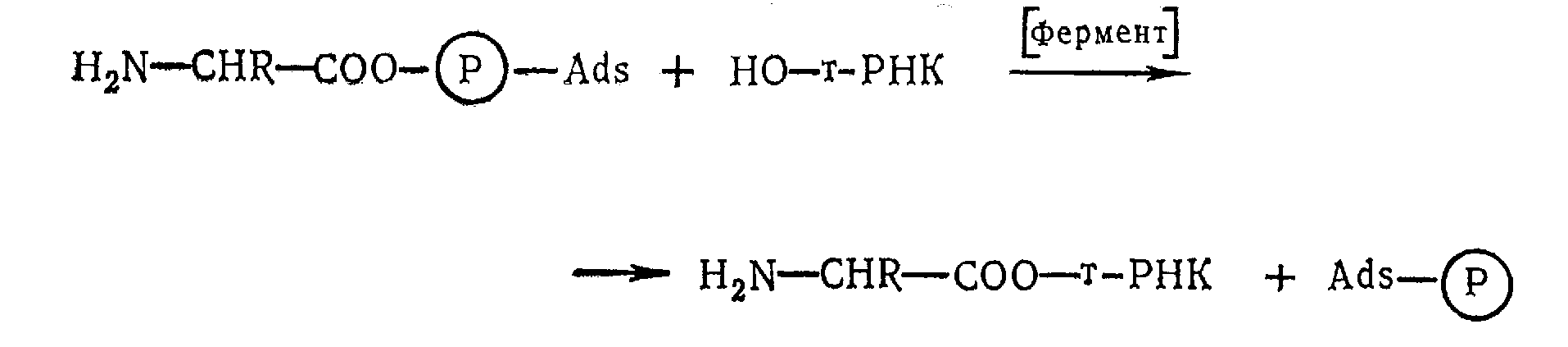
а второй присоединяет триплет нуклеотидов (кодон), который комплиментарен трем нуклеотидам, находящимся на этом конце т-РНК. В соответствии с адапторной гипотезой Крика, в синтезе белка участвует 20 специфических т-РНК (по одной на каждую аминокислоту). Решающую роль в выборе аминокислоты играет селективный фермент, обладающий сродством к боковой группе (R) этой кислоты и к ее адаптеру.
Образовавшиеся АК — т-РНК затем диффундируют к рибосомам, которые ориентируют их относительно одноцепочной и-РНК таким образом, чтобы обеспечить точное «считывание» генетического кода, т. е. строго определенную последовательность остатков аминокислот. В дальнейшем расщепляется богатая энергией связь АК — т-РНК с возникновением энергетически бедной пептидной связи:
H2N—CHR'—СОО—т-РНК' + Н2N—CHR"—COO—т-РНК" →
→ H2N—CHR'—CO—NH—CHR"—COO—РНК" + т-РНК'
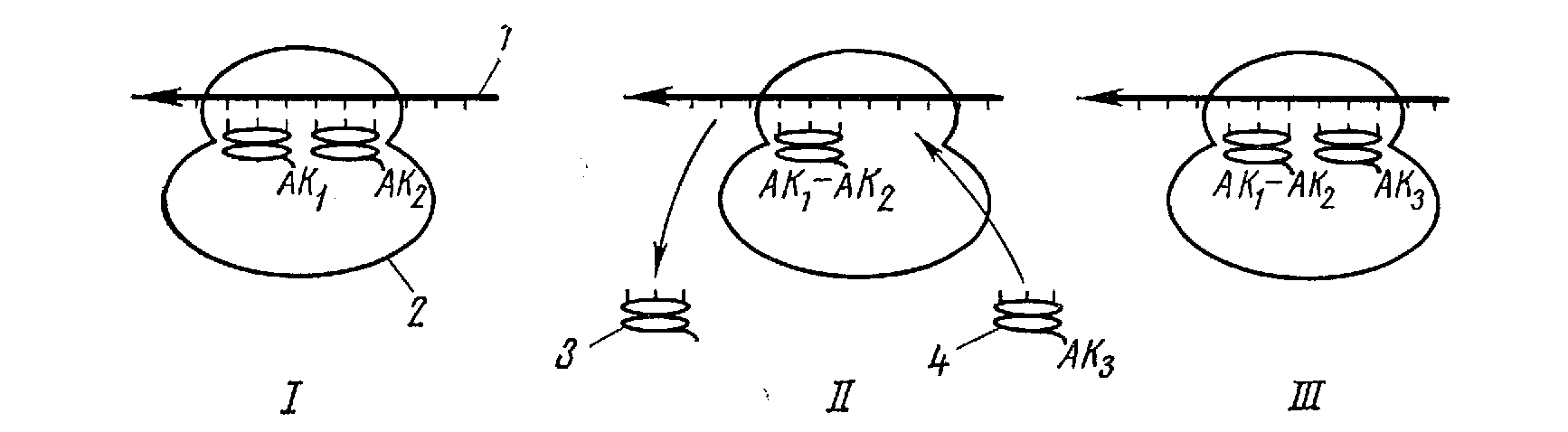
Рис. 15. Схематическое изображение этапов биосинтеза белка: 1 — и-РНК; 2 — рибосома; 3—т-РНК; 4 — комплекс т-РНК — аминокислота
4. На рис. 15 в схематическом виде изображен один из этапов роста полипептидной цепи [69, 70]. В результате присоединения новых АК — т-РНК и движения рибосом (точнее, полирибосом) по цепи и-РНК образуется белковая макромолекула, которая затем отделяется от матрицы и-РНК.
Биосинтез белка сопровождается уменьшением свободной энергии, несмотря на то, что F образования пептидной связи из самих аминокислот больше нуля; объясняется это тем, что пирофосфатная связь АТФ поставляет недостающую энергию смешанному ангидриду и комплексу АК — т-РНК, являющимся как бы активированными предшественниками синтеза белка.
Можно осуществить синтез полипептидов, сходных по химическому строению с белками, в лабораторных условиях; для этого сначала отщепляют воду от двух молекул аминокислот, предварительно «защищая» аминогруппу одной из них и карбоксильную группу другой:
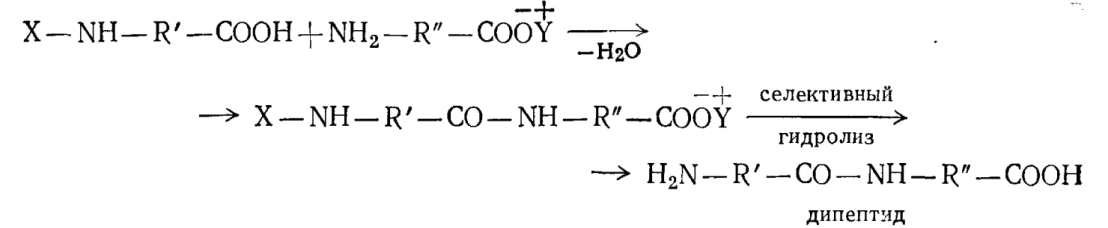
где Х – защитная группа, например n-NO2–C6H4–S– , а Y+ — протонизированный триэтиламин, HN+(C2H5)3.
Аналогично из дипептида синтезируют трипептид, из него — тетрапептид и т. д., получая в конечном итоге полипептиды со строго определенным чередованием аминокислотных остатков. Во всех этих реакциях дегидратирующим агентом обычно служат диимиды:
C6H11—N=C=N—С6Н11 + Н2О → С6Н11NHCONHC6H11
Принципиально новым подходом к этому вопросу является твердофазный пептидный синтез, где растущая цепь все время химически связана со стороны группы СООН с таким трехмерным полимером, как хлорметилированный сополимер стирола и дивинилбензола, подвергнутый еще нитрованию (полимер обозначен через П):
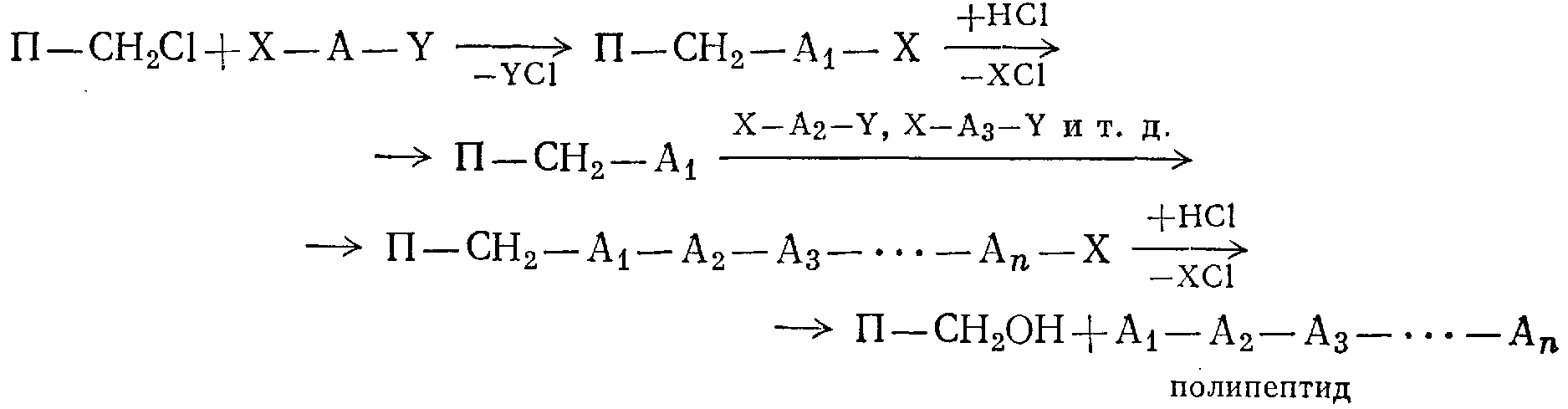
(здесь А1 А2, . . ., Аn — остатки различных аминокислот).
Вследствие нерастворимости прикрепленного к сетчатому полимеру полипептида упрощается его очистка, повышается выход (при обычном методе синтеза инсулина, состоящего из 221 стадии, суммарный выход ничтожно мал; новый метод дает выход до 68%) и практически исключается рацемизация. Новый метод может быть автоматизирован, и с некоторыми изменениями он пригоден для синтеза полисахаридов и полинуклеотидов.
При воспроизведении ДНК возможно появление «ошибочных» звеньев, например вследствие перехода остатка тимина в енольную форму:
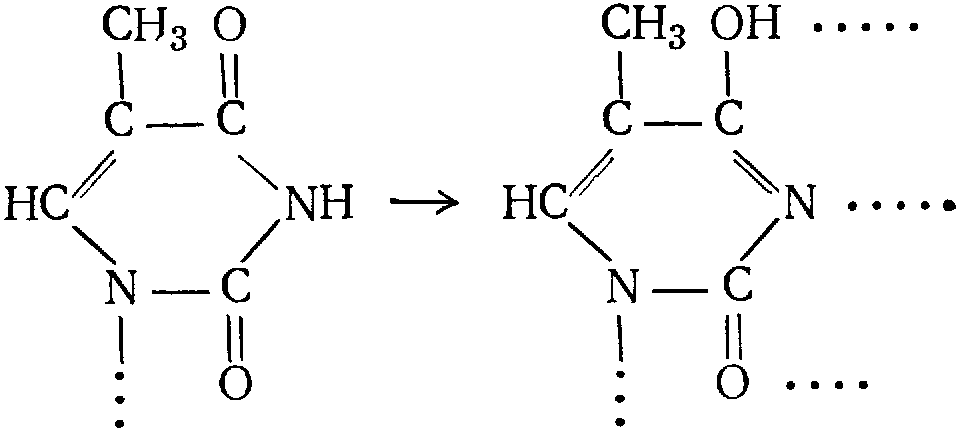
Т.к. новая форма в отличие от исходной способна образовывать три водородные связи вместо двух, «дочерняя» двойная спираль будет содержать «неправильные» пары. Этот эффект может быть вызван радиоактивным облучением или замещением аминогруппы органического основания на группу ОН: R—NH2 + HO—NO → R—OH + N2 + H2O.
Даже незначительные «повреждения» в макромолекуле ДНК имеют большое значение, так как они носят наследственный характер и могут передаваться от ДНК к РНК и от РНК к аминокислотам. В результате изменится не только весь ход биосинтеза в клетке, но также свойства, ферментативная активность и сама природа образующихся белков [54-56,69, 70].