
Природа мембранного потенциала покоя
| Вид материала | Документы |
- 2 Современные представления о генерации мембранного потенциала покоя, 153.42kb.
- Міністерство охорони здоров’я України Український центр наукової медичної інформації, 290.67kb.
- Конкурсе методических разработок Тема: «Природа источник познания», 20.22kb.
- Медицинский портал студентам, врачам, медицинские книги, 246.39kb.
- Уроки з курсу “Я І Україна”, 590.67kb.
- М. Ю. Лермонтова Вцелом ряде произведений М. Ю. Лермонтова храм является значимым пространством, 252.48kb.
- 1. Дифференциальный каскад (ДК) на биполярных транзисторах: схема, режим покоя каскада, 54.47kb.
- Розділ 3 Економіка підприємства та організація виробництва, 221.25kb.
- План управления (менеджмент-план) природного парка «зона покоя укок» на 2009-2013, 3525.68kb.
- Задачи: углубить общее представление о строении эукариотической клетки на основе раскрытия, 356.11kb.
Комплекс потенциалов мозга, связанных с движениями
Одним из важных направлений в исследовании психофизиологии двигательного акта является изучение комплекса колебаний потенциалов мозга, связанных с движениями (ПМСД). Значение этого феномена для понимания физиологических механизмов организации движения очень велико, потому что изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и хронометрировать эти процессы, т.е. установить временные границы их протекания.
Компонентный состав ПМСД. Впервые этот комплекс, отражающий процессы подготовки, выполнения и оценки движения был зарегистрирован в 60-е годы. Оказалось, что движению предшествует медленное отрицательное колебание — потенциал готовности (ПГ). Он начинает развиваться за 1,5 - 0,5 с до начала движения. Этот компонент регистрируется преимущественно в центральных и лобно-центральных отведениях обоих полушарий. За 500-300 мс до начала движения ПГ становится асимметричным — его максимальная амплитуда наблюдается в прецентральной области, контралатеральной движению. Примерно у половины взрослых испытуемых на фоне этого медленного отрицательного колебания незадолго до начала движения регистрируется небольшой по амплитуде положительный компонент. Он получил название "премоторная позитивность" (ПМП). Следующее по порядку быстро нарастающее по амплитуде отрицательное колебание, так называемый моторный потенциал (МП), начинает развиваться за 150 мс до начала движения и достигает максимальной амплитуды над областью моторного представительства движущейся конечности в коре головного мозга. Завершается этот комплекс потенциалов положительным компонентом примерно через 200 мс после начала движения.
Функциональное значение компонентов. Принято считать, что потенциал готовности (ПГ) возникает в моторной коре и связан с процессами планирования и подготовки движения. Он относится к классу медленных отрицательных колебаний потенциала мозга, возникновение которых объясняют активацией нейрональных элементов соответствующих участков коры.
Гипотезы относительно функционального значения ПМП различны.
Это колебание рассматривают и как отражение подачи центральной команды от коры к мышцам, и как результат релаксации коры после завершения определенного этапа организации движения, и как отражение процессов подавления ассоциированных движений другой конечности, и как обратную связь от мышечных афферентов. В настоящее время некоторые авторы полагают, что ПМП являются лишь отражением начала моторного потенциала.
При регистрации МП у обезьян в составе МП были выделены два субкомпонента. Первый субкомпонент соотносят с активацией моторной коры, связанной с инициацией движения (синаптическая активность пирамидных нейронов), а второй — с активацией полей 2.3 и 4 по Бродману. Регистрация МП у человека, больного эпилепсией, позволила выделить в нем три компонента. Первый компонент был назван потенциалом инициации. Он имеет высокую амплитуду и возникает после начала движения в прецентральной контралатеральной коре. Второй, возникающий после начала миограммы, более локализованный в контралатеральном соматосенсорном поле может быть связан как с инициацией движения, так и с сенсорной обратной связью. Третий компонент отражает импульсацию, поступающую с мышечных афферентов в кору.
Следующий за МП позитивный потенциал рассматривается как отражение обратной афферентации, поступающей с периферических рецепторов, восходящей активности от моторных центров, операции сравнения между моторной программой и нейрональной картиной ее исполнения или процессов релаксации коры после выполнения движения
Вопрос 32. Связанные с событиями потенциалы.
Сенсорные стимулы вызывают изменения в суммарной электрической активности мозга, которые выглядят как пocледовательность из нескольких позитивных и негвтивных волн, которая длится в течение 0,5-1 с после стимула. Этот ответ получил название вызванного потенциала (evoked pоtential). Его нелегко выделить из фоновой ЭЭГ. В 195I г. Дж. Даусон (G. Dаwson) разработал технику когерентного накопления или усреднения ответов. Использовалась процедура синхронизации ЭЭГ относительно момента предъявления стимула который поэтому многократно повторялся. Сначала использовалась суперпозиция — наложение, нескольких реакций (участков ЭЭГ, следующих за стимулом). Обычно это выполнялось на фотопленке, чтo позволяло выявить наиболее устойчивые части реакции на стимул. Затем процедура суперпозиции была заменена на суммацию участков ЭЭГ и получение усредненного вызванного потенциала (average evoked рotеntial) (Шагас Ч., 1975; Рутман Э.М., 1975).
Эффективность этой процедуры была наглядно продемонстрирована при выявлении звуковых стволовых вызванных потенциалов (ВП). Из-за их очень малой амплитуды требуется просуммировать и усреднить несколько тысяч единичных ответов. На рис. 2 пpeдставлены основные группы компонентов звукового усредненного ВП. По латентному периоду компоненты делятся на три группы: потенциалы ствола мозга (с латенцией до 10-12 мс), средне-латентные (до 50 мс) и длиннолатентные (болеe 100 мс) потенциалы. Звуковые стволовые потенциалы состоят из 7 отклонений. Волна I зависит oт реакции волокон слухового нерва улитки. Волна II с латенцией 3,8 мс возникает в том случае, если импульсы слухового нерва достигают ствола мозга. Волна III отражает реакцию верхней оливы на уровне моста. Волна IV с латенцией около 4,5 мс связана с активностью латеральных лемнисков. Волна V имеет латенцию около 5,2 мc и отражает активность нижнего двухолмия. Фазы VI— VII — распространение сигналов по таламо-кортикальной радиации, они совпадают с медленной негативностью, предшествующей корковому ответу. Ранние компоненты нечувствительны к сну, наркозу. Они вызываются звуковыми тонами частотой 2000-4000 Гц. Звуки ни частоте, ниже 2000 Гц вызывают только волну V.
Cтволовые потенциалы — высокочувствительный инструмент для тестирования слуховой функции. Они позволяют определить сохранность слухового анализатора на периферическом и стволовом уровнях. Особенно это важно при обследовании слуха у детей, в том числе у новорожденных, когда словесные реакции не могут быть использованы. Значение этого теста возрастает в связи с тем фактором, что даже незначительная потеря слуха в раннем детстве может привести к существенной задержке развития речи. Стволовые звуковые потенциалы применяют также в клинике для выявления опухолей, определения коматозного состояния, обследования пациентов с демиелинизацией волокон. Если стволовые потенциалы полностью oтсутствуют, можно говорить о смерти мозга.
Позже техника усреднения ВП была применена для выявления потенциалов, связанных с движением. Участки ЭЭГ усреднялись относительно не стимула, а начала движения. Это дало возможность исследовать моторные потенциалы и потенциалы готовности, предшествующие движению. Для обозначения всех групп потенциалов был введен общий для них термин — «потенциалы, связанные с событиями» (ПСС), объединяющий ВП, моторный потенциал и др.
На основе многоканальной регистрации ЭЭГ был разработан метод картирования биотоков мозга (brain mapping). Картирование дает представление о пространственном распределении по коре любого выбранного показателя электрической активности мозга. Это может быть ВП, один из его компонентов или альфа-ритм (или другие частотные полосы спектра ЭЭГ). Значения мощности выбранного показателя подразделяются на уровни. В одном варианте каждому уровню приписывается свой цвет и изменение локуса активности выглядит как перемещение определенного цвета по карте. В другом варианте значения показателя, принадлежащие одному уровню, соединяются изолиниями, как на топографических картах, на которых можно видеть возвышенности и впадины. Рассматриваются карты, полученные в разное время и в разных условиях. Этот метод позволяет выявить фокусы активности мозга. Используется процедура вычитания одной карты потенциалов из другой, что позволяет связать паттерн ЭЭГ-активности с той или другой когнитивной операцией.
№ 33. Таламический водитель ритма
Клетки-пейсмекеры (водители ритма) – особая разновидность клеток, способных генерировать потенциал действия самостоятельно, без раздражения извне. Деятельность этих клеток нарушает общий закон о необходимости первопричины движения. Пейсмекеры расположены по всему организму человека – в т.ч., в таламических структурах мозга и в сердце. Обращаю внимание, что в тексте пойдёт речь не о таламических пейсмекерах, а о функции таламуса, которая будет сравниваться с функцией пейсмекеров сердца. Так что воспринимайте формулировку данного вопроса как метафору
Структуры головного мозга (ГМ), которые отвечают за биологические мотивационные состояния, обладают различными свойствами.
Разрушение гипоталамических центров полностью исключает биологические мотивации – животное может лежать в окружении пищи и погибнуть от голода. Т.е., в структуре мотивационного возбуждения гипоталамическим центрам принадлежит ведущая роль.
Гипоталамические центры в структуре «голодного» мотивационного возбуждения гораздо чувствительнее к химическим веществам по сравнению с корой ГМ. Раствор атропина
0, 0001%, введённый через специальную иглу в латеральный гипоталамус, устраняет голодную мотивацию во всех структурах ГМ.
Механизм формирования биологической мотивации у животных напоминает возникновение возбуждения в синусоидном узле сердечной мышцы, где располагаются клетки-пейсмекеры.
- В сердце возбуждение возникает ритмически. Та же картина – и в мотивациогенных центрах гипоталамуса. Возбуждение там возникает периодически, по триггерному типу, по мере нарастания потребности до критического уровня. Оно сохраняется, пока существует потребность, и затем исчезает.
- Как и пейсмекер сердца, структуры гипоталамуса по сравнению с другими структурами ГМ, обладают повышенной возбудимостью к электрическим и химическим раздражителям.
- Так же как и в сердечной мышце, по отношению к гипоталамическим образованиям, другие структуры мозга выстраиваются по определённому градиенту возбудимости до коры включительно.
- Мотивационные центры гипоталамуса держат в функциональной зависимости структуры других уровней мозга. Выключение этих центров приводит к распаду всей системы объединённых в мотивационное возбуждение элементов.
Концепция пейсмекерной роли гипоталамических центров в формировании основных биологических мотиваций принадлежит Анохину и Судакову. Знание этой роли определяет врачебную тактику влияния на них с помощью препаратов.
№ 34. Роль ретикулярной формации в поддержании активности коры ГМ.
Ретикулярная формация - совокупность нейронов и соединяющих их нервных
волокон, расположенных в стволе мозга и образующих сеть. Ретикулярная формация простирается через весь ствол головного мозга от верхних шейных спинальных сегментов до промежуточного мозга. Анатомически она может быть разделена на ретикулярную формацию продолговатого мозга, варолиевого моста и среднего мозга. Вместе с тем, в функциональном отношении в ретикулярной формации разных отделов мозгового ствола есть много общего. Поэтому целесообразно рассматривать ее как единую структуру. Ретикулярная формация представляет собой сложное скопление нервных
клеток, характеризующихся обширно разветвленным дендритным деревом и
длинными аксонами, часть из которых имеет нисходящее направление и образует
ретикулоспинальные пути, а часть восходящее. В ретикулярную формацию
поступает большое количество путей из других мозговых структур. С одной
стороны, это коллатерали волокон, проходящих через ствол мозга сенсорных
восходящих систем, - эти коллатерали заканчиваются синапсами на дендритах и
соме нейронов ретикулярной формации. С другой стороны, нисходящие пути,
идущие из передних отделов мозга (в том числе, пирамидный путь), тоже дают
большое количество коллатералей, которые входят в ретикулярную формацию и
вступают в синаптические соединения с ее нейронами. Обилие волокон
поступает к нейронам ретикулярной формации из мозжечка. Таким образом, по
организации своих афферентных связей эта система приспособлена к
объединению влияний из различных мозговых структур.
Данные о том, что ретикулярная формация играет важную роль в поддержании
нормальной деятельности коры больших полушарий, были получены еще в
тридцатые годы нашего столетия. Бельгийский нейрофизиолог Ф. Бремер (1935), проводя
перерезку головного мозга на различных уровнях и наблюдая за
функциями отделенных от остальной центральной нервной системы участков
головного мозга, обратил внимание на то, что имеется чрезвычайно
существенное различие между животным, у которого перерезка проведена по
между передними и задними холмами
четверохолмия, и животным, у которого линия разреза прошла между
продолговатым и спинным мозгом. Первый препарат был назван Бремером cerveau isolee, а второй encephale
isolee (изолированный передний мозг и изолированный головной мозг).
Первая перерезка оставляет ниже места разреза почти всю
ретикулярную формацию; перерезка же ниже продолговатого мозга оставляет в
связи с высшими отделами мозга все стволовые структуры. У животного с
ceryeau isolee остаются связанными с корой головного мозга такие важнейшие
афферентные системы, как обонятельная и зрительная. Однако такое животное
не реагирует на световые и обонятельные раздражения; глаза у него закрыты,
и оно фактически не вступает в контакт с внешним миром. У животного же с
encephale isolee реакции полностью сохраняются; такое животное открывает
глаза на свет, реагирует на запахи, т.е. ведет себя как бодрствующее, а не
как спящее животное.
Бремер сделал вывод, что для бодрствующего состояния высших отделов мозга необходимо непрерывное поступление к ним афферентных импульсов, в
частности, от такой обширной рецептивной зоны, как зона тройничного нерва.
Предполагалось, что эта импульсация поступает в кору больших полушарий по
восходящим афферентным путям и поддерживает высокую возбудимость ее
нейронов. Однако дальнейшие исследования показали, что для поддержания
бодрствующего состояния коры больших полушарий важно не просто поступление
к ней импульсации по афферентным системам. Если сделать перерезку ствола
мозга так, чтобы не повредить основные афферентные системы (например,
систему медиальной петли), но перерезать восходящие связи ретикулярной
формации, то животное тем не менее впадает в сонное состояние, конечный
мозг перестает активно функционировать. Следовательно, для поддержания бодрствующего состояния конечного мозга
важно, чтобы афферентная импульсация первоначально активировала
ретикулярные структуры мозгового ствола. Влияния же ретикулярных
структур по восходящим путям каким-то образом определяют функциональное
состояние конечного мозга. Проверить такой вывод можно путем прямого
раздражения ретикулярных структур. Такое раздражение было проведено Моруцци и Мегоуном.
Оно всегда дает однозначные результаты в виде характерной поведенческой
реакции животного. Если животное находится в сонном состоянии, оно
просыпается, у него появляется ориентировочная реакция. После прекращения
раздражения животное снова возвращается в сонное состояние. Переход от
сонного к бодрствующему состоянию в период раздражения ретикулярных
структур отчетливо проявляется не только в поведенческих реакциях, он может
быть зарегистрирован по объективным критериям деятельности коры больших
полушарий, в первую очередь по изменениям ее электрической активности.
Для коры больших полушарий характерна постоянная электрическая
активность. Эта электрическая активность состоит из небольших по амплитуде (30-100 мкВ) колебаний, которые легко отводятся не только от открытой поверхности мозга, но и от кожи головы. У человека в спокойном дремотном состоянии такие колебания
имеют частоту 8-10 в секунду и являются довольно регулярными (альфа-ритм).
Во время активности регулярные колебания сменяются сразу же значительно меньшими по амплитуде и более высокочастотными колебаниями (бета-ритм). Когда
правильные колебания большой амплитуды сменяются низковольтными, частыми
колебаниями, то это говорит о том, что клеточные элементы коры
начинают функционировать менее синхронно, поэтому такой тип активности
называется реакцией десинхронизации. Таким образом, переход от спокойного,
неактивного состояния коры к активному связан в электрическом отношении с
переходом от синхронизированной активности ее клеток к
десинхронизированной. Характерным эффектом восходящих ретикулярных влияний на корковую электрическую активность является именно реакция десинхронизации.
Реакция десинхронизации не ограничивается каким-нибудь одним участком коры, а регистрируется от больших ее областей. Это говорит о том, что восходящие ретикулярные влияния являются генерализованными.
Описанные изменения в электроэнцефалограмме не являются единственным
электрическим проявлением восходящих ретикулярных влияний. В определенных
условиях можно выявить более прямые эффекты ретикулярной импульсации,
поступающей к коре больших полушарий. Они впервые были описаны в 1940 г.
американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали
вызванную электрическую активность коры при различных афферентных влияниях.
При раздражении какой-либо афферентной системы, в соответствующей
проекционной зоне коры обнаруживается электрический ответ, указывающий на
приход к этой области афферентной волны, этот ответ получил название
первичного ответа. Кроме этого локального ответа, афферентное раздражение
вызывает длиннолатентный ответ, возникающий в обширных областях коры мозга.
Этот ответ Форбс и Мориссон назвали вторичным ответом. То обстоятельство, что вторичные ответы возникают со скрытым периодом,
значительно превышающим скрытый период первичного ответа, ясно говорит о
том, что они связаны с поступлением афферентной волны в кору не по прямым,
а по каким-то окольным связям, через дополнительные синаптические
переключения. Позже, когда было применено прямое раздражение ретикулярной
формации, было показано, что оно может вызвать ответ такого же типа. Это
позволяет заключить, что вторичный ответ является электрическим проявлением
поступления в кору больших полушарий афферентное волны через
ретикулокортикальные связи. Через ствол мозга проходят прямые афферентные пути, которые после
синаптического перерыва в таламусе поступают в кору больших полушарий.
Приходящая по ним афферентная волна вызывает первичный электрический ответ
в соответствующей проекционной зоне с коротким скрытым периодом.
Одновременно афферентная волна по коллатералям ответвляется в ретикулярную
формацию и активирует ее нейроны. Затем по восходящим путям от нейронов
ретикулярной формации импульсация тоже поступает в кору, но уже в виде
задержанной реакции, возникающей с большим скрытым периодом. Эта реакция
охватывает не только проекционную зону, но и большие участки коры, вызывая
в них какие-то изменения, важные для бодрствующего состояния.
Швейцарский физиолог В. Гесс (1929) впервые показал, что в стволе
мозга можно найти точки, при раздражении которых у животного наступает сон.
Гесс назвал эти точки центрами сна. Позже Моруцци и сотр. (1941) также
обнаружили, что, раздражая некоторые участки ретикулярной формации заднего
мозга, можно у животных вызвать в коре вместо десинхронизации синхронизацию
электрических колебаний и, соответственно, перевести животное из
бодрствующего состояния в пассивное, сонное. Поэтому можно думать, что в
составе восходящих путей ретикулярной формации действительно существуют не
только активирующие, но и инактивирующие подразделения, последние каким-то
образом снижают возбудимость нейронов конечного мозга.
35. Гипотезы о природе суммарной биоэлектрической активности мозга.
Возникновением различных форм биоэлектрической активности сопровождается любой акт жизнедеятельности – мышечное сокращение, деятельность сердца, а также деятельность мозга. Регистрация Б.Э.А. возможна с помощью спец. аппаратуры. В частности, при измерении Б.Э.А. мозга мы получаем электроэнцефалограмму мозга.
Было выдвинуто несколько гипотез о природе суммарной Б.Э.А.:
- Изначально считали, что Б.Э.А. мозга создается суммой потенциалов действия клетки. Но от этой теории отказались.
- Когда было выяснено, что эл. сигнал от нейрона передается через синапс, то было выдвинуто предположение, что Б.Э.А. – это сумма постсинаптических потенциалов. Это мнение и господствует в настоящее время.
- Также высказывалось предположение, что Б.Э.А. – это сумма потенциалов галиальных клеток, но эта гипотеза пока не подтвержена.
Билет 37. Развитие нервной системы в онтогенезе.
Онтогенез – процесс индивидуального развития особи (с момента оплодотворения до смерти организма).
Первоначальные деления зиготы приводят к формированию бластулы – зародыша (в виде полого шара), стенка которого образована одним слоем клеток. Затем в результате перемещения клеток бастулы превращается в двуслойную гаструлу. Возникающие при этом слои: эктодерма-наружный и энтодерма-внутренний, называют зародышевыми листками. В дальнейшем образуется третий – мезодерма. Нервная ткань за исключением клеток микроглии образуется из эктодермы.
Процесс, посредством которого часть клеток эктодермы превращается в специализированную нервную ткань, из которой позже развиваются спинной и головной мозг, называется пейруляцией или нейтральной индукцией. Определяющее событие для нейруляции – взаимодействие эктодермы и мезодермы. Индуцирующее действие оказывают также элементы хорды (основы будущего позвоночника).
В конце второй недели зародышевого развития участок эктодермы на дорсальной (спинной) стороне зародыша утолщается, образуя нервную пластинку. Клетки ее интенсивно размножаются и дифференцируются, приобретая узкую цилиндрическую форму. В результате интенсивного деления и неравномерного роста края нервной пластинки поднимаются, образуя нервные валики, между которыми лежит нервная бороздка.
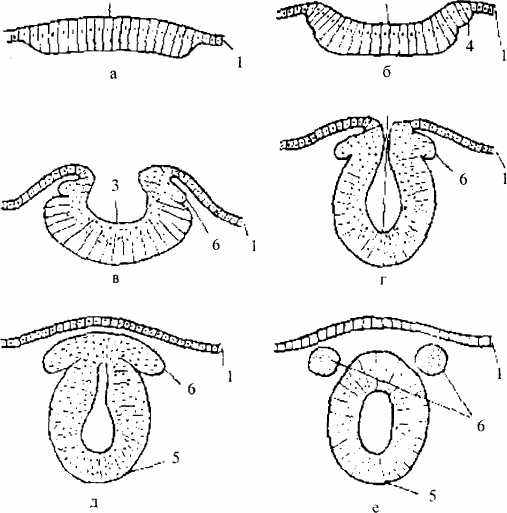
Рис. 1 /. Начальный период развития нервной системы
(поперечный разрез):
а - е — последовательные стадии развития: 1 — эктодерма:
2 — нервная пластинка; 3 — нервная бороздка; 4 — нервные вачики;
5— нервная трубка; 6— ганглиозная пластинка
Затем края нервной бороздки смыкаются, образуя нервную трубку. Она постепенно погружается в мезодерму и замыкается — сначала посередине, позже (к концу 4 недели развития) — на переднем и заднем концах. От нервной трубки по обеим ее сторонам отделяется тяж клеток — так называемая ганглиозная пластинка (нервный гребень). Ее клетки впоследствии дают элементы периферической нервной системы, паутинной и мягкой мозговых оболочек. Оставшаяся нервная трубка состоит из одного слоя клеток— нейроэпителия. В дальнейшем в результате деления этих клеток образуется слой эпендимы и слой, дающий начало нейробластам и спонгиобластам. Из нейробластов образуются нервные клетки, из спонгиобластов — глиальные клетки. Нейробласты с определенного момента прекращают делиться и начинают образовывать отростки, т.е. постепенно принимают вид зрелого нейрона. Вслед за ростом отростков следует их миелинизация и образование синапсов.
В конце 3 недели развития ростральный (головной) конец нервной трубки преобразуется в мешковидное расширение, дающее начало головному мозгу, а каудальный (хвостовой) отдел дает начало спинному мозгу. Головной конец нервной трубки скоро подразделяется на три расширения — три первичных мозговых пузыря (стадия трех мозговых пузырей). Полости этих пузырей, несколько изменяя форму, сохраняются во взрослом мозгу в виде мозговых желудочков и мозгового водопровода. Самым ростральным пузырем является первичный передний мозг, за ним следует первичный средний мозг, последний пузырь, за которым уже начинается спинной мозг, это первичный задний мозг.
На 2 месяце развития первый и третий первичные мозговые пузыри с помощью борозд разделяются, образуя каждый по два вторичных мозговых пузыря (стадия пяти мозговых пузырей). Первичный передний мозг делится на конечный мозг (полушария большого мозга и базальные ядра) и промежуточный мозг. С каждой стороны промежуточного мозга вырастает глазной пузырь, формирующий нервные элементы сетчатки глаза. Глазной бокал, образованный этим выростом, вызывает изменения в лежащей непосредственно над ним эктодерме, что приводит к возникновению из нее хрусталика. Первичный задний мозг разделяется на собственно задний мозг, включающий мозжечок и варолиев мост, и продолговатый мозг. Средний мозг сохраняется как единое целое.
Отдельные части нервной трубки растут с разной скоростью. В результате этого одновременно с формированием пяти мозговых пузырей образуются изгибы зачатка головного мозга (рис. 12). Сначала— основной изгиб в области среднего мозгового пузыря, обращенный выпуклостью дорсально; потом шейный изгиб на границе головного и спинного мозга, также выпуклостью дорсально; последний — мостовой изгиб в области заднего мозгового пузыря выпуклостью вентрально.
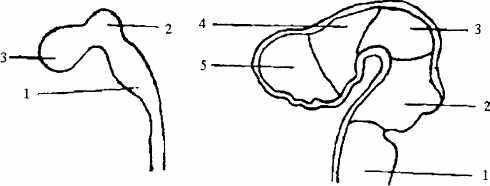
Рис. 12. Развитие головного мозга
А. Стадия трех мозговых пузырей:
1 — задний мозг\ 2 — средний мозг\ 3 — передний мозг.
Б. Стадия пяти мозговых пузырей:
1 — продолговатый мозг; 2 — мост и мозжечок; 3 — средний мозг; 4 — промежуточный мозг; 5— конечный мозг
После формирования мозговых пузырей в структурах ЦНС происходят сложные процессы внутренней дифференцировки и роста.
В возрасте 10-20 недель образуются все основные отделы нервной системы. К этому моменту заканчивается миграционный период ее развития, т.е все нейроны перемещаются туда, где они будут находиться во взрослом мозгу. Полушария постепенно становятся самой большой частью нервной системы, происходит выделение основных долей, начинается образование борозд и извилин. Из оболочек в ткань мозга врастают кровеносные сосуды. В спинном мозгу формируются шейное и поясничное утолщения. Окончательный вид приобретает мозжечок.
В последние месяцы эмбрионального развития в нервной системе заканчивается формирование внутренней структуры мозга (его ядер и трактов). Начинается активная миелинизация сначала спинного (в возрасте 20 недель), а затем (в возрасте 36 - 40 недель) головного мозга, которая в основном заканчивается только к 10 - 12 годам.
Мозг новорожденного весит примерно 350 г, т.е. 10% от всего веса тела. Вес мозга взрослого человека около 1250 - 1350 г. Так как деление подавляющего большинства нервных клеток прекращается еще до рождения, увеличение массы мозга происходит за счет роста тел нейронов и их отростков, миелинизации нервных волокон, деления и роста клеток нейроглии.
Таким образом, нервная система проходит следующие стадии развития:
- Стадия пролиферации – процесс деления нейробластов на внутренней стороне нервной трубки (деление нервных клеток происходит только внутриутробно)
- Стадия миграции (перемещение клеток в «свою часть» нервной системы)
незрелые нейроны расселяются и находят себе постоянное место.
- Стадия специализации
Превращается в нервную клетку, вырастает аксон, образуются синапсы.
Конус роста – дистальная уплощенная часть аксона, которая прокладывает путь через окружающую ткань (Конус роста также образовывается при травме аксона). В конусе роста содержится фактор роста нервов (ФРН) – это белок, способствующий росту и развитию аксона.
Билет 38. «Время рефлекса» и синаптическая задержка.
Время рефлекса – это длительность реакции от момента действия раздражителя до момента самого действия (=до начала реакции).
Время рефлекса складывается из:
- Длительности возбуждения сенсорных рецепторов
- Проведения возбуждения по афферентным волокнам
- Синаптической передачи сигнала
- Проведения возбуждения по эфферентным волокнам
- Самой реакции
Основное время рефлекса приходится на синаптическую передачу возбуждения -синаптическую задержку. Синаптической задержкой называется затрата времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постсинаптической мембране потенциалов. При относительно высокой скорости распространения импульса по аксону, через синапс сигнал идет медленно. В большинстве центральных нейронов синаптическая задержка составляет около 0.1-0.3 мс. После этого требуется еще время на развитие возбуждающего постсинаптического потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1.5 мс. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.
39. Конвергенция. Пространственная и временная суммация.
Конвергенция (возбуждения) – принцип общего конечного пути. Поступление возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип «воронки Шеррингтона»). Объясняется это наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС может располагаться до 10 000 синапсов, на мотонейронах – до 20 000 синапсов. Явление конвергенции возбуждения в ЦНС имеет широкое распространение. Примером может служить конвергенция возбуждений на спинальном мотонейроне. Так, к одному и тому же спинальному мотонейрону подходят первичные афферентные волокна (рис. 4.5, б), а также различные нисходящие пути многих вышележащих центров ствола мозга и других отделов ЦНС. Явление конвергенции весьма важно, т.к. оно обеспечивает, например, участие одного мотонейрона в нескольких различных реакциях. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чихания т дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг. На рис. 4.5, а показаны два афферентных волокна, каждое из которых отдает коллатерали к четырем нейронам таким образом, что три нейрона из общего их числа, равного пяти, образуют связи с обоими афферентными волокнами. На каждом из этих трех нейронов конвергируют два афферентных волокна.

Поскольку на один мотонейрон может конвергировать множество коллатералей аксонов, генерация потенциала действия (ПД) в каждый момент зависит от общей суммы возбуждающих и тормозящих синаптических влияний, ПД возникают лишь в том случае, когда преобладают возбуждающие влияния. Конвергенция может облегчать процесс возникновения возбуждения на общих нейронах в результате пространственной суммации подпороговых ВПСП (возбуждающих постсинаптических потенциалов), либо блокировать его вследствие преобладания тормозных влияний.
Суммация возбуждения в ЦНС. Одно из основных свойств нервных центров.
Это явление было открыто великим русским физиологом И.М.Сеченовым в 1868 г. в опыте на лягушке. Раздражение конечности лягушки слабыми редкими импульсами не вызывало реакцию, а более частые раздражения такими же слабыми импульсами сопровождались ответной реакцией – лягушка совершала прыжок.
Различают временную (последовательную) и пространственную суммацию (рис. 4.7, а, б).
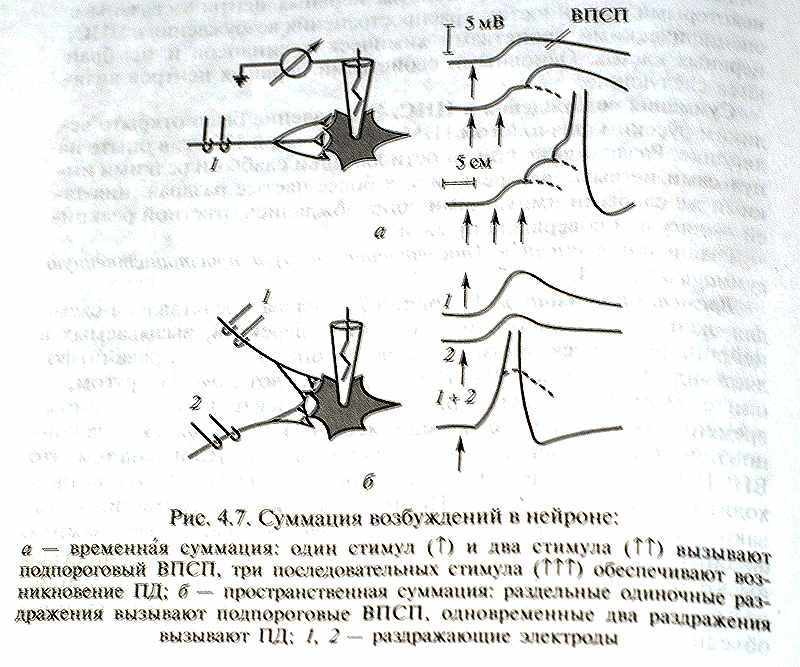
Временная суммация. На рис. 4.7, а слева представлена схема для экспериментального тестирования эффектов, вызываемых в нейроне ритмической стимуляцией аксона. Запись справа позволяет видеть, что если ВПСП быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временному ходу (несколько миллисекунд), достигая в конце концов порогового уровня. Временная суммация обусловлена тем, что ВПСП от предыдущего импульса ещё продолжается, когда происходит следующий импульс. Поэтому данный вид суммации называют также последовательной суммацией, которая играет важную физиологическую роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в нейронных объединениях нервных центров.
Пространственная суммация.
(рис. 4.7, б) Раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, тогда как при одновременной стимуляции обоих аксонов возникает ПД – процесс, который не может быть обеспечен одиночным ВПСП. Пространственная суммация связана с такой особенностью распространения возбуждения, как конвергенция.
Последействие – это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Последействие вызывает ряд причин: а) длительное существование ВПСП (полисинаптического или высокоамплитудного), когда при одном ВПСП возникает несколько ПД; б) многократные появления следовой деполяризации, что свойственно нейронам ЦНС (при Екр возникает ПД); в) циркуляция возбуждения по замкнутым нейронным цепям. Существование ВПСП и многократные появления следовой деполяризации действуют десятки-сотни миллисекунд, циркуляция возбуждения может продолжаться минуты и даже часы. Последействие играет важнейшую роль в процесса обучения (кратковременная память).
Словарь:
ПД – потенциал действия
ВПСП – возбуждающий постсинаптический потенциал
40. Физиология вегетативной нс (ВНС).
ВНС (автономная нс) – часть нс, регулирующая работу внутр. органов, просвет сосудов, обмен веществ и энергии.
Общие физиологический свойства (по сравнению с соматической):
- меньшая возбудимость,
- меньшая функциональная лабильность,
- меньшая скорость проведения возбуждения в вегетатив. нервах,
- пролонгированный во времени за счёт следовых фаз (до 150мс) потенциал действия,
- временная и пространственная суммация возбуждений в ганглиях.
- функции внс: поддержание постоянства внутр. среды, приспособление её к изменяющимся условиям окр. среды и деятельности организма.
- влияния внс на организм обычно не находятся под непосредств. контролем сознания.
- регуляция функций внутр. органов внс может осуществляться (< совершенно) при полном нарушении связи с цнс. Объясняется тем, что эффекторный нейрон внс находится за пределами цнс. Синапсы с ганглионар. нейронами образуют преганглионар. волокна, явл. аксонами нейронов, тела которых находятся в различ. отделах цнс, а также аксоны периферич. афферентных нейронов, образующих периферич. экстра- и интраорганные рефлекторные дуги. => при повреждении вегетативных центров сохраняются экстра- и интраорганные рефлексы, участвующие в регуляции функций внутр. органов. В эфферентной части сомат. нс все нейроны находятся в цнс, аксоны мотонейронов подходят непосредственно к эффекторным мышечным клеткам без перерыва. => повреждение передних корешков сомат. нервов спинного мозга или корешков черепных нервов ведёт к параличу двигат. активности соответствующих мышц.
- генерализованный (диффузный) хар-р распространения возбуждения в периферич. отделе внс объясняется, во-первых, феноменом мультипликации в вегетатив. ганглиях. Конвергенция и дивергенция в симпат. ганглиях обеспечивают высокую надёжность проведения возбуждения, т.к. ведут к пространственной и временной суммации постсинаптич. потенциалов.
Хим. организация. ВНС осуществляет свои регулирующие влияния главным образом путём хим. медиаторных и рецепторных мех-мов. На основании различий м/у хим. свойствами медиаторов и рецепторов внс м.б. разделена на:
1. Холинергическую часть внс. Холинергическими явл.:
а) все преганглионарные нейроны,
б) парасимпатические нейроны, иннервирующие потовые железы,
в) ганглионарные симпатические нейроны, иннервирующие сосуды скелетных мышц и осуществляющие при их раздражении вазодилатацию.
2. Норадренергическая часть внс. К таким относится больш-во ганглионарных симпатич. нейронов, выделяющих в своих окончаниях норадреналин. Особый мех-м – надпочечники, в кот. симпатич. нейроны утратили свои аксоны и секретируют норадреналин непосредственно в кровеносное русло.
Отдельные нейроны в симпатич. ганглиях секретируют дофамин. Часть симпатич. преганглионарных нейронов синтезируют гормон, освобождающий гонадотропин.
В ответ на стимуляцию симпатич. нервов в их окончаниях вместе с норадреналином выделяются: дофамин, β-гидроксилаза, метэнкефалин и др. биологически активные вещ-ва.
Тонус симпатич. нервов подвержен циркадианным колебаниям – днём выше, чем ночью.
В деят-сти нейронов ВНС в качестве модуляторов принимают участие различные олигопептиды идр. Биологически активные вещ-ва.
В симпатич. ганглиях осуществляют моделирующее действие вещ-во П, простагландин Е и серотонин. Активность ацетилхолина на отдельных нейронах тесно связана с влияниями вазоактивного интестинального пептида (ВИП), норадреналина, АТФ и нейропептида Y.
Отделы внс – симпатич. нс и парасимпатич. нс. Их принципиальное отличие в том, что у эффекторных нейронов и постганглионар. волокон различные медиаторы: у парасимпат. – ацетилхолин, у симпат. – норадреналин. => эффекты стимуляции симпат. и парасимпат. нервов, как правило, противоположны.
Рецепторы по мех-му действия дел. на 2 типа: ионотропные (когда ионный канал и место связывания медиатора не находятся на одной белковой молекуле), и метаботропные (когда через белок-рецептор активизируется цепочка внутриклеточных биохим. реакций).
41. Физиологические свойства симпатической НС (СНС)
Благодаря мех-мам мультипликации в симпатических ганглиях распространяющиеся из них возбуждения широко охватывают сразу несколько органов. В результате разветвлённых влияний на иннервируемые органы СНС оказывает на них и на организм в целом генерализованные влияния (наиболее отчётливо прослеживается при эмоц. реакциях).
При возбуждении СНС усиливаются сердеч. сокращения, увелич. их ритм, повышается артер. давление, улучшается оксигенация тканей, усил. метаболизм, нарастает содержание глюкозы в крови, увелич. скорость проведения возбуждения в скелетных мышцах, повыш. их тонус, расширяются бронхи, увелич. объём лёгочной вентиляции, расшир. зрачки, увелич. секреция катехоламинов надпочечниками. Одновременно сниж. тонус пищеварит. тракта, ослаб. процессы всасывания и ферментативного расщепления в кишечнике.
СНС участвует в формир-нии агрессии, стресса, болевых реакций, мобилизует организм на борьбу, бегство, активное взаимодействие с окр. миром.
Хим. особенности. В снс передача с пре- на ганглионарные нейроны осуществ. с помощью ацетилхолина, а на эффекторы – посредством норадреналина. Исключение: постганглионарные сипатич. волокна к потовым железам и отдельным сосудам, в окончанииях которых выделяется ацетилхолин.
СНС представлена на периферии на значит. протяжении постганглионарными волокнами типа С, => возбуждение по СНС распростран. медленнее, чем по парасимпатической.
Эффекты снс более продолжительны, чем в парасимпат. нс.
Электрические потенциалы в симпатич. ганглиях характеризуются продолжительными следовыми явлениями.
При стимуляции преганглионарных волокон в симпатич. ганглиях сначала регистрируется быстрый возбуждающий постсинаптический потенциал (ВПСП), кот. сменяется пролонгированным медленным ТПСП. Вслед за этим снова регистрируется медленный ВПСП (длится минуты).
Начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы. Медленный ТПСП определяется действием дофамина, кот. секретируется малыми по размеру интернейронами ганглия. Интернейроны, в свою очередь, активизируются через мускариновые рецепторы.
Локализация преганглионарных и ганглионарных нейронов и иннервируемые ими органы. В отличие от парасимпат. нервов, которые выходят из различ. органов ЦНС, все симпат. нервы выходят из спинного мозга, иннервируют все органы и ткани организма. В последнем шейном и двух первых грудных сегментах спинного мозга расположен спиноцилиарный центр, иннервирующий гладкие мышцы глаза, в т.ч. мышцу, расширяющую зрачок. Эффекторные нейроны этого пути лежат в краниальном шейном симпат. ганглии. От 2-4 грудных сементов спин. мозга отходят симпат. волокна, иннервирующие слюнные железы. В первых трёх сегметах спин. мозга расположены эффектор. симпат. нейроны, иннервир. сердце. Ганглионарные клетки этого пути лежат в основном в звёздчатом ганглии или (реже) в узлах пограничного симпат. ствола. Сосуды и потовые железы иннервируются ото всех симпат. преганглионарных нейронов спин. мозга.
Больш-во симпат. симпат. ганглиев удалено от иннервируемых ими органов, поэтому от этих ганглиев идут длинные постганглионар. аксоны. Исключение – некоторые небольшие ганглии, располож. рядом с половыми органами.
Симпат. нервы регулируют функции всех органов и тканей организма, включая ЦНС и сенсорные рецепторы.
Медиаторы и рецепторы.
1. преганглионар. нейроны и рецепторы ганглионар. нейронов. Эфферентный вход в вегетативном ганглии (экстра- и интрогенный) представлен возбуждающим холинергическим преганглионаным волокном, образующим синапс с ганглионар. нейроном при помощи Н-холинорецепотра (медиатор – ацетилцелин). Рецептор получил своё название (Д. Ленги) из-за чувствительности к никотину: малые его дозы возуждают нейрон, малые – блокируют. В вегетативных ганглиях кроме него имеются различные нейропептиды (М-холинорецепторы, вставочные адренергические клетки, регулирующие возбудимость нейронов).
2. От ганглионарных нейронов отходят постганглионар. симпат. волокна, в окончаниях которых главным медиатором явл. норадреналин – около 90%. Он синтезируется из аминокислоты тирозина в аксоне ганглионар. нейрона, в основном в терминал. симпат. окончаниях, около 1% - в теле нейрона. В ответ на раздражение симпат. нейрона вместе с норадреналином выдел. также белок хромагрин дофамин-β-гидроксилаза, мет-энкефалин. Эту смесь дополняют медиаторы соседних нервных окончаний, гормоны, метаболиты и ионы эффекторных клеток.
3. Инактивация медиатора. 75-80% выделившегося в синаптическую щель норадреналина захватывается обратно пресинаптической мембраной и поступает в пузырьки. Не попавший туда норадреналин разрушается моноаминоксидазой (МАК), локализованной на пресинаптич. мембране. Часть выделившегося в синаптич. щель и не связавшегося с пост- и пресинаптич. рецепторами норадреналина разрушается в области рецепторов постсинаптич. Мембраны ферментом катехол-О-метилтран… феразой (КОМТ) клеточных эффекторов, другая – диффундирует в интерстиций… кровь. Вклад МАО и КОМТ в процессы инактивации выделившихся в синаптич. щель катехоламинов невелик, а доля участия этих ферментов в разных органах и тканях различна: в ЦНС действует в основном МАО, в периферич. симпат. НС – КОМТ.
4. Эффекторные рецепторы. Выделившийся норадреналин действует на α- и β-постсинаптич. адренорецепторы, являющиеся …гликопротедами. Это деление рецепторов основано на чувствительности их к различным фармакологическим препаратам: α-адренорецепторы блокируются фентоламином, а β- - пропранолом. Оба типа рецепторов дел. на 2 подтипа: α1 и α2, β1 и β2-адренорецепторы. В больш-ве органов, реагирующих на катехоламины, содержатся оба рецептора.
Связь постгаглионар. симпат. окончаний с адренорецепторами. Считается, что симпат. иннервацию имеют только α1 и β1-адренорецепторы, которые активизируются в основном медиатором норадреналином. α2- β2- преимущественно не имеют иннервации, расположены вне синапсов на клетках-эффекторах и активизируются циркулирующим в крови адреналином и диффундирующим норадреналином поле его выделения пресинаптич. окончаниями. α2- β2- обнаружены также на пресинаптич. рецепторах, где они выполняют регулирующую функцию. Прямая связь посредством синапса имеется в сердце, жировой ткани, сосудах и в ряде гладкомышечных органов (мышцы зрачка, матки, кишечник и т.д.) непрямой тип регуляции осуществляется в больш-ве органов: скеленая мускулатура, эндокрин. железы и т.д. В них медиатор из сосудистых симпат. сплетений диффундирует к окружающим сосуд клеткам либо разносится кроью. Другой вариант: симпат. нервные волокна образуют синаптич. контакты с нейронами инраорганной НС.
Эффекты активации α- β-адренорецепторов. В физиолог. условиях реакция какого-либо органа на адреналин и норадреналин, поступ. с кровью либо выделяющиеся при возбуждении симпат. нервов, зависит от преобладания α- или β-адрнорцптров в данном органе и различной их возбудимости.
Активация α1- и α2- мембран клеток-эффекторов ведёт к деполяризации их и ↑ активности органа, в т.ч. сфинктеров желудочно-кишечного тракта. Однако у миоцитов желудка и кишечника развив. гиперполяризация, при этом мускулатура органов расслабляется.
Если же симпат. ствол раздражается в грудной полости, то в больш-ве случаев регистрир-ся не тормозные, а стимуляторные реакции желудка и 12перстной кишки. Это осуществ. с помощью серотонинергических нервных волокон, имеющихся в составе симпат. стволов.
Активация β-адрнорцптров вызывает разнонаправленные эффекты в различных органах: в сердце возникает гиперполяризация, угнетение функции органа, расширение сосудов. При одновременной активации α- и β- сосуды сужаются следствие преобладания α-.
Механизм действия катехоламинов на эффектоные органы.
Во всех видах адрнпцпторов катехоламины взаимодействуют посредством G-белка. При этом возникают электрофизиоог. и биохим. процессы. При деполяризации наблюд. Усиление функции органа, при гиперполяризации – угнетение, например угнетение сокращений кишки. При активации обоих типов рецепторов возник. метаболические сдвиги с помощью вторых посредников (биохим. действия медиатора): при активации α- - инозитол-3-фосфата и Ca2+, β- - аденилатциклазы-цАМФ.
Чувствительность эффекторных клеток к медиатору может понижаться или повышаться.
Билет 42