
Федеральная программа книгоиздания России Рецензенты: канд психол наук С. А. Исайчев, доктор биол наук И. И. Полетаева Равич-Щербо И. В. и др. Р12
| Вид материала | Программа |
- Вестник балтийской педагогической академии вып. 94. – 2010 г. Актуальные проблемы нравственного, 2431.92kb.
- Рецензенты: профессор, доктор психол наук Филонов Л. Б., вед науч сотрудник, канд психол, 2609.63kb.
- Научный выпуск вестник балтийской педагогической академии вып. 29. – 2000 г. Поиск, 1745.18kb.
- Общеобразовательная программа дошкольного образования Авторский коллектив, 5619.19kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Введенским Игорем Витальевичем Рецензенты доктор психол наук В. А. Лабунская канд психол, 375.9kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- Пояснительная записка, 12621.4kb.
- В организации совместных отношений, 1137.14kb.
Естественным отбором называется процесс дифференциального воспроизводства потомства генетически различными организмами в популяции. Фактически это означает, что носители определенных генетических вариантов (т.е. определенных генотипов) имеют больше шансов выжить и оставить потомство, чем носители других вариантов (генотипов). Дифференциальное воспроизводство может быть связано с действием разных факторов, среди которых называются смертность, плодовитость, оплодотворяемость, успешность спаривания и продолжительность репродуктивного периода, выживаемость потомства (иногда ее называют жизнеспособностью).
Мерой способности особи к выживанию и размножению является приспособленность. Однако, поскольку размер популяции обычно ограничен особенностями среды, в которой она существует, эволюционная результативность особи определяется не абсолютной, а относительной приспособленностью, т.е. ее способностью к выживанию и размножению по сравнению с носителями других генотипов в данной популяции. В природе приспособленность генотипов не постоянна, а подвержена изменению. Тем не менее в математических моделях значение приспособленности принимается за константу, что помогает при разработке теорий популяционной генетики. Например, в одной из наиболее простых моделей предполагается, что приспособленность организма полностью определяется структурой его генотипа. Кроме того, при оценке приспособленности допускается, что все локусы осуществляют независимые вклады, т.е. каждый локус может анализироваться независимо от других.
Выделяются три основных типа мутаций: вредоносные, нейтральные и благоприятствующие. Большинство новых мутаций, возникающих в популяции, являются вредоносными, так как снижают приспособленность их носителей. Отбор обычно действует против таких мутантов, и через некоторое время они исчезают из популяции. Данный тип отбора называется отрицательным (стабилизирующим). Однако существуют мутации, появление которых не нарушает функциониро-
117
вание организма. Приспособленность таких мутантов может быть так же высока, как и приспособленность аллелей-немутантов (исходных аллелей) в популяции. Эти мутации являются нейтральными, и естественный отбор остается равнодушным к ним, не действуя против них (дизруптивный отбор). При действии дизруптивного отбора внутри популяции обычно возникает полиморфизм — несколько отчетливо различающихся форм гена (см. гл. IV). Третий тип мутантов появляется крайне редко: такие мутации могут повысить приспособленность организма. В этом случае отбор может действовать так, что частота встречаемости аллелей-мутантов может повыситься. Данный тип отбора называется положительным (движущим) отбором.
ПОДСТАНОВКА ГЕНОВ
Предельным случаем эволюционирования популяции является полное исчезновение из нее исходных аллелей. Подстановкой генов (полной заменой одного аллеля на другой) называется процесс, в результате которого аллель-мутант вытесняет исходно доминировавший аллель «дикого типа». Иными словами, в результате действия различных популяционных процессов (например, мутационного процесса, случайного дрейфа генов, отбора) в популяции обнаруживаются только аллели-мутанты: аллель-мутант появляется в популяции в единственном числе в результате единичной мутации, а затем, после смены достаточного количества поколений, его частота достигает 100%, т.е. он фиксируется в популяции. Время, требующееся аллелю для достижения 100% частоты встречаемости, называется временем фиксации. Очевидно, что не все аллели-мутанты достигают 100% встречаемости и фиксируются в популяции. Обычно бывает наоборот; большинство аллелей-мутантов в течение нескольких поколений элиминируется. Вероятность того, что данный аллель-мутант зафиксируется в популяции, обозначается величиной, называемой вероятностью фиксации.
Новые мутанты возникают в популяциях постоянно, при этом в качестве одного из сопровождающих мутацию процессов разворачивается процесс подстановки генов, в которых аллель А заменяется новым аллелем Б, а тот в свою очередь заменяется аллелем В и т.д. Динамика этого процесса описывается понятием «скорость процессов подстановки генов», отражающим количество подстановок и фиксаций в единицу времени.
3. ФАКТОРЫ, ВЛИЯЮЩИЕ НА ДИНАМИКУ
ИЗМЕНЕНИЯ ЧАСТОТ ГЕНОТИПОВ В ПОПУЛЯЦИИ
Мутации, миграция, отбор и дрейф генов влияют на динамику частот как конкретных аллелей, так и целостных генотипов. Определенные типы скрещивания влияют только на частоты генотипов. Среди этих процессов - инбридинг и ассортативное скрещивание.
118
ИНБРИДИНГ
Закон Харди-Вайнберга действует лишь тогда, когда скрещивание случайно и вероятность скрещивания двух генотипов равна произведению их частот. Инбридинг же представляет собой один из вариантов неслучайного скрещивания, когда потомство производится особями, являющимися генетическими родственниками друг другу. Поскольку родственные особи в генетическом отношении более сходны между собой, чем не состоящие в родстве организмы, постольку инбридинг ведет к повышению частоты гомозигот и снижению частоты гетеро-зигот по сравнению с теоретически ожидаемой при случайном скрещивании (хотя частоты аллелей при этом не меняются). Крайним случаем инбридинга является самооплодотворение или самоопыление — формы размножения, которые широко распространены у некоторых видов растений. Общая закономерность заключается в том, что в популяциях инбридинг повышает частоту фенотипического проявления вредных рецессивных аллелей, поскольку повышает вероятность «встречи» двух рецессивных генов, определяющих то или иное отклонение от нормы.
Мерой генетических последствий инбридинга служит коэффициент инбридинга, представляющий собой вероятность того, что у какой-либо особи в данном локусе окажутся два аллеля, идентичные по происхождению, т.е. точные копии аллеля, имевшегося в генотипе одного из прародителей этой особи в каком-то из предшествующих поколений. Коэффициент инбридинга обычно обозначается буквой F (табл. 5.1).
Таблица 5.1
Коэффициент инбридинга (F) в потомстве от родственных скрещиваний
| Тип скрещивания | Степень родства | F |
| Сиблинги | I | 1/4 |
| Дядя-племянница, тетя-племянник Двоюродные брат— сестра | II | 1/8 |
| Двоюродные дядя-племянница или двоюродные тетя-племянник Троюродные брат— сестра Троюродные дядя— племянница или троюродные тетя-племянник | III | 1/32 1/64 1/128 |
| Четвероюродные брат— сестра | IV | 1/256 |
119
Ряс. 5.1. Результаты выполнения теста Векслера детьми от кровосмсшан-ных браков [по 363].
«—• — результаты выполнения тестон детьми от кровосмешанпых (двоюродных
братьев и сестер) браков; —- — результаты выполнения тестов детьми из контрольной выборки.
У растений при самоопылении коэффициент самый большой (F= 1/2). У человека супружеские отношения между родителями и детьми или между братьями и сестрами считаются кровосмешением; в большинстве культур (но не во всех) существует запрет на подобные браки. Скажем, в династиях египетских фараонов крово-смешанные браки встречались часто. В Японии браки между двоюродными сестрами и братьями поощряются и в некоторых областях составляют до 10% общего числа браков. В Индии существуют касты, одобряющие браки между дядей и племянницей. А вот в римской католической церкви, напротив, требуется получение специального разрешения на заключение брака между любыми родственниками, которые кровно ближе друг другу, чем родственники третьей степени.
В литературе можно найти множество примеров того, что инбридинг повышает вероятность проявления рецессивных заболеваний. Кроме того, как показали исследования, инбридинг приводит к снижению среднепопуляционных значений по ряду поведенческих и психологических признаков. Например, согласно некоторым исследованиям, в результате инбридинга среди потомков близкородственных браков наблюдается некоторое снижение IQ (рис. 5.1).
120
АССОРТАТИВНОСТЬ
(НЕСЛУЧАЙНЫЙ ПОДБОР СУПРУЖЕСКИХ ПАР)
Вопрос о том, что движет людьми при заключении брачных союзов, остается загадкой до сих пор. Этот вопрос привлекает внимание как философов и поэтов, так и специалистов по планированию семьи. В рамках психогенетики также существует раздел, посвященный этой теме. Изучение закономерностей ассортативного подбора супружеских пар (неслучайного заключения браков) — важная линия психогенетических исследований, поскольку ассортативность может изменять оценки наследуемости признака, влиять на его вариативность в популяции и т.д. Один из основных результатов этих исследований указывает на то, что ассортативность практически всегда осуществляется в положительном направлении, т.е. браки в подавляющем большинстве случаев заключаются между людьми, похожими друг на друга; в супружеских парах, как правило, существует сходство по многим фено-типическим признакам.
Подобно инбридингу, ассортативность браков влияет только на частоты генотипов, но не на частоты аллелей. Если мы задумаемся о влиянии отдельно взятого локуса на признак, по которому происходит ассортативный подбор, то окажется, что ассортативность, как и инбридинг, снижает гетерозиготность. Иными словами, в результате ассортативности гомозиготы заключают браки с гомозиготами, а ге-терозиготы в каждом поколении производят одну или несколько гомозигот. В конечном счете, если ассортативность выражена достаточно сильно, она может существенным образом снизить генетическую изменчивость в популяции. Например, различия по росту и весу представляют собой в основном результат влияния множества генов. Если бы признак роста не был существенным фактором при выборе спутников жизни и высокие женщины выходили бы замуж за невысоких мужчин так же часто, как и за высоких, то потомки высоких женщин были бы среднего роста. Однако хорошо известно, что супруги подбираются по росту неслучайно, дети высоких женщин чаще всего имеют высоких отцов, что, в свою очередь, делает высокими и их. В этом смысле ассортативность увеличивает разброс по признаку в популяции, поскольку дети от ассортативных браков имеют фенотипическое значение признака, отстоящее от среднепопуляционного дальше, чем если бы браки заключались случайно.
Ассортативность браков представляет собой одну из самых интересных проблем психогенетики. Ее механизм еще не понят, однако хорошо известен тот факт, что наличие ассортативности изменяет популяцион-ное распределение значений по фенотипу, по которому наблюдается ассортативность. Поэтому, проводя исследование любого признака, по которому наблюдается ассортативность, исследователи стремятся учитывать в статистическом анализе корреляции между супругами (несколько подробнее феномен ассортативности рассматривается в гл. VI).
121
* * *
Итак, мы рассмотрели закономерности существования генов в популяциях. Частоты генов в популяциях, находящихся в состоянии покоя, определяются законом Харди-Вайнберга. Однако реальные популяции никогда не бывают в состоянии покоя, и закон Харди-Вайнберга постоянно нарушается. В реальных популяциях в каждый данный момент времени одновременно присутствуют и взаимодействуют все основные факторы популяционной динамики. Необходимо различать популяционные факторы, влияющие на изменения частот аллелей и частот генотипов. На изменение частот аллелей в популяции более других процессов влияют мутации, миграции, естественный отбор и генетический дрейф. Частота встречаемости генотипов в популяции подвержена влиянию определенных типов скрещивания, в том числе инбридинга и ассортативного подбора супружеских пар.
Глава VI
ГЕНОТИП В КОНТЕКСТЕ: ТИПЫ СРЕДОВЫХ ВЛИЯНИЙ И ГЕНОТИП-СРЕДОВЫХ ЭФФЕКТОВ
1. ОСНОВНЫЕ ПОНЯТИЯ
Индивидуум (носитель генотипа) существует не в вакууме, а среди подобных ему индивидуумов. Кроме того, сам генотип не спрятан, как Кащеева смерть, за семью замками на конце иголки, а чрезвычайно открыт и крайне чувствителен к любому оказываемому на него влиянию. Изучение генотипа невозможно и неадекватно без изучения среды, в которой он находится. Генетические исследования заметно выигрывают, когда в них используются хорошо отлаженные психологические методики, оценивающие характеристики среды. Подобным же образом психологические и педагогические исследования средо-вых условий, влияющих на формирование тех или иных поведенческих признаков, выигрывают от использования накопленных психогенетикой знаний о том, как функционирует генотип, что такое норма реакции и каковы пределы пластичности генотипа. Время противопоставления «двух факторов» — генов и среды — осталось позади. Сегодня мы знаем достаточно для того, чтобы без тени сомнений утверждать: формирующаяся индивидуальность не делится на то, что в ней от среды, и на то, что — от генотипа. Развитие по сути своей является процессом переплетения и взаимодействия генов и среды, развитие и есть их взаимодействие.
В контексте психогенетического изучения средовых особенностей чрезвычайно важны три момента.
122
* * *
Итак, мы рассмотрели закономерности существования генов в популяциях. Частоты генов в популяциях, находящихся в состоянии покоя, определяются законом Харди-Вайнберга. Однако реальные популяции никогда не бывают в состоянии покоя, и закон Харди-Вайнберга постоянно нарушается. В реальных популяциях в каждый данный момент времени одновременно присутствуют и взаимодействуют все основные факторы популяционной динамики. Необходимо различать популяционные факторы, влияющие на изменения частот аллелей и частот генотипов. На изменение частот аллелей в популяции более других процессов влияют мутации, миграции, естественный отбор и генетический дрейф. Частота встречаемости генотипов в популяции подвержена влиянию определенных типов скрещивания, в том числе инбридинга и ассортативного подбора супружеских пар.
Глава VI
ГЕНОТИП В КОНТЕКСТЕ: ТИПЫ СРЕДОВЫХ ВЛИЯНИЙ И ГЕНОТИП-СРЕДОВЫХ ЭФФЕКТОВ
1. ОСНОВНЫЕ ПОНЯТИЯ
Индивидуум (носитель генотипа) существует не в вакууме, а среди подобных ему индивидуумов. Кроме того, сам генотип не спрятан, как Кащеева смерть, за семью замками на конце иголки, а чрезвычайно открыт и крайне чувствителен к любому оказываемому на него влиянию. Изучение генотипа невозможно и неадекватно без изучения среды, в которой он находится. Генетические исследования заметно выигрывают, когда в них используются хорошо отлаженные психологические методики, оценивающие характеристики среды. Подобным же образом психологические и педагогические исследования средо-вых условий, влияющих на формирование тех или иных поведенческих признаков, выигрывают от использования накопленных психогенетикой знаний о том, как функционирует генотип, что такое норма реакции и каковы пределы пластичности генотипа. Время противопоставления «двух факторов» — генов и среды — осталось позади. Сегодня мы знаем достаточно для того, чтобы без тени сомнений утверждать: формирующаяся индивидуальность не делится на то, что в ней от среды, и на то, что — от генотипа. Развитие по сути своей является процессом переплетения и взаимодействия генов и среды, развитие и есть их взаимодействие.
В контексте психогенетического изучения средовых особенностей чрезвычайно важны три момента.
122
Во-первых, генетические исследования постоянно указывают на критическую роль средовых факторов в формировании психологических различий между людьми. Многочисленные психогенетические работы нагляднейшим образом показали, сколь важна роль генетических факторов для объяснения межиндивидуальной вариативности по самым разным признакам. В некоторых случаях (например, для вариативности* показателей интеллекта) генетические влияния объясняют 50% фенотипической изменчивости, Чем, однако, объясняются остальные 50%? Ответ на этот вопрос очень прост: в большинстве своем за оставшиеся 50% отвечает среда, точнее особенности среды, в которой развиваются и живут носители генотипов.
Во-вторых, в контексте генетики количественных признаков понятие среды определяется намного шире, чем в психологии. Согласно этому определению, понятие «среда» включает все типы средовых влияний — общесемейные, индивидуальные и любые другие (включая ее физические и физиологические компоненты, пренатальные условия, диету, болезни раннего детства и т.д.), в то время как в психологии средовые условия обычно приравниваются только к социально-экономическим и психологическим условиям взросления ребенка.
В-третьих, психогенетика концентрирует свои усилия на вопросе о том, что есть (в данный момент в данной популяции), а не на вопросе о том, что может произойти. Например, высокие значения коэффициента наследуемости, полученного при изучении межиндивидуальной вариативности по росту, констатирует тот факт, что в данное время в данной популяции дисперсия роста объясняется в основном генетическими различиями между членами данной популяции (что есть). Однако определенное средовое вмешательство (например, изменение питания и увеличение количества витаминов в рационе) может оказать влияние на формирование межиндивидуальных различий в популяции по такому высоконаследуемому признаку, как рост (что может произойти). Считается, что именно такие средовые изменения привели к увеличению среднего роста в современных европейских популяциях по сравнению с аналогичными оценками, полученными, например, в начале XX в. Индивидуальные же различия по росту внутри каждого поколения находятся под жестким генетическим контролем.
При изучении генотипа в его средовом контексте исследователи выделяют два класса влияний - влияния среды и генотип-средовые влияния, т.е. совместные, сопряженные влияния среды и генотипа.
* В данном учебнике понятия «изменчивость» (подробнее о ней — в гл. I) и «вариативность» используются как синонимы. Тем не менее авторы отдают предпочтение словосочетаниям «фенотипическая изменчивость», «генотипическая изменчивость», но «средовая вариативность». Напомним, что статистическим эквивалентом понятия изменчивости является понятие дисперсии.
123
Для психолога одним из наиболее интересных аспектов психогенетики является ее способность дать надежную информацию о наличии и степени средовых влияний на формирование исследуемого признака. Психогенетические исследования могут не только определить, влияет ли среда на развитие и проявление этого признака (и если да, то в какой степени), но и указать конкретный класс средовых влияний, в котором надо искать определенный содержательный фактор, являющийся критической средовой переменной для анализируемого признака. Например, психогенетическое исследование может определить, что популяционная изменчивость по интеллекту зависит от общесемейной среды, и тем самым направить исследовательскую работу психологов на поиск конкретных компонентов именно этой среды (например, социально-экономических характеристик семьи), значимых для формирования интеллекта. В рамках теоретической психогенетической модели исследователи выделяют два основных типа средовых влияний: общие и индивидуальные.
Термином общая среда (синонимы - общесемейная, межсемейная, систематическая, семейная, разделенная; чаще всего обозначается символом Е— shared или Ес—common, общая) описываются типы средовых влияний, одинаковых для членов изучаемых пар родственников (родители—дети, сиблинг—сиблинг и т.д.). Выделяют два класса общих средовых влияний: (1) общесемейная среда, характеристики которой различны в разных семьях, но едины для всех членов одной семьи (социоэкономический статус семьи, уровень образования родителей, жилищные условия, исповедуемая семьей религия и т.п.); и (2) семейная среда, общая для конкретных пар членов данной семьи (близнецовая среда, среда сиблингов, родительско-детская среда и т.д.); это — среда, общая только для членов данной диады (например, школьная среда близнецов, обучающихся в одном классе, будет общей для близнецов, но не для родительско-детских пар).
Термином «индивидуальная среда» (синонимы — внутрисемейная, уникальная, специфическая, различающаяся, неразделенная, случайная; чаще всего обозначается EN — nonshared, т.е. среда, не являющаяся единообразной для всех членов семьи) описываются типы среды, различные у разных членов одной семьи. Индивидуальные средовые характеристики — это набор тех средовых условий, влияния которых делают членов одной семьи непохожими друг на друга. Например, как ни стараются родители сиблингов относиться к ним одинаково объективно, этого никогда не случается. Индивидуальная среда включает широкий спектр разных характеристик, начиная от разнообразия пре-натальных условий и кончая социально-психологическими особенностями среды, специфическими для данного индивида. Примерами таких средовых влияний могут служить характеристики круга друзей и рабочей обстановки, которые, как правило, не единообразны, специфичны для каждого члена семьи.
124
Некоторые исследователи предлагают более детальную классификацию сред и разделяют общие и индивидуальные средовые влияния на (1) систематические (повторяющиеся, долгосрочные; примерами систематических общесемейных влияний могут служить заработная плата родителей, а индивидуальных систематических влияний — школьная среда ребенка или постоянное общение с близким другом) и (2) случайные (краткосрочные; пример случайных общесемейных средовых воздействий — смерть члена семьи, а случайных индивидуально-средовых влияний - болезнь или встреча с интересным человеком)*.
Эти теоретически выделенные типы средовых влияний представлены в центральной модели генетики количественных признаков в терминах компонентов фенотипической дисперсии. Далее мы покажем, как сопоставление разного типа родственных корреляций позволяет выделять разные компоненты среды. Однако некоторые средовые влияния (например, систематические и несистематические эффекты) в рамках центральной модели разделить нельзя. Для их изучения традиционную (классическую) модель разложения фенотипической дисперсии, в состав которой входят только латентные переменные, необходимо модифицировать с тем, чтобы включать в нее измеряемые переменные (подробнее об этом в гл. VIII).
Ученые давно осознали тот факт, что псевдодихотомическое разделение факторов, значимых для формирования индивидуальности, на те, которые «от генов», и те, которые «от среды», является искусственным и упрощенным. Помимо двух главных составляющих фенотипической дисперсии — генетической и средовой — существует несколько факторов, возникающих в результате взаимодействия (или совместных действий) среды и генов, и часто этим факторам принадлежит весьма существенная роль. При описании гено-средовых эффектов выделяются три разных класса явлений: генотип-средовые корреляции (ГС-корреляции), или ковариации; генотип-средовые взаимодействия (ГС-взаимодействия); ассортативность.
Средовые влияния, переживаемые человеком, часто не случайны, а, напротив, коррелируют с особенностями его генотипа или даже вызываются ими. Это явление было многократно описано. Смысл генотип-средовой корреляции хорошо передает утверждение «каждому по способностям». Теоретически предполагается, что если среда достаточно разнообразна и общество предоставляет человеку возможность выбора, каждый выбирает и получает среду, коррелирующую (положительно или отрицательно) с его генетически обусловленной индивидуальностью. (Например, человек с генетически заданным
* Надо иметь в виду, что термином «случайный» обозначается не причина возникновения данного события, а распределение данного обстоятельства в популяции по закону случайных чисел.
125
медленным индивидуальным темпом едва ли выберет профессию, в которой деятельность требует высокой скорости.)
Позитивная ГС-корреляция характеризует ситуацию двойного преимущества для носителя определенного генотипа. Так, дети композитора Иоганна Себастьяна Баха, будучи наследственно музыкально одаренными, выросли в обогащенной музыкальной среде, поскольку отец посвящал много времени их музыкальным занятиям. Семья математиков Бернулли примечательна не только наследственной математической одаренностью, но и, как следствие ее, «математизированной» средой (см. гл. VII).
Негативная ГС-корреляция отражает ситуацию регрессии к попу-ляционному среднему, т.е. тенденцию к снижению фенотипического значения признака, например, у потомков по сравнению с родителями из-за реакции среды на их генетическую индивидуальность (подробнее о ГС-корреляциях см. гл. VIII).
В целом ГС-корреляция отражает феномен неслучайного распределения сред между различными генотипами. «Хороший генотип» обычно получает и «хорошую среду»; «плохой» может выбрать адекватную себе, но объективно «плохую» среду. Понятно, что этот феномен может искажать оценку наследуемости: в случае положительной корреляции, когда и генотип, и среда действуют в одном направлении, она может быть завышена, в случае отрицательной, наоборот, занижена. Это — существенная помеха в исследованиях, которая должна, по возможности, контролироваться,
ГС-взаимодействие следует
четко отличать от ГС-корреляции. Подобное взаимодействие ничего общего с идеей распределения «каждому по способностям» не имеет. Оно отражает непосредственный акт совместного действия, совместного
Обедней- Нормаль- Обогащен- влияния генов и средовых фак-ная пая пая торов на фенотип. ГС-взаимо-
Типы среды действие описывает генетичес-
кий контроль чувствительности к средовым различиям и обна-
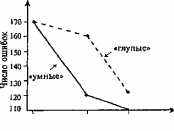
Рис. 6.1. Взаимодействие генотипа и руживается в том, что один и среды (по Cooper a. Zubek, 1958). тот же генотип в р
Среднее число ошибок при решении ла- , ж
г - даст разные фенотипы (а раз-
биринтной задачи «умными» и „ „
«глупыми» крысами, выращенными в ные генотипы в одной и той
обедненной, нормальной и обогащенной же среде тоже получат разное
средах. фенотипическое выражение)
(рис. 6.1).
126
В этих экспериментах при помощи направленного отбора были выведены линии «умных» (быстро обучающихся) и «глупых» (обучающихся медленно) крыс. В стандартной («нормальной») среде «глупые» делали в среднем на 50 ошибок больше, чем «умные». Однако в обедненной среде эта разница исчезала: количество ошибок у обеих линий становилось одинаковым, причем «умные» резко «глупели»; в обогащенной же среде, наоборот, «умнели» «глупые» —у них количество ошибок резко сокращалось. Обратим внимание на то, что реакция на плохую среду выше у «умных»; создание же более благоприятных условий сильнее сказывалось на обучаемости «глупых» крыс. (Вспомним понятие «норма реакции», — гл. I.)
Понятно, что у человека экспериментально исследовать ГС-взаимодействие невозможно. Но примером может служить генетическая сопротивляемость какому-либо заболеванию. Люди, генетически склонные к заболеванию, не проявляют симптомов заболевания только до тех пор, пока их среда обитания свободна от патогенов. И наоборот, генетически устойчивые к заболеванию индивидуумы не проявляют симптомов заболевания даже при наличии патогена в их среде. Иначе говоря, изменение среды внесением в нее патогена оказывает весьма разные (порой, трагически разные) влияния на фенотипы индивидуумов, генетически склонных и генетически устойчивых к заболеванию.
Ассортативностью называется неслучайное заключение браков на основе сходства по любым факторам. О ней уже шла речь в гл. V как об одном из факторов, меняющих частоты генотипов в популяциях. Исследователи, применяющие метод семьи (см. далее и гл. VII) для анализа психологических признаков, часто ссылаются на этот феномен. Мерой ассортативности принято считать корреляцию между супругами.
В большинстве современных обществ браки заключаются на основе взаимного интереса и привязанности будущих супругов друг к другу, что подразумевает сходство супругов по ряду фенотипических признаков, например по интеллекту, интересам, увлечениям и т.д. Высокая положительная ассорта -тивность была обнаружена у таких признаков, как уровень образования, религиозные и политические установки, социально-экономическое положение. Умеренная ассортативность установлена для физических показателей и когнитивных характеристик, например когнитивных стилевых особенностей и специальных способностей.
Исследования также продемонстрировали, что личностные качества и особенности темперамента не являются значимым критерием при подборе спутника жизни: корреляции между этими признаками у супругов достаточно низки. Однако до сих пор нет исследований, в которых были бы выявлены отрицательные корреляции между супругами по какому-либо (физическому или психологическому) признаку.
Ассортативность может искажать оценки влияния как генетических, так и средовых составляющих фенотипической дисперсии и потому должна учитываться во многих генетико-математических моде-
127
лях, точнее, в любых моделях, где она может занижать или завышать оценки искомых параметров. Важно помнить, что ассортативность не может интерпретироваться как побочная переменная, поскольку, строго говоря, переменной не является. В психогенетических моделях ассортативность представлена в качестве определенного коэффициента [44], искажающего истинную фенотипическую корреляцию в парах родитель-ребенок. Статистически контролировать ассортативность можно только в условиях использования комбинированных психогенетических моделей, т.е. в квазиэкспериментальных исследованиях, объединяющих несколько методов психогенетики (например, близнецовый и семейный).
Например, тенденция заключать браки с теми, кто близок нам по уровню интеллекта, давно была подмечена как в экспериментальных работах, так и в житейских наблюдениях. В среднем корреляция супругов по IQ составляет примерно 0,30-0,40. Такое сходство вовсе не означает, что мы подбираем жизненных партнеров по результатам выполнения интеллектуальных тестов. Эта корреляция определяется и интеллектуальной близостью будущих супругов, и тем, что общение с интеллектуально равным партнером психологически комфортно, и массой других обстоятельств. Однако, независимо от механизма ассортативности, в результате заключения браков между людьми, похожими по уровню интеллектуальных способностей, в зачатии нового организма принимают участие генотипы, сходство которых выше того, которое определяется законом случайных чисел. Данная закономерность особенно важна на концах распределения оценок познавательных способностей, поскольку было отмечено, что ассортативность проявляется ярче как среди одаренных, так и среди интеллектуально отсталых людей. В группе с низкими образованием и интеллектом Т.Д. Думитрашку получила очень высокую ассортативность по интеллекту (r = 0,68) и никаких корреляций по темпераменту, когнитивному стилю, креативности [53; 54]. В подобных случаях ассортативность увеличивает вероятность того, что произойдет «удвоение» наследуемой генной информации и, соответственно, потомок таких родителей унаследует двойную дозу генов, предрасполагающих к фенотипу, носителями которого являются его родители.
В последние 5-10 лет психогенетики обнаружили три весьма неожиданных явления: (а) у детей, растущих в одной семье, среда формирует скорее различия, чем сходства; (б) многие психологические инструменты (опросники, данные, полученные методом наблюдения, и пр.), используемые для измерения характеристик среды, показывают неожиданно высокий уровень генетического контроля, и (в) при разложении фенотипической дисперсии подавляющего большинства психологических признаков, изучаемых психогенетикой, роль общесемейной среды оказывается незначительной. Все это дает возможность сформулировать гипотезу о том, что люди создают или находят определенные средовые условия, соответствующие их генотипам, а не являются пассивными «жертвами» своих генов или «доставшейся» им среды. Иными словами, индивидуальный генотип оказывается «конструктором» индивидуальной среды. Эти данные позволили аме-
128
риканской исследовательнице С. Скарр сформулировать концепцию, получившую образное обозначение «генотип —> среда», т.е. путь от генотипа к среде, обратный привычному «влиянию среды на индивидуальность» [396]. О продуктивности такого подхода речь пойдет далее. В соответствии с этими тремя наблюдениями и построена данная глава. Во-первых, мы рассмотрим основные результаты изучения общей семейной среды; во-вторых, уделим особое внимание анализу находок, полученных в исследованиях индивидуальной среды; в-третьих, проанализируем феномен ГС-корреляции, описывая те особенности среды индивидуальности, которые коррелируют с ее генотипом; и, наконец, более детально познакомим с понятием ГС-взаимодействия, или феноменом так называемой чувствительности к среде.