
Н. Н. Алипова, канд биол наук О. В. Левашова и канд биол наук М. С. Морозовой под редакцией акад. П. Г. Костюка москва «мир» 1996 ббк 28. 903 Ф50
| Вид материала | Документы |
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- М. А. Ляшко доц., канд физ мат наук; Т. Н. Смотрова доц., канд, 2299.13kb.
- Современные направления развития физической культуры, спорта и туризма, 4493.6kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Пособие для врачей и среднего медицинского персонала Минск, 5480.63kb.
- Ббк 63. 3(0) Н72, 4378.93kb.
- Тезисы докладов, 4290.75kb.
- Строительные нормы и правила нагрузки и воздействия сниП 01. 07-85* министерство строительства, 1162.86kb.
- Ббк 63. 3(0) Н72, 5546.58kb.
17.2. Система задней доли гипофиза
Функциональная организация
Связь между гипоталамусом и гипофизом. Согласно классическому определению эндокринной системы следует считать, что некоторые участки центральной нервной системы функционируют как эндокринные железы: они вырабатывают гормоны и выделяют их в кровь для доставки к органумишени. Одной из таких структур является гипофиз. Он расположен внутри черепа в костной ямке турецкого седла, хорошо защищающей его от повреждения. Гипофиз - это сложный орган, состоящий из трех совершенно разных частей (рис. 17.7). Задняя доля гипофиза, или нейрогипофиз, образована окончаниями аксонов нервных клеток, тела которых находятся в гипоталамусе. Передняя доля гипофиза, или аденогипофиз, представляет собой не нервную структуру, а скопление клеток, секретирующих гормоны. Передняя и задняя доли гипофиза разделены тонким слоем клеток, обра-
1
 ) В последние годы многие исследователи приходят к выводу, что рецепторы стероидных гормонов локализованы в ядре, а в цитоплазму они попадают при субклеточном фракционировании (Jasc J. Μ. Baulieu Ε. Ε. J. Steroid Biochem., 24, 177, 1987).- Прим. перев.
) В последние годы многие исследователи приходят к выводу, что рецепторы стероидных гормонов локализованы в ядре, а в цитоплазму они попадают при субклеточном фракционировании (Jasc J. Μ. Baulieu Ε. Ε. J. Steroid Biochem., 24, 177, 1987).- Прим. перев. 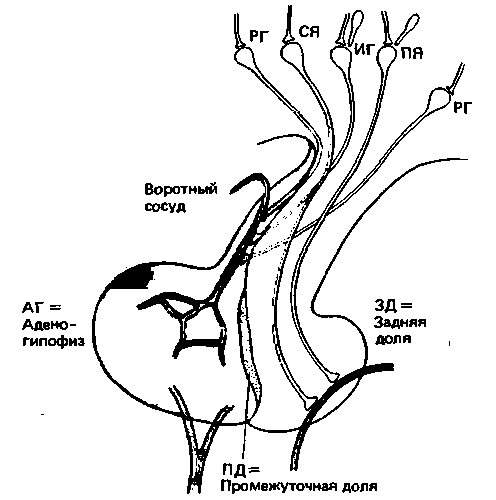 |
| Рис. 17.7. Расположение передней (АГ), промежуточной (ПГ) и задней (ЗД) долей гипофиза по отношению к гипоталамусу и воротной системе. Задняя доля гипофиза состоит из терминален аксонов крупных клеток супраоптического и паравентрикулярного ядер (СЯ и ПЯ соответственно) гипоталамуса. Промежуточная доля состоит всего из одного-двух слоев клеток, и у человека, вероятно, не выполняет никакой функции. Передняя доля имеет эпителиальное строение. Активность ее клеток регулируется рилизинг- и ингибирующими нейрогормонами (РГ и ИГ соответственно), секретируемыми клетками гипоталамуса. Гипоталамические нейроны, секретирующие зти нейрогормоны, иннервируются лимбической системой и средним мозгом, а также нейронами самого гипоталамуса. Таким путем клетки гипоталамуса получают информацию о внутренних и внешних воздействиях |
зующих промежуточную долю, которая иннервируется нервами, идущими из гипоталамуса. Промежуточная доля имеет большое значение у низших позвоночных и значительно меньшее у млекопитающих. Поскольку неизвестны какие-либо патологические проявления, связанные с промежуточной долей гипофиза, секретируемые ею гормоны не будут рассматриваться в настоящем разделе (см., однако, с. 401).
Гормоны задней доли гипофиза. В гипоталамусе имеются две группы очень крупных клеток, образующих супраоптическое и паравентрикулярное ядра. Аксоны образующих эти ядра нейронов проходят по ножке гипофиза в турецкое седло и образуют здесь заднюю долю гипофиза (рис. 17.7). В расширенных терминалях этих аксонов хранятся
IM ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
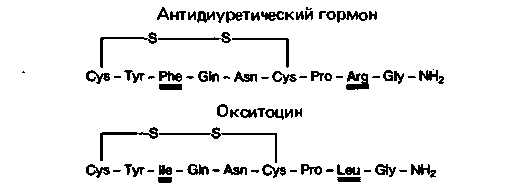 |
| Рис. 17.8. Аминокислотные последовательности антидиуретического гормона (АДГ), называемого также вазопрессином, и окситоцина |
два гормона - окситоцин и антидиуретический гормон (АДГ), или вазопрессин. Название «антидиуретический гормон» предпочтительнее: оно лучше отражает функцию гормона, поскольку у человека он обладает вазопрессорной активностью при очень высоких концентрациях, которые обнаруживаются только в патологических условиях (с. 540).
Из задней доли гипофиза АДГ и окситоцин высвобождаются в общую систему кровообращения. Молекулы обоих гормонов состоят из 9 аминокислотных остатков, два из которых - остатки цистеина. Последние связаны между собой дисульфидным мостиком, в результате чего образуется кольцо из пяти аминокислотных остатков (рис. 17.8).
Только при наличии такого кольца гормоны обладают биологической активностью. Поскольку две молекулы цистеина, соединенные дисульфидной связью, образуют одну молекулу цистина, окситоцин и АДГ обычно называют октапептидами. Установлено, что оба пептида синтезируются не в виде октапептидов. а в виде значительно более крупных молекул-предшественников. С помощью современных методов генной инженерии удалось определить аминокислотную последовательность этих предшественников [4. 14]. Высокомолекулярные предшественники окситоцина и АДГ синтезируются в аппарате Гольджи нейронов крупноклеточных ядер гипоталамуса. Раньше считали, что окситоцин синтезируется только в паравентрикулярном ядре, а АДГ-в супраоптическом, но теперь установлено, что и в том, и в другом ядрах синтезируются оба гормона. Однако в каждой клетке может вырабатываться только один из этих гормонов.
Аксонный транспорт и секреция гормонов. Молекулы предшественников путем аксонного транспорта поступают в заднюю долю гипофиза. В теле нейрона, аксоне и его терминалях АДГ и окситоцин, находящиеся в транспортных гранулах, выщепляются из молекулы-предшественника. Недавно из гранул были выделены крупные пептиды, названные нейрофизинами, и определен их состав. Вначале думали, что нейрофизины синтезируются отдельно от окситоцина и АДГ и служат белками - перенос-
чиками этих гормонов. Однако теперь установлено, что нейрофизины являются составной частью больших молекул-предшественников, от которых отщепляются окситоцин и АДГ. Помимо двух нейрофизинов (один образуется из предшественника окситоцина, а другой-из предшественника АДГ) гранулы содержат другие пептидные фрагменты, роль которых неизвестна.
Окситоцин и АДГ хранятся в гранулах терминальных структур аксонов нейрогипофиза. Потенциал действия, возникающий в клетках супраоптического ядра или паравентрикулярного ядра, передается по аксону в концевую структуру. Деполяризация терминали через механизм сопряжения электрического потенциала с секрецией приводит к высвобождению гормона путем экзоцитоза из нейросекреторных гранул в кровеносную систему.
Антидиуретический гормон
Действие АДГ. В физиологических условиях органом-мишенью АДГ служат почки. Действие этого гормона на осморегуляцию впервые было обнаружено в пионерских опытах Вернея [42], который вводил в сонную артерию кошек гиперили гипотонический раствор NaCl и записывал потенциалы действия супраоптического ядра. Гипертонический раствор повышал активность нейронов супраоптического ядра, а гипотонический вызывал ее отчетливое снижение. Иными словами, гиперосмотический стимул вызывал повышение электрической активности нейронов, секретирующих АДГ, в результате чего больше АДГ выделялось в кровь и доставлялось к органу-мишени -собирательным трубочкам и выводным протокам почек. В отсутствие АДГ эпителий собирательных трубочек почек непроницаем для воды, а под действием АДГ становится проницаемым, что обеспечивает пассивную реабсорбцию воды (с. 807). Таким образом, в условиях гиперосмолярности АДГ обеспечивает образование гиперосмолярной мочи и минимальную, по возможности, потерю воды. В итоге осмотическая нагрузка на организм уменьшается. Соответственно в гипоосмолярных условиях секреция АДГ уменьшается. Употребление алкоголя еще сильнее снижает секрецию АДГ, чем объясняется значительный диурез после приема гипотонической жидкости вместе с алкоголем. Когда секреция АДГ уменьшается или прекращается, эпителий собирательных трубочек становится непроницаемым для воды, а моча в трубочках-умеренно концентрированной. Введение в этих условиях АДГ вызывает повышение проницаемости собирательных трубочек для воды и концентрирование мочи.
При введении больших количеств АДГ происходит выраженное сужение артерий и в результате повышение кровяного давления (вазопрессорное
ГЛАВА 17. ЭНДОКРИНОЛОГИЯ 391
действие гормона), из-за чего в англо-американской литературе обычно используют название «вазопрессин» (AVP = аргинин-вазопрессин). Резкое падение кровяного давления при кровопотере или шоке вызывает увеличение секреции АДГ и вследствие этого повышение кровяного давления. Вместе с тем в воротной системе печени АДГ вызывает снижение кровяного давления.
Осморецепторы и регуляция объема жидкости. Из
сказанного следует, что в организме должны существовать осморецепторы, способные определять существующее в данный момент осмотическое давление, а также барорецепторы, контролирующие степень наполненности кровеносных сосудов (и, следовательно, кровяное давление в системах с высоким и низким давлением), чтобы информация обоих видов поступала в клетки гипоталамуса, секретирующие АДГ. Осмотическое давление крови в основном, вероятно, регистрируется гипоталамусом. Возможно, осморецептивностью обладают сами клетки, секретирующие АДГ. Имеются указания на то, что осморецепторы присутствуют также в воротных сосудах между желудочно-кишечным трактом и печенью и сигнализируют В гипоталамус о местном осмотическом давлении. Барорецепторы, посылающие сигналы в нейроны, секретирующие АДГ, локализованы в каротидном и аортальном синусах, т.е. в участках с высоким артериальным давлением (с. 533). Аналогичные барорецепторы, выполняющие ту же функцию, находятся в грудном отделе, где давление крови низкое, и в предсердии (с. 535). Благодаря этим барорецепторам осуществляется тонкая регуляция секреции АДГ, обеспечивающая повышение его концентрации в крови при вертикальном и снижение при горизонтальном положении тела (рефлекс Гауэра-Генри; с. 540).
Патофизиологические аспекты. При нарушении секреции АДГ развивается заболевание, называемое несахарным диабетом (diabetes insipidus) (с. 798). Как следует из названия, при этом заболевании образуется большое количество мочи, не имеющей сладкого вкуса. Когда-то врачи, чтобы поставить диагноз, действительно пробовали мочу больного на вкус, и если она оказывалась сладкой, диагностировали сахарный диабет (diabetes mellitus-латинское mel означает сладость). Если же моча была очень разбавленной, она не имела сладкого вкуса; отсюда и возникло название заболевания. В те времена еще не было известно, что причиной заболевания является дефицит АДГ.
В настоящее время АДГ получают синтетическим путем, поэтому лечение редко встречающегося несахарного диабета не представляет серьезной проблемы.
Окситоцин
Окситоцин, подобно АДГ, образуется в крупных клетках супраоптического и паравентрикулярного
ядер гипоталамуса, транспортируется по их аксонам в заднюю долю гипофиза и здесь хранится. Органом-мишенью окситоцина служат миометрий (мышечной слой матки) и миоэпителий молочной железы.
«Рефлекс выделения молока». В физиологических условиях молочные железы начинают выделять молоко в течение 24 ч после родов, и в это время младенец уже может сосать. Акт сосания служит сильным раздражителем для сосков, имеющих обильную иннервацию. По нервным путям стимул передается в гипоталамические нейроны, вырабатывающие окситоцин. Окситоцин вызывает сокращения миоэпителиальных клеток, выстилающих железу; эти сократительные структуры в отличие от других сократительных структур организма-мышц имеют эпителиальное происхождение. Миоэпителиальные клетки располагаются вокруг альвеол железы, и при их сокращении молоко выдавливается. Таким образом, от младенца для извлечения содержимого железы не требуется активного сосания, поскольку ему помогает «рефлекс выделения молока».
В опытах на животных показано, что сосательный стимул активирует одновременно все нейроны, вырабатывающие окситоцин, который выделяется поэтому сразу в большом количестве (рис. 17.9). Это приводит к одновременному сокращению всех миоэпителиальных клеток и прогрессирующему повышению давления в молочной
 |
| Рис. 17.9. Схема рефлекса молокоотделения. Нейроны супраоптического и паравентрикулярного ядер, секретирующие окситоцин, периодически и синхронно активируются в результате механического раздражения шейки матки или сосков. Вследствие этого в кровь выбрасывается окситоцин. который, достигнув молочных желез, повышает в них давление молока. Под действием окситоцина активируется также мускулатура матки |
392 ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
железе. «Рефлекс выделения молока» запечатлен в живописи-на картинах, где мать смотрит на младенца и у нее из груди течет молоко. Клиницисты действительно знают, что после отнятия от груди лактация в присутствии младенца прекращается медленнее, чем в его отсутствие.
Действие на сеиситизированный миометрий. При
механическом раздражении влагалища и шейки матки возникают нервные импульсы, которые поступают в гипоталамус и вызывают выделение окситоцина {рефлекс Фергюсона). К концу беременности под действием эстрогенов резко повышается чувствительность миометрия к окситоцину. Приблизительно после 280-го дня беременности секреция окситоцина повышается, что приводит к слабым сокращениям миометрия, проталкивающим плод по направлению к шейке матки и влагалищу. Растяжение этих тканей регистрируют присутствующие в них многочисленные механорецепторы, и сигнал передается в гипоталамус. Последний отвечает высвобождением новых порций окситоцина, благодаря чему давление на механорецепторы еще более усиливается. В конечном итоге этот процесс переходит в роды, в ходе которых плод и плацента изгоняются. После изгнания плода раздражение механорецепторов шейки матки и влагалища прекращается, и выброс окситоцина временно прекращается.
Другие эффекты окситоцина. У небеременных женщин рефлекс Фергюсона не имеет большого значения, хотя окситоции. выделяющийся во время коитуса, может вызывать слабые сокращения матки, облегчающие продвижение спермы. Относительно роли окситоцина в организме мужчин пока ничего неизвестно. Возможно, он усиливает перистальтику мышцы, поднимающей яичко, во время эякуляции.