
Н. Н. Алипова, канд биол наук О. В. Левашова и канд биол наук М. С. Морозовой под редакцией акад. П. Г. Костюка москва «мир» 1996 ббк 28. 903 Ф50
| Вид материала | Документы |
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- М. А. Ляшко доц., канд физ мат наук; Т. Н. Смотрова доц., канд, 2299.13kb.
- Современные направления развития физической культуры, спорта и туризма, 4493.6kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Пособие для врачей и среднего медицинского персонала Минск, 5480.63kb.
- Ббк 63. 3(0) Н72, 4378.93kb.
- Тезисы докладов, 4290.75kb.
- Строительные нормы и правила нагрузки и воздействия сниП 01. 07-85* министерство строительства, 1162.86kb.
- Ббк 63. 3(0) Н72, 5546.58kb.
Глава 17. ЭНДОКРИНОЛОГИЯ. В. Вутке
17.1. Общая эндокринология
Гормоны как носители информации
Функции гормонов. Согласно классическому определению, гормоны-это продукты внутренней секреции, т.е. химические вещества, которые вырабатываются специализированными железами, выделяются в кровь и разносятся ею по телу. Область физиологии, связанная с изучением этих веществ, называется эндокринологией. Гормоны служат химическими носителями информации; достигнув органа-мишени, они оказывают на него специфическое воздействие. Специфичность действия гормонов обеспечивается присутствием в клетках молекулрецепторов. Рецепторами соответствующего гормона обладают только клетки органа-мишени, способные благодаря этому «считывать» химически закодированную информацию.
Под влиянием гормонов находятся те функции организма, для запуска или регуляции которых требуются минуты или часы. Таким образом, передача гормональной информации осуществляется в десятки раз медленнее, чем нервная передача, позволяющая организму немедленно реагировать на факторы окружающей среды или внутренние функциональные изменения.
Образование гормонов. Гормоны вырабатываются секреторными клетками. Такие клетки либо образуют компактные органы (железы), либо разбросаны по одной или в виде скоплений внутри органов, предназначенных для синтеза гормонов. Образовавшиеся гормоны хранятся в гранулах внутриклеточных органеллах, отделенных от цитоплазмы мембраной. В гранулах содержится большое число молекул гормона, погруженных в белковый матрикс. В ответ на специфический стимул гормон высвобождается, мембрана гранулы сливается с плазматической мембраной и в месте слияния образуется отверстие, через которое молекулы гормона выбрасываются в межклеточное пространство. Этот процесс называется экзоцитозом (с. 17). Гранулы и процесс экзоцитоза хорошо изучены морфологически. Процесс экзоцитоза гормонов из секреторных клеток подобен высво-
бождению нейромедиаторов из нервных окончаний.
Классификация гормонов. Поскольку передаваемая гормоном информация закодирована в его молекулярной структуре, для понимания механизма действия гормонов необходимо иметь хотя бы общее представление об их химическом строении. Более подробные сведения по этому вопросу можно найти в учебниках биохимии.
Все гормоны представляют собой либо белки (в том числе производные аминокислот), либо липиды. В зависимости от локализации рецепторов в клетках-мишенях гормоны можно разделить на три группы [27, 32].
Первую группу составляют гормоны липидной природы. Будучи жирорастворимыми, они легко проникают через клеточную мембрану и взаимодействуют с рецепторами, локализованными внутри клетки,-как правило, в цитоплазме (рис. 17.1).
Вторая группа - белковые и пептидные гормоны. Они состоят из аминокислот и по сравнению с гормонами липидной природы имеют более высокую молекулярную массу и менее липофильны, из-за чего с трудом проходят через плазматическую мембрану. Рецепторы этих гормонов находятся на поверхности клеточной мембраны (рис. 17.1), так что белковые и пептидные гормоны в клетку не проникают.
Третью химическую группу гормонов составляют низкомолекулярные тиреоидные гормоны, образованные двумя аминокислотными остатками, связанными между собой эфирной связью. Эти гормоны легко проникают во все клетки тела и взаимодействуют с рецепторами, локализованными в ядре (рис. 17.1). Одна и та же клетка может иметь рецепторы всех трех типов, т.е. локализованные в ядре, цитозоле и на поверхности плазматической мембраны. Кроме того, в одной и той же клетке могут присутствовать разные рецепторы одного типа; например, на поверхности клеточной мембраны могут находиться рецепторы разных пептидных и/или белковых гормонов.
Механизмы действия гормонов. Главное условие осуществления всех эндокринных функций - это присутствие в клетках-мишенях специфических рецеп-
ГЛАВА 17. ЭНДОКРИНОЛОГИЯ 385
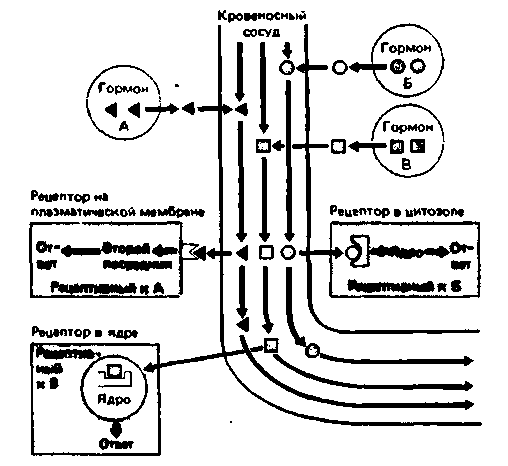 |
| Рис. 17.1. Механизмы действия гормонов. Три эндокринные клетки секретируют в межклеточное пространство в непосредственной близости от капилляров гормоны А, Б и В. Молекулы гормонов диффундируют в кровеносный сосуд и доставляются кровью к клеткам мишеням, содержащим рецепторы этих гормонов. Гормон А взаимодействует с рецептором, находящимся на поверхности плазматической мембраны. Гормон-рецепторный комплекс стимулирует образование вторичного посредника, который индуцирует ответ кпетки. Гормон Б взаимодействует с рецептором, находящимся в цитоплазме. Образующийся комплекс транслоцируется в ядро и воздействует на геном (например, изменяет синтез ядерной ДНК), что приводит к изменению синтеза белка. Гормон В взаимодействует с рецептором, локализованным в ядре, т.е. действует практически так же, как гормон Б, с той только разницей, что вго рецептор исходно локализован в ядре |
торов, позволяющих считывать информацию, закодированную в гормоне. При взаимодействии гормона с рецептором, находящимся в цитоплазме, в ядре или на поверхности плазматической мембраны, образуется гормон-рецепторный комплекс (рис. 17.1) [27].
Существуют два механизма действия гормонов, принципиально различающихся по признаку того, где образуется гормон-рецепторный комплекс - внутри клетки или на ее поверхности.
Гормон-рецепторный комплекс, образующийся внутри клетки, может непосредственно влиять на экспрессию генетической информации, т. е. сам оказывает действие на геном и в результате на процессы синтеза в клетке (рис. 17.2). Таким образом, гормон-рецепторный комплекс может индуцировать или подавлять синтез белка [27].
Иначе действует гормон-рецепторный комплекс,
образующийся на поверхности клетки. Взаимодействие гормона с рецептором, локализованным на плазматической мембране, влияет на активность клетки посредством сложного биохимического механизма, как правило, с участием второго посредника внутри клетки. Наиболее известным примером активации второго посредника служит образование циклического аденозинмонофосфата (цАМФ) из аденозинтрифосфата (АТФ). Гормон-рецепторный комплекс активирует аденилатциклазу, которая катализирует дефосфорилирование АТФ и превращение его в цАМФ (рис. 17.3). На последующих этапах цАМФ влияет на функциональную активность клетки [8]. Например, цАМФ может изменять проницаемость клеточной мембраны для ионов, и тогда в ответ на гормональный стимул меняется скорость клеточной секреции.
К числу других веществ, способных, по-видимому, действовать в качестве вторых посредников, относятся циклический гуанозинмонофосфат (цГМФ), кальмодулин и фосфоинозитол. В случае этих соединений высокоспецифичное действие гормон-рецепторного комплекса на клетку опосредовано неспецифическим внутриклеточным механизмом-образованием одного или нескольких вторых посредников.
 |
| Рис. 17.2. Взаимодействие гормона с внутриклеточным рецептором. Гормон диффундирует через плазматическую мембрану и взаимодействует с рецептором. Гормон-рецепторный комплекс переносится в ядро и действует на синтез ДНК, изменяя скорость транскрипции и количество информационной (матричной) РНК (мРНК). Увеличение или уменьшение количества мРНК влияет нв синтез белка в процессе трансляции, что приводит к изменению функциональной активности клетки |
386 ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
 |
| Рис. 17.3. Механизм действия гормона, образующего комплекс с рецептором, локализованным на плазмати• ческой мембране. Достигнув рецептора и взаимодействуя с ним, гормон вызыввет его трансформацию. Трансформация приводит к активации аденилатциклвзы, также локализованной в плазматической мембране. Аденилатциклаза катализирует дефосфорилирование АТР с образованием сАМР. сАМР вызывает в клетке разнообразные эффекты, которые приводят в конечном счете к физиологическому ответу клетки. (АТР-аденозинтрифосфат, АТФ; сАМР-циклический аденозинмонофосфат, цАМФ) |
Синтез и распад гормонов
Синтез гормонов. Синтез гормонов липидной природы и гормонов - производных аминокислот (тиреоидных гормонов) будет рассмотрен в разделе, посвященном их функциям (с. 397 и 404). Белковые и пептидные гормоны образуются в аппарате Гольджи путем трансляции генетической информации, закодированной в РНК, в аминокислотную последовательность. Синтез этих гормонов ничем не отличается от синтеза других белков тела. Следует, однако, сказать, что исходно гормон образуется не в биологически активной форме, а в виде более крупной неактивной молекулы, так называемой препроформы гормона [4]. Помимо аминокислотной последовательности самого гормона в этой молекуле содержится ряд дополнительных остатков, которые отщепляются под действием ферментов на этапе, называемом посттрансляционным процессингом. В окончательном виде гормон упаковывается в гранулы и хранится в них готовым для секреции. Таким образом, в процессе синтеза белковых и пептидных гормонов образуются не только сами гормоны, но и добавочные пептиды. О биологической функции этих пептидов пока известно немногое, но становится все более очевидным, что в некоторых системах эти «остатки» имеют определенное биологическое значение. В качестве примера ниже будет рассмотрен проопиомеланокортин (с. 400) [4, 7, 12].
Распад гормонов. Белковые и пептидные гормоны быстро распадаются в организме при участии двух принципиально разных механизмов.
- Многие молекулы гормонов, присутствующие в крови, никогда не вступают во взаимодействие с рецепторами, при этом они, что важно, не должны циркулировать слишком долго. В различных органах (печени, легких, мозгу, почках) [12] существует ряд ферментных систем, участвующих в расщеплении белковых и пептидных гормонов. Поскольку связывание гормона с рецептором обратимо, часть связавшихся с рецепторами молекул в конечном счете высвобождается и также подвергается ферментативному расщеплению.
- Некоторые гормоны после взаимодействия с рецепторами подвергаются расщеплению внутри самой клетки-мишени, находясь в комплексе с рецептором, или после отщепления от него; распад происходит в лизосомах.
Новые аспекты эндокринологии
Паракринное действие гормонов. Прежде чем перейти к рассмотрению отдельных гормональных систем, следует остановиться на недавних открытиях, заставляющих пересмотреть многие традиционные представления и ввести новые понятия. Согласно принятому определению, гормоны - это носители химической информации, вырабатываемые секреторными клетками и выделяемые в кровь, которая доставляет их к органу-мишени. Характерный признак органа-мишени - это способность считывать информацию, закодированную в гормоне. В некоторых случаях информация может считываться клетками, находящимися в непосредственной близости от клетки, секретирующей гормон. При этом гормон просто диффундирует через межклеточное пространство к клетке-мишени, т.е. не является гормоном в классическом виде, поскольку не переносится кровью. Когда такие носители информации действуют на соседние клетки, их называют паракринными гормонами или гормонами местного действия [2]. Иногда их называют также тканевыми гормонами. Еще раньше это название было закреплено за простагландинами, которые, как недавно установлено, действуют именно таким образом. Теперь мы знаем, что некоторые классические гормоны также могут действовать паракринным путем, а следовательно, могут называться «тканевыми гормонами». На рис. 17.4 проведено сравнение между гормональной и паракринной регуляцией. Сейчас уже известно, что наряду с классическими гормонами существует множество веществ, обладающих паракринным действием. В принципе к таким веществам можно отнести и классические нейромедиаторы, с той только
ГЛАВА 17. ЭНДОКРИНОЛОГИЯ 387
 |
| Рис. 17.4. Различие между гормональной и паракринной регуляцией. Железистая клетка А вырабатывает гормон, который в соответствии с классическим определением доставляется к клетке-мишени кровью. Железистая клетка Б вырабатывает гормон, обладающий паракринным действием, т.е. влияющий на соседние клетки. Этот же гормон, однако, может доставляться к органу-мишени и кровью |
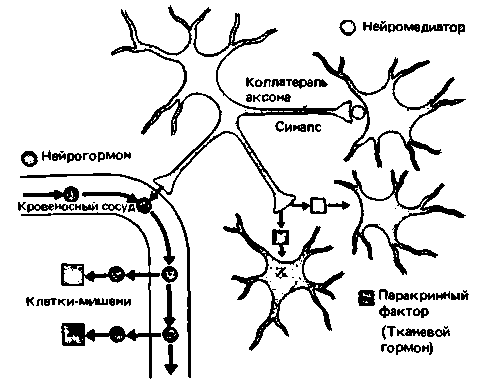 |
| Рис. 17.5. Различие между нейромедиатором, нейрогормоном и паракринным нейрогормоном. Нейрон вырабатывает некое вещество. Если оно выделяется в синаптических окончаниях, то соответствует определению нейромедиатора. Аксон того же нейрона может иметь коллатераль, которая оканчивается на кровеносном сосуде и высвобождает это же вещество в кровоток - в таком случае оно ведет себя как нейрогормон. Из другой коллатерали аксона то же вещество диффундирует в окружающую ткань и действует на соседнюю группу клеток, т. е. обладает паракринным действием |
разницей, что источником химической информации в этом случае являются не специализированные клетки внутренней секреции, а нервные клетки. Нейромедиаторы не поступают в кровь, а диффундируют через узкую синаптическую щель в сторону постсинаптической нервной клетки. На постсинаптической мембране нейромедиатор, как и гормон, связывается со специфическим рецептором.
Нейрогормоиы. Недавно было установлено, что нервные клетки также могут вырабатывать пептидные и белковые гормоны [7, 12] и часто выделяют свои секреты в кровь, т.е. продуцируют нейрогормоны. Таким образом, следует признать, что большие участки центральной нервной системы способны выполнять эндокринную функцию. В чем же тогда заключается основное различие между гормоном и нейромедиатором? Оно состоит в том. что нейромедиатор диффундирует через синаптическую щель, тогда как нейрогормон попадает в органмишень через кровеносную систему. В некоторых случаях терминали аксона нервной клетки выделяют вырабатываемое ею вещество в виде нейромедиатора, а коллатерали аксона того же нейрона оканчиваются на кровеносном сосуде и высвобождают то же вещество в виде нейрогормона (рис. 17.5).
Поскольку выделяемые пептиды влияют на
активность соседних нервных клеток, их следует также считать нейромедиаторами. Нейрофизиологические опыты показали, что нейроны, выделяющие тот или иной классический нейромедиатор, могут быть подразделены на субпопуляции, высвобождающие разные нейропептиды. О функциях этих субпопуляций известно мало. В настоящее время представляется вероятным, что многие нейроны способны вырабатывать помимо классических нейромедиаторов один или несколько пептидов.
Методы изучения
В 1848 г. А. Бертольд опубликовал опыты, положившие начало современным исследованиям гормонов. К тому времени было известно, что у петухов после кастрации атрофируется гребень и поведение становится менее агрессивным. При этом существовало мнение, что связь между семенниками и центральной нервной системой осуществляется при помощи нейронов. Бертольд кастрировал петухов и имплантировал им семенники в брюшную полость. У таких петухов сохранялся высокий гребень и агрессивное доминантное поведение. Этот опыт показал, что из семенников в центральную нервную систему поступает один или несколько гуморальных сигналов. Аналогичные подходы используют и сегодня при изучении характера действия гормонов. С этой целью удаляют орган, секретирующий гормон, и наблюдают за последствиями. Действие гормона можно также изучать с помощью биологических методов, основанных на введении
388 ЧАСТЬ IV. ПРОЦЕССЫ НЕРВНОЙ И ГУМОРАЛЬНОЙ РЕГУЛЯЦИИ
гормона подходящему животному и анализе биологического эффекта [3].
Подобные методы обладают, однако, крайне низкой чувствительностью, поэтому они к настоящему времени почти полностью вытеснены высокочувствительными иммунологическими методами, позволяющими получать специфические антитела, которые связываются только с данным гормоном и не взаимодействуют с его предшественниками или продуктами распада, а также с другими сходными молекулами. Комплекс гормона с антителом можно осадить и определить количественно. Этим методом проводят некоторые полуколичественные тесты (например, тесты на беременность).
Еще более высокой чувствительностью обладают методы радиоиммунологического анализа (РИА). Они основаны на использовании специфических антител, которые инкубируют со смесью радиоактивно меченных и немеченых молекул гормона. При этом общее число молекул гормона превышает связывающую способность антител, благодаря чему происходит конкуренция за места связывания. Молекулы гормона, связавшиеся с антителами, и свободные молекулы разделяют, затем измеряют радиоактивность связанной фракции и по калибровочной кривой определяют количество гормона в биологическом образце (подробное описание метода см. в руководствах по клинической химии). С помощью таких современных методов можно измерять ничтожно малые количества гормона (вплоть до фемтомолей).
Эндокринные цепи регуляции
Системы обратной связи. Эндокринный контроль можно рассматривать как цепь регуляторных эффектов (с. 333), в которой результат действия гормона прямо или косвенно влияет на элемент, определяющий содержание доступного гормона. Как правило, такое взаимодействие происходит по принципу отрицательной обратной связи и заключается в том, что при воздействии гормона на клетки органа-мишени их ответ, влияя на источник секреции гормона, вызывает ее подавление. Схематически такая система представлена на рис. 17.6. Сигнал отрицательной обратной связи может иметь гуморальную или нервную природу. Крайне редко встречается положительная обратная связь, когда ответ клетки-мишени на действие гормона в свою очередь вызывает усиление секреции.
В любом случае передача сигнала обратной связи на том или ином участке осуществляется химическим путем. В регуляторных эффектах нервной системы в качестве посредников участвуют нейромедиаторы. В тех случаях, когда посредниками в механизме обратной связи служат гормоны, контролирующий элемент должен содержать клетки, имеющие рецепторы этих гормонов; роль сигнала обратной связи выполняет концентрация самого гормона. В простейшем варианте обратной связи гормон регулирует скорость своей секреции через так называемые ауторецепторы. Этот механизм можно рассматривать как аналог пресинаптического торможения (самоторможения) выделения нейромедиаторов (с. 353).
 |
| Рис. 17.6. Схематическое изображение эндокринной цепи регуляции. Контроллер сравнивает истинную величину контролируемой переменной с «заданным значением» и посылает сигнал, вызывающий соответствующие изменения функции эндокринной железы. Скорость секреции гормонов эндокринной железой может изменяться под влиянием различных возмущающих факторов. Секретируемые железой гормоны регулируют систему, отвечающую на гормональную информацию соответствующим физиологическим эффектом. Одновременно сигнал о новом значении контролируемой переменной поступает в контроллер, который и замыкает цепь |
Значительно более распространен другой вид цепи эндокринной регуляции, в которой гормон А стимулирует секрецию гормона Б, а гормон Б угнетает секрецию гормона А. Обычно, однако, такие системы устроены гораздо сложнее и включают как гуморальные, так и нервные сигналы, передаваемые более чем одним посредником (нейромедиатором или гормоном).
Нейроэндокринная цепь регуляции. Нейроэндокринология изучает взаимодействие приносимых кровью гормонов с центральной нервной системой и ответ последней на эти гормоны. Ответ включает целый комплекс изменений в образовании и высвобождении нейромедиаторов и/или нейрогормонов и имеет результатом увеличение или уменьшение секреции других гормонов. Нейроэндокринологи изучают не только регуляцию уровней гормонов, осуществляемую цепями регуляции в центральной нервной системе, но и соответствующие изменения форм поведения (например, связь между секрецией гонад и половым поведением, регуляцией уровня глюкозы в крови и чувством голода, осморегуляцией и чувством жажды).
В нейроэндокринных цепях регуляции обратную связь с центральной нервной системой осуществля-
ГЛАВА 17. ЭНДОКРИНОЛОГИЯ 389
ют гормоны, секрецию которых стимулируют продукты, выделяемые передней долей гипофиза (аденогипофиза). Следовательно, должны существовать нейроны, чувствительные к этим гормонам. Действительно, как было установлено, тиреоидные гормоны, а также стероидные гормоны половых желез и надпочечников могут влиять на активность нейронов в различных участках мозга. Особенно хорошо изучены нейроны, «читающие» стероидные гормоны. Рецепторы стероидов в этих нейронах, подобно другим рецепторам липидных гормонов, находятся в цитоплазме1'. Гормон-рецепторный комплекс действует непосредственно на геном, стимулируя или угнетая его активность, в результате чего происходят изменения в метаболизме нервной клетки. При этом может меняться ее электрическая активность, так что сдвиги в концентрации стероида в крови могут влиять и на нейроны, которые непосредственно с ним не взаимодействуют. Кроме того, в центральной нервной системе существуют нейроны, которые влияют на клетки, секретирующие стероидные гормоны, не прямо, а чаще всего через гипофиз. Информация об уровне стероидного гормона в крови поступает в мозговые центры, влияющие на поведение, и таким путем происходит координация между гормональным статусом и поведением.