
Динамика популяций и сообществ мелких млекопитающих как показатель состояния лесных экосистем (на примере Каспийско-Балтийского водораздела)
| Вид материала | Автореферат |
- Методика комплексной оценки состояния сообществ и популяций доминирующих видов или, 1278.81kb.
- Эколого-морфологическая характеристика размножения мелких млекопитающих из популяций,, 635.9kb.
- «Оценка экологического состояния экосистем и растительных сообществ водно-болотных, 3457.77kb.
- Сухачев александр Николаевич динамика клеточных популяций гемоцитов асцидии, 331.84kb.
- Биотехнология в воспроизводстве и селекции крупного рогатого скота имеет особое значение., 299.72kb.
- Использование природного капитала лесных экосистем с учетом трансграничных факторов, 613.87kb.
- Объявляет конкурс на позицию постдока в области Тканевой инженерии и Регенеративной, 191.04kb.
- А. В. Марков Эволюционный прогресс Конспект, 153.21kb.
- Правила охраны и промысла морских млекопитающих в Дальневосточном рыбохозяйственном, 336.4kb.
- М. А. Давыдов Кпроблеме благосостояния крестьян в пореформенной России, 310.26kb.
Исследованы особенности динамики численности и фенетического разнообразия популяций ценозообразующих видов с разными пространственными структурами и стратегиями в условиях европейской южной тайги (рыжей и красной полевок, полевки-экономки). Выявлены тренды, периодические высокочастотные и низкочастотные колебания многолетних временных рядов численности модельных видов. Произведен расчет их примерного вклада в общее варьирование численности и построены статистические модели. Описаны неметрические вариации признаков и составлены региональные каталоги фенов. Определены наиболее полиморфные признаки, которые могут иметь индикационную значимость при изучении динамики фенетического разнообразия. Выявлены пространственно-биотопические, временные и внутрипопуляционные особенности формирования фенетической структуры и разнообразия модельных видов в различных экологических условиях.
Популяционная динамика рыжей полевки в естественных и нарушенных лесных экосистемах. Основной модельный вид на онтогенетическом и популяционном уровне исследований - рыжая полевка, которая в лесных массивах формирует континуальные популяции.
Динамика численности. В период 1981-2007 гг. динамика численности рыжей полевки в коренных ельниках представляла собой нестационарный процесс (рис. 9).
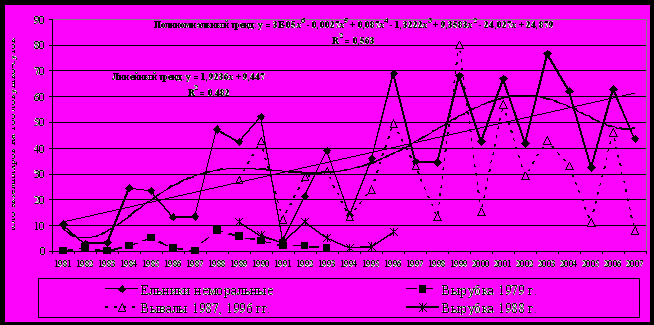 |
| Рис.9. Многолетняя динамика и тренды численности рыжей полевки в неморальных ельниках, на участках массовых ветровалов и на сплошных вырубках |
Существуют положительные тренды, на долю которых приходится более 50% варьирования исходных рядов. Достаточно реалистично эти зависимости описываются полиномиальными моделями шестой степени. На фоне увеличения численности наблюдаются периодические ее колебания. Отмечено явное упрощение структуры циклов за данный промежуток времени: от 4-летних с типичным S-образным ростом к трехлетним, а затем к простым двухгодичным. Переходы к новым режимам популяционной динамики произошли после катастрофических ветровалов 1987 и 1996 гг. Одним из триггерных механизмов смены популяционных циклов можно считать подснежное размножение, которое было зарегистрировано зимой 1987-1988 гг., вероятно, как ответная реакция на значительное и резкое нарушение стационарности экосистем в результате широкомасштабного катастрофического ветровала. Установлено также, что конкретные сочетания климатических характеристик могут оказывать заметное воздействие на уровень численности в различные фазы популяционных циклов (Минаева, Истомин и др., 2001; Истомин, 2005, 2007). В крупном лесном массиве многолетние и сезонные изменения численности в разных типах коренных и производных местообитаний тесно сопряжены (r=0,74). Существенные перестройки общего и сезонного хода динамики численности рыжей полевки происходят на ранних стадиях возобновления вырубок. Средние показатели летней численности вида гораздо ниже (в 6-10 раз), синхронность изменений выражена слабо. В ненарушенных ельниках сезонный пик достигается в середине-конце лета, а на ранних стадиях вырубок – осенью. Это, вероятно, связано с существенным вкладом процесса расселения в динамику численности данного вида на вырубках и притоком особей из лесных экосистем, окружающих вырубленные площади (Истомин, 1999).
Динамика фенотипического разнообразия. Для 40 исследованных элементарных признаков черепа рыжей полевки выявлено 162 фена; для параконида и талонуса - 13 комплексных морфотипов.
Важнейшая характеристика популяций – ее пространственная структура, от которой в значительной степени зависят характер и скорость микроэволюционных событий. Основные представления о типах популяционной структуры и соответствующие теоретические модели были разработаны достаточно давно (Wright, 1951, 1969; Kimura, 1953; Kimura, Weiss, 1964; Malecot, 1967, 1975 и др.). Однако характер эволюционных преобразований в больших непрерывных и в совокупности мелких дискретных популяций до сих пор принадлежит к числу мало изученных вопросов. При исследовании континуальной популяции рыжей полевки нами обнаружено, что в годы высокой численности в диапазоне удаленностей до 4 км пространственные группировки не являются строго изолированными расстоянием. Корреляция между величиной фенетической дистанции и расстоянием по комплексу признаков была недостоверной (r=0,20). Число фенов на отдельных участках варьировало незначительно, доминирующие морфотипы в основном также были одинаковы. Несколько больший уровень различий получен между группировками, населяющими разные типы местообитаний, вне зависимости от расстояния между ними. Наибольшей специфичностью отличались бореальные ельники. Фенетические дистанции между группировками заметно возрастали в годы низкой численности, что вероятно, в первую очередь определяется меньшей интенсивностью генетического обмена между ними в эти периоды (рис. 10).
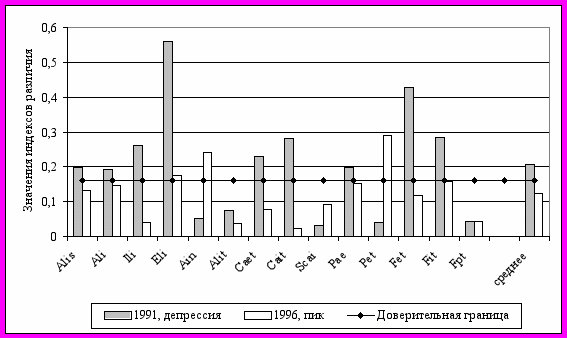 | Рис. 10. Фенетические дистанции между популяционными группировками рыжей полевки из неморальных и бореальных ельников в годы с низкой и высокой численностью |
Сравнивали популяционно-феногенетические особенности рыжей полевки в крупном сплошном лесном массиве и в системе мелких лесных резерватов, изолированных верховыми болотами. В биотопическом плане, с позиций единовременности и пространственной разобщенности расстоянием субпопуляции «болотных изолятов» и территориальные группировки непрерывной популяции находились в относительно сходных условиях. Однако своеобразие сравниваемых выборок из континуальной и дискретной популяций достаточно велико: из 32 признаков по 18 получены статистически значимые отличия. Обнаружено, что в дискретной популяции заметно выше общий уровень фенотипического разнообразия, но ниже доля редких фенов. Для совокупности популяций «болотных изолятов» отмечено достоверное увеличение флуктуирующей асимметрии для качественных и количественных признаков. Это может свидетельствовать о том, что возрастание общего фенотипического разнообразия происходит, прежде всего, за счет увеличения уровня случайных изменений развития. Однако для некоторых признаков наблюдается независимость изменения величин фенотипического разнообразия и флуктуирующей асимметрии. Поэтому нельзя полностью исключать роль генетической составляющей в повышении общего фенотипического разнообразия. Следовательно, на основании косвенных данных частично подтверждается известный факт увеличения суммарной генетической изменчивости в совокупности подразделенных популяций (Ланде, Бэрроуклафф, 1989). Подобный результат вполне может рассматриваться как одна из форм компенсационных явлений в ответ на видовое обеднение биоты (Чернов, 2005), что наблюдается в мелких лесных резерватах. В целом полученные различия свидетельствуют о разной микроэволюционной ситуации в рассматриваемых популяциях. Ранее нами было показано, что даже очень близкие виды могут быть отличны в своих отношениях к основным характеристикам изолятов: площади, гетерогенности среды, степени изоляции от источников иммиграции (Истомин, 1984, 1990). Поэтому вполне вероятно ожидать неодинаковые направленности и скорости микроэволюционных процессов различных видов в условиях инсуляризации.
Обнаружено, что массовые ветровалы и сплошные вырубки весьма существенным и сходным образом могут изменять фенетическую структуру и популяционное разнообразие типично лесного вида рыжей полевки (рис. 11).
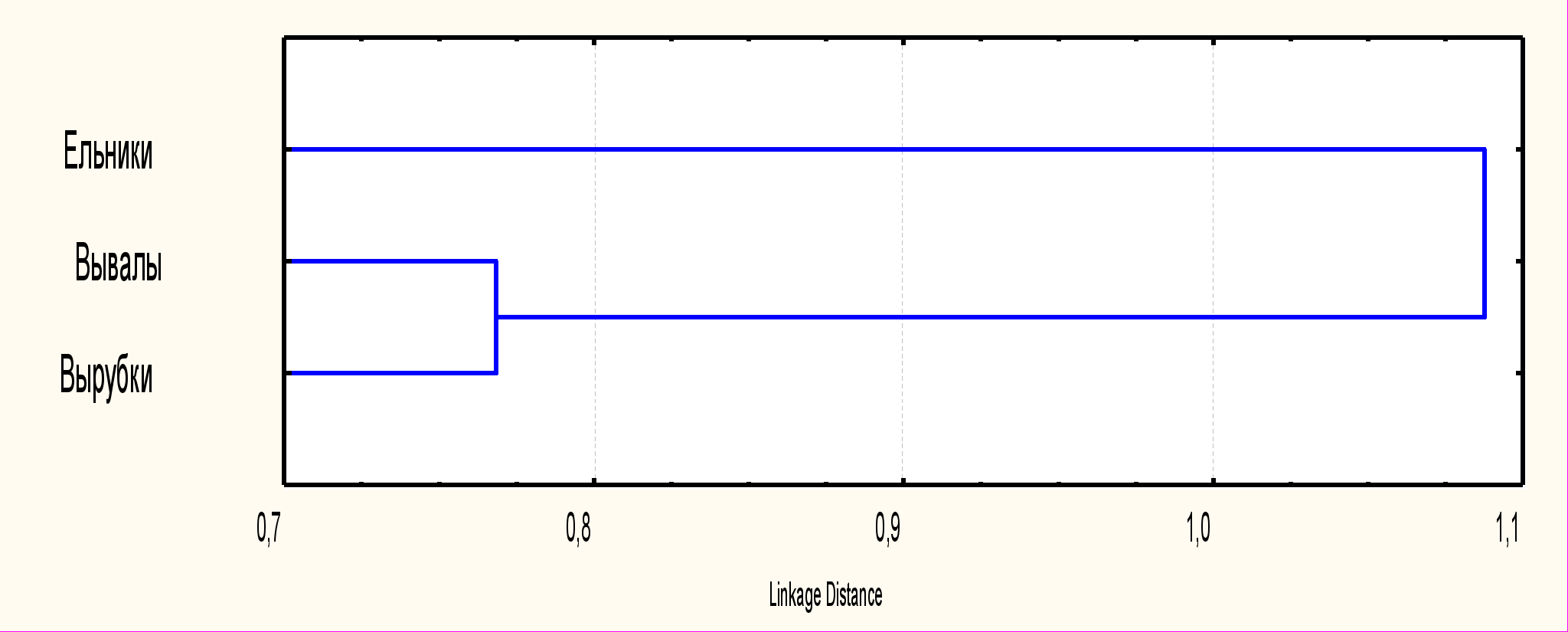 | Рис. 11. Дендрограмма сходства фенетической структуры популяционных группировок рыжей полевки из сравниваемых типов местообитаний |
Между группировками вывалов и вырубок существует гораздо большее сходство, чем при их сравнении с ненарушенными ельниками, несмотря на более существенные изолирующие расстояния. Степень фенетических дистанций между группировками ельников и вывалов значимо повышается в годы низкой численности. Под влиянием рассматриваемых форм нарушения зрелых лесных экосистем происходит некоторое уменьшение общей степени популяционного разнообразия или модифицируется его структура (табл. 2, 3).
Таблица 2. Феногенетическое разнообразие и скоррелированность развития признаков черепа в группировках рыжей полевки из различных местообитаний (серии 1991 г.)
| Показатели феногенетического разнообразия и скоррелированность развития признаков | Ельники n=72 | Вывалы n=110 | Вырубки n=94 |
| Общее число морф | 57 | 51 | 55 |
| Сумма значений общего показателя разнообразия | 44,85 | 39,09 | 43,24 |
| Среднее значение общего показателя разнообразия | 2,80 | 2,44 | 2,70 |
| Средняя доля редких фенов | 0,198 | 0,201 | 0,195 |
| Сумма значений флуктуирующей асимметрии | 177 | 281 | 293 |
| Среднее значение флуктуирующей асимметрии | 11,1 | 17,6 | 18,3 |
| Доля достоверных коэффициентов корреляций | 84% | 76% | 66% |
| Среднее значение коэффициентов корреляций | 0,61 | 0,52 | 0,49 |
Внутрииндивидуальное разнообразие в среднем всегда достоверно выше в группировках, заселяющих вывалы и вырубки, что указывает на снижение стабильности онтогенеза и совершенства его регуляторных процессов у типичного лесного вида в дестабилизированных экосистемах, по сравнению с климаксными равновесными ельниками.
Таблица 3. Феногенетическое разнообразие группировок рыжих полевок из коренных ельников и участков сплошных вырубок (серии 1984-1988 гг.; * - различия достоверны)
| Показатели | Признаки | Ельники | Вырубки |
| Показатель флуктуирующей асимметрии | Параконид М1 * | 14,3 (n=919) | 25,0 (n=44) |
| Талонус М3 ** | 12,9 (n=855) | 28,6 (n=49) | |
| Число морф | Параконид М1 | 6 (n=990) | 5 (n=224) |
| Талонус М3 | 7 (n=878) | 7 (n=235) | |
| Среднее значение общего показателя разнообразия | Параконид М1* | 4,26±0,15 | 3,60±0,23 |
| Талонус М3 | 4,25±0,26 | 3,70±0,26 | |
| Доля редких фенов | Параконид М1 | 0,15±0,03 | 0,14±0,06 |
| Талонус М3* | 0,34±0,04 | 0,15±0,06 |
Это дополнительно подтверждается снижением скоррелированности развития линейных признаков черепа (Истомин, 2001, 2007). Несогласованность изменений внутрииндивидуального и популяционного разнообразия косвенно может свидетельствовать об определенном вкладе генетической составляющей в уменьшение общего фенотипического разнообразия популяций типично лесного вида на участках массовых ветровалов и сплошных вырубок.
Более подробно исследована динамика фенетического разнообразия рыжих полевок в 1982-1992 гг. в коренных неморальных ельниках. В этот период укладывается несколько популяционных волн: два полных цикла с типичным S-образным ростом (1982-1985 гг., 1986-1990 гг.) и начало третьего цикла (см. рис 9). Полученные результаты указывают на достаточно высокую степень фенетических отличий популяционных группировок в различные годы, как по отдельным признакам, так и по их комплексу (рис. 12).
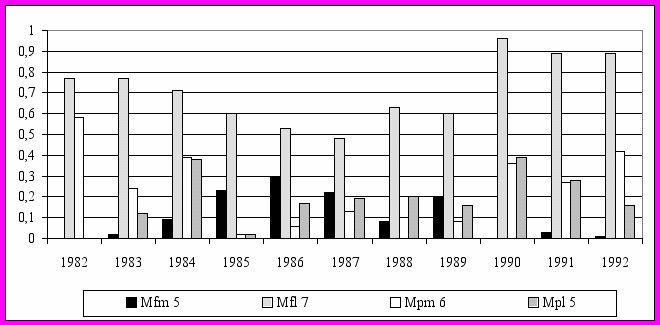
Рис. 12. Динамика частоты встречаемости некоторых фенов в группе вариабельных краниологических признаков
Обнаружено, что наиболее существенные перестройки феногенетического состава происходили на последней фазе 4-5 летних популяционных циклов (1985, 1990 гг.) перед снижением численности (рис. 13). Причем, степень фенетического сходства выборок последнего года популяционных циклов была несколько выше с начальной фазой следующего цикла, чем с предшествующим годом своего цикла. В рамках одного цикла прослеживаются закономерные изменения частоты встречаемости отдельных фенов в зависимости от уровня численности. Отмечена определенная периодичность формирования фенетической структуры популяции рыжей полевки в ходе многолетней динамики численности (см. рис. 12), что может свидетельствовать об устойчивости феногенетического разнообразия вида в коренных лесах.
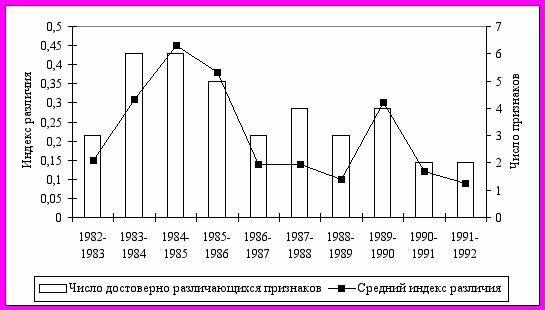
Рис. 13. Динамика степени различий фенетической структуры популяций рыжей полевки
Для оценки влияния на динамику фенетической структуры процесса воспроизводства и выживаемости были сформированы серии черепов перезимовавших животных и неполовозрелых сеголеток из осенних отловов (октябрь), уходящих на зимовку за период 1990-1993 гг. Обнаружено, что специфика воспроизводства может определять довольно быстрые и значимые изменения феногенетической структуры в ходе одного репродуктивного сезона. Наиболее существенные перестройки зарегистрированы в сезон с активным участием в размножении сеголеток ранних сроков рождения. При этом в определенной степени в репродукции выражен возрастной кросс, а осенние выборки неполовозрелых особей состоят из двух поколений. Выявлены изменения фенетической структуры в зависимости от условий перезимовки популяции. При сравнении осенних и весенних выборок для ряда признаков отмечены вариации, устойчиво из года в год демонстрирующие увеличение доли своего присутствия от осени к весне. Характер изменения частоты встречаемости данных фенов не зависел ни от специфики климатических условий, ни от плотности популяции. Это косвенно может свидетельствовать о хорошей выживаемости особей-носителей данных фенов, которые можно считать своеобразными маркерами животных с высокой неизбирательной жизнеспособностью. В то же самое время были встречены фены, частота которых в популяции от осени к весне следующего года менялась неодинаковым, но вполне закономерным образом, демонстрируя определенную связь со степенью благоприятности условий перезимовки. В свою очередь итоговая смертность популяции, безусловно, зависит от конкретного соотношения феногенотипов, имеющих неодинаковую выживаемость при различных сочетаниях погодных и других факторов. Указанные обстоятельства, несомненно, должны учитываться при анализе и прогнозировании динамики популяций.
В настоящее время практически не изучены аспекты феногенетики переносчиков зоонозных заболеваний на внутри- и межпопуляционном уровнях. В работе предпринимается попытка фенетического тестирования особей рыжей полевки по степени их восприимчивости и инфекционной чувствительности к основным возбудителям природно-очаговых болезней региона и оценивается влияние эпизоотий на фенетическое разнообразие популяций вида. Обнаружено, что фенетическая структура группировок особей, зараженных лептоспирозом, достоверно отличалась от таковой неинфицированных особей. Группировка особей, инфицированных ГЛПС, имела меньший уровень отличий от незараженных животных и величина среднего значения фенетических дистанций не достигала порогового уровня значимости. Отмечено снижение доли редких фенов в инфицированных группах особей. В целом, гораздо более существенно воздействие лептоспирозов, поскольку вмешивается фактор патогенеза при заражении особей, который снижает стабильность их индивидуального развития (рис. 14) и может увеличивать смертность конкретных феногенотипов в популяции. Полученные результаты дают основание предполагать, что феногенетическая структура популяций основного носителя инфекций может оказывать существенное воздействие на общий ход течения эпизоотии. Это, по-видимому, связано с различной инфекционной чувствительностью определенных феногенотипов особей.
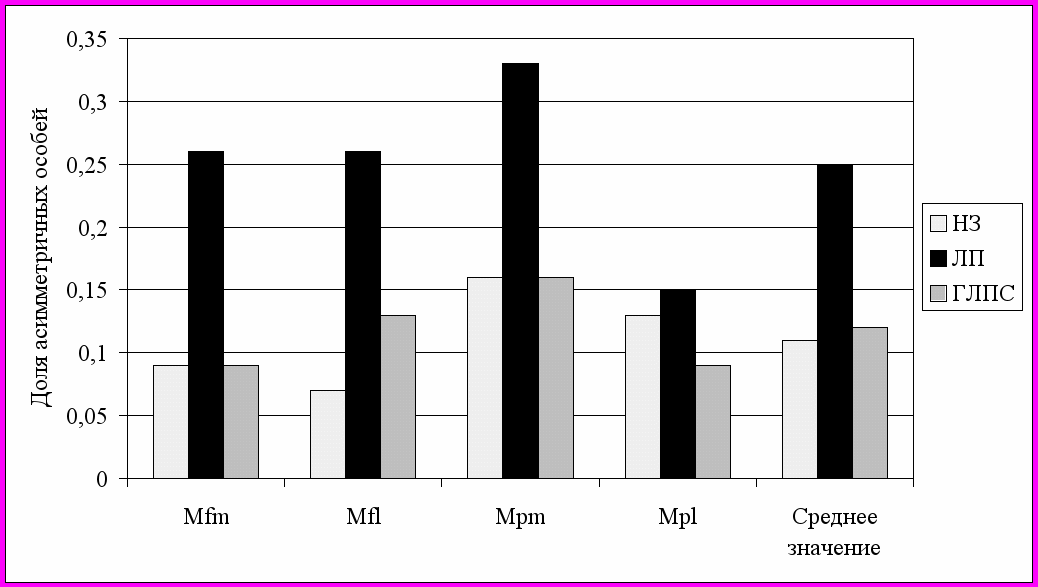 | Рис. 14. Доля асимметричных особей в инфицированных и неинфицированных группировках рыжей полевки по наиболее полиморфным признакам; НЗ – незараженные особи, ЛП – инфицированные лептоспирозом; ГЛПС – инфицированные геморрагической лихорадкой. |
Для мышевидных грызунов характерна сложная функциональная структурированность популяций. В данном разделе рассматриваются фенетические особенности оседлых и расселяющихся рыжих полевок, а также особей с различными типами онтогенеза. Роль указанных структурно-функциональных группировок грызунов в динамике ведущих популяционных параметров, безусловно, очень велика.
Процесс расселения во многом определяет пространственную структуру и жизнеспособность популяций. Он чрезвычайно сложен для исследований, поэтому существует много нерешенных проблем по разным его аспектам (Myers, Krebs, 1971; Hilborn, 1975; Fairbairn, 1978; Коли, 1979; Staffard, Staout, 1983; Gliwicz, 1986; Rease et all, 1989; Лукьянов, Лукьянова, 1996; Лукьянов, 1997 и др.). Генетическая обусловленность расселения является одним из наименее изученных вопросов. Нами установлено, что в континуальной популяции рыжей полевки группировки оседлых и расселяющихся особей фенетически разнородны (рис. 15). В то же самое время различия фенетической структуры расселяющихся особей, отловленных на разных участках и в разные периоды расселения, выражены несущественно и носят недостоверный характер. Это может косвенно свидетельствовать об определенной генетической предрасположенности особей к расселению. Обнаружено, что довольно активно расселяются массовые фенотипы. Однако для некоторых немногочисленных у оседлых особей вариаций отмечено существенное увеличение частоты их встречаемости в группировках расселяющихся особей. Вероятно, именно эти признаки и фены могут быть рекомендованы в качестве маркировочных при изучении процессов дисперсии в популяциях рыжей полевки.
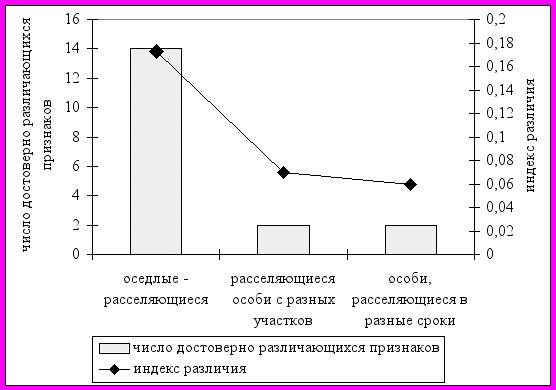 | Рис. 15. Число достоверно различающихся признаков (из 24) и величина индекса различия между сравниваемыми группировками оседлых (n=166) и расселяющихся (n=160) особей |
Темпы роста численности популяций грызунов во многом определяются степенью участия в репродукции сеголеток. Этот показатель очень изменчив, что связано с наличием двух альтернативных путей онтогенеза: однофазный рост и двухфазный рост (Шварц, 1980; Большаков и др., 1991; Оленев, 1991 и др.). В результате этого в популяции имеются разные функционально-физиологические группы особей (ФФГ). В изучаемой нами популяции рыжей полевки доля особей с однофазным ростом, размножающихся в год своего рождения (3 ФФГ), варьировала в широких пределах (0 - 43% - для самцов, 1 - 55% – для самок). У данной группы, прежде всего у самцов, обнаружен более высокий уровень флуктуирующей асимметрии качественных и количественных признаков. Это свидетельствует о снижении стабильности развития особей с однофазным ростом, что в определенной степени может уменьшать их жизнеспособность и сокращать продолжительность жизни. У перезимовавших животных (1ФФГ, бывшая 2ФФГ) доля асимметричных особей становится еще меньше, чем у их предшественников. Так, в случае с комплексным признаком непарная петля параконоида в суммарной группировке (1983-1987 гг.) перезимовавших особей, отмечено только 7,6% асимметричных особей. Тогда как во 2ФФГ суммарная доля асимметричных особей составляла 15,8%, а у особей с однофазным ростом (3ФФГ) – 19,7%. Эти результаты дополнительно подтверждают предположение о взаимосвязи стабильности онтогенеза и общей продолжительности жизни особей.