
Курс лекцій з основ біології для самостійної роботи студентів спеціальності 090804
| Вид материала | Курс лекцій |
- Тематичний план аудиторної та самостійної роботи 7 Перелік тем та питань самостійної, 209.58kb.
- Робоча навчальна програма для студентів спеціальності 060100 "Правознавство" Київ 2009, 1909.37kb.
- Робоча навчальна програма для студентів спеціальності 060100 "Правознавство" Затверджено, 742.86kb.
- Методичні рекомендації до практичних занять та самостійної роботи з "Основ екології", 726.43kb.
- Робоча навчальна програма для студентів спеціальності 0304 "Міжнародні відносини" напряму, 503.7kb.
- Робоча навчальна програма для студентів спеціальності 030301 журналістика Затверджено, 191.2kb.
- Робоча навчальна програма для студентів спеціальності 030300 Історія. Затверджено, 245.88kb.
- Робоча навчальна програма для студентів спеціальності 030303 видавнича справа та редагування, 220.01kb.
- Робоча навчальна програма для студентів спеціальності 030302 „Реклама І зв’язки з громадськістю, 446.01kb.
- Робоча навчальна програма для студентів спеціальності „Реклама та зв’язки із громадськістю, 261.8kb.
Вплив середовища визначається формулою С=100%-Н. Тоді С=100% – 65% = 35%. Отже, у випадку шизофренії переважає вплив спадковості, але суттєву роль відіграють і умови середовища.
Таблиця 4 – Конкордантність деяких ознак людини у однояйцевих (ОБ) і двояйцевих (ДБ) близнят, %
| Ознаки | Конкордантність (%) | |
| ОБ | ДБ | |
| Нормальні: | ||
| Групи крові (АВ0) | 100 | 46 |
| Колір очей | 99,5 | 28 |
| Колір волосся | 97 | 23 |
| Патологічні: | ||
| Клишоногість | 32 | 3 |
| Щілина губи | 33 | 5 |
| Шизофренія | 70 | 13 |
| Гіпертонія | 26,2 | 10 |
На підставі даних таблиці видно, що для багатьох захворювань поряд із спадковим компонентом значну роль відіграють умови середовища, при яких відбувається реалізація генотипу у фенотипі.
Труднощі близнюкового методу пов’язані, по-перше, з відносно низькою частотою народження близнюків у популяції (1:86 – 1:88), що ускладнює добір достатньої кількості пар з даною ознакою; по-друге, з ідентифікацією монозиготності близнюків, що має велике значення для достовірних результатів.
Метод дерматогліфіки
Дерматогліфіка (від грец. derma – шкіра, gliphe – ма-
лювати) – це визначення рельєфу шкіри на долонях (пальмоскопія), пальцях (дактилоскопія), підошвах (плантоскопія). На відміну від інших частин тіла тут є епідермальні виступи – гребені, які утворюють складні візерунки. Встановлено, що візерунки є індивідуальною характеристикою людини і не змінюються впродовж життя. Дерматогліфічні дослідження мають важливе значення у визначенні зиготності близнюків, у діагностиці багатьох спадкових захворювань, а також в окремих випадках спірного батьківства, у судовій медицині, у криміналістиці для ідентифікації особи.
 Дактилоскопія. Папілярні лінії на подушечках пальців вивчають на відбитках, які наносять на папір після змащування пальців друкарською фарбою. Детальне дослідження візерунків проводять за допомогою лупи. Папілярні лінії різних напрямків ніколи не перетинаються, але можуть у певних пунктах зближуватися, утворюючи трирадіуси, або дельти. Не дивлячись на індивідуальну неповторність візерунків, виділяють три їх основні типи: дуги А (англ. arch – дуга); петлі L (англ. lor – петля) і завиткові візерунки W (англ. whorl – завиток). Дугові візерунки зустрічаються дуже рідко (6%), у цьому Рисунок 12 – Пальцеві візерунки: 1-завиток, 2- петля,
Дактилоскопія. Папілярні лінії на подушечках пальців вивчають на відбитках, які наносять на папір після змащування пальців друкарською фарбою. Детальне дослідження візерунків проводять за допомогою лупи. Папілярні лінії різних напрямків ніколи не перетинаються, але можуть у певних пунктах зближуватися, утворюючи трирадіуси, або дельти. Не дивлячись на індивідуальну неповторність візерунків, виділяють три їх основні типи: дуги А (англ. arch – дуга); петлі L (англ. lor – петля) і завиткові візерунки W (англ. whorl – завиток). Дугові візерунки зустрічаються дуже рідко (6%), у цьому Рисунок 12 – Пальцеві візерунки: 1-завиток, 2- петля, візерунку є лише один 3- дуга
напрям папілярних
ліній. Петлеві візерунки найбільш поширені (близько 60%). Це замкнений з одного боку візерунок, у якому лінії, не доходять до протилежного краю. Завиткові візерунки займають середнє місце за поширеністю (34%). Вони мають вигляд концентричних кіл, овалів, спіралей. Завитки мають дві дельти. На пальцях ніг є також три типи візерунків, але у іншому співвідношенні (більший процент дуг). Тактильні візерунки на підошві у людини редуковані порівняно з мавпами і займають меншу площу.
Пальмоскопія. Рельєф долоні дуже складний, у ньому виділяють ряд полів, подушечок і долонних ліній. Центральну долонну ямку оточують шість підвищень – подушечок. Біля основи великого пальця – тенар, біля протилежного краю долоні – гіпотенар, навпроти міжпальцевих проміжків знаходяться міжпальцеві подушечки. Біля основи ІІ, ІІІ, ІV i V пальців знаходяться пальцеві трирадіуси - місця, у яких сходяться три напрямки папілярних ліній. Їх позначають латинськими літерами a, b, c, d. Поблизу браслетної складки, яка відділяє кисть від передпліччя, розміщується головний (осьовий) долонний трирадіус (t). Якщо провести лінії від трирадіусів a і d до t, то утворюється кут долоні аtd (рис. 13), у нормі він не перевищує 57°.
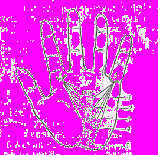
Рисунок 13 – Кут atd в нормі і при хромосомних
аномаліях:
1 – синдром Патау;
2 – синдром Дауна;
3 – синдром Шерешевського-Тернера;
4 – норма;
5 – синдром Клайнфельтера
На формування дерматогліфічних візерунків можуть впливати деякі пошкоджуючі фактори на ранніх стадіях ембріонального розвитку. Так, при внутрішньоутробній дії вірусу корової краснухи у дитини спостерігаються певні відхилення у візерунках, які подібні до таких при хворобі Дауна. Метод дерматогліфіки використовують при уточненні діагнозу хромосомних синдромів у людей зі змінами каріотипу. Менше показові дані дерматогліфічного аналізу при захворюваннях генної природи.
Біохімічний метод використовується для діагностики хвороб обміну речовин, причиною яких є зміни активності окремих ферментів. За допомогою біохімічних методів відкрито близько 5000 молекулярних хвороб, які є наслідком прояву мутантних генів. При різних типах захворювання можна або визначити сам аномальний білок-фермент, або проміжні продукти обміну. Дефекти ферментів установлюють шляхом визначення вмісту в крові і сечі продуктів метаболізму, що є результатом функціонування даного білка. Дефіцит кінцевого продукту, що супроводжується накопиченням проміжних і побічних речовин порушеного метаболізму, свідчить про дефіцит ферменту в організмі. Об’єктами біохімічної діагностики є сеча, піт, плазма і сироватка крові, формені елементи крові, культури клітин (фібробласти і лімфоцити). Програми первинної біохімічної діагностики спадкових хвороб можуть бути масовими і селективними. Відомі масові просіюючи програми для діагностики фенілкетонурії, спадкового гіпотиреозу та ін.
Біохімічна діагностика спадкових порушень обміну включає два етапи. На першому етапі вибирають ймовірні випадки захворювань, на другому більш точними і складними методами уточнюють діагноз захворювання. Для визначення в крові, сечі або амніотичній рідині проміжних, побічних і кінцевих продуктів обміну, крім якісних реакцій із специфічними реактивами на певні речовини, використовують хроматографічні методи дослідження амінокислот та інших органічних речовин.
Показаннями для застосування біохімічних методів діагностики новонароджених є такі симптоми: судоми, кома, блювота, гіпотонія, жовтяниця, специфічний запах сечі і поту, ацидоз, припинення росту. У дітей біохімічні методи використовуються у випадках підозри на спадкові хвороби обміну речовин (затримка фізичного і розумового розвитку, втрата набутих функцій, специфічна для будь-якої спадкової хвороби клінічна картина).
Порушення первинних продуктів генів виявляють за допомогою біохімічних методів, а локалізацію відповідних ушкоджень у спадковому матеріалі – за допомогою методів молекулярної генетики.
Цитогенетичний метод
Принципи цитогенетичних досліджень сформувалися протягом 20-30-х років ХХ століття на класичному об’єкті генетики – дрозофілі і деяких рослинах. Метод ґрунтується на мікроскопічному дослідженні хромосом. Нормальний каріотип людини становить 46 хромосом, із них 22 пари аутосом і 2 статеві хромосоми. До 1956 р. кількість хромосом у людини не була точно встановлена, це вдалося шведським вченим Д. Тийо і А. Левану. На той час у лабораторії успішно культивувалися клітини людини (клітини кісткового мозку, культури фібробластів або лейкоцитів периферичної крові, поділ яких стимулювали фітогемаглютиніном). За допомогою колхіцину зупиняли процес мітозу на стадії метафази, оскільки інактивувалися нитки веретена поділу; потім клітини оброблялися гіпотонічним розчином. У результаті набрякання і розривання клітинних мембран хромосоми виявлялися вільними і віддаленими одна від одної (метафазні пластинки). Це дало можливість підрахувати їх і проаналізувати. Найважливіше завдання полягає у вмінні розрізняти індивідуальні хромосоми у даній метафазній пластинці.
Цей метод застосовується для діагностики хромосомних захворювань. Хромосомні хвороби – це широка група спадкових патологічних станів, причиною якого є зміни кількості хромосом або порушення їх структури. У першому випадку у загальній генетиці використовується термін «геномні мутації», у іншому – «хромосомні мутації».
Існує багато аномалій каріотипу: аномалії аутосом та аномалії статевих хромосом.
Найвідомішими аномаліями аутосом є трисомія-21, трисомія-13 і трисомія-18.
Трисомія-21 (синдром Дауна) була описана англійським лікарем Л. Дауном у 1866 р. Причина патології – зайва 21-ша хромосома в каріотипі (трисомія 21-ї хромосоми) – каріотип 47, 21+. Хвороба Дауна – найбільш поширена з усіх хромосомних аномалій. Частота народжуваності дітей становить 1:500 – 1:700 новонароджених. Синдром Дауна характеризується такими ознаками: укорочені кінцівки, маленький череп, аномалії будови обличчя (плескаве, широке перенісся). Очні щілини вузькі, з косим розрізом, є складка верхньої повіки біля внутрішнього кута ока - епікант. Спостерігається різного ступеня розумова відсталість.
Трисомія-13 (синдром Патау). Каріотип – 47, 13+. Вперше ця аномалія каріотипу описана у 1960 р. К. Патау. При цій аномалії спостерігаються щілина м’якого і твердого піднебіння, незаростання губи, недорозвинення або відсутність очей, неправильно сформовані вуха, деформація кисті і стопи, трапляються полідактилія і синдактилія (зрощення пальців), численні порушення з боку внутрішніх органів – серця, нирок, травної системи. Частота народження дітей із синдромом Патау – 1:14 500 народжених живими.
Трисомія-18 (синдром Едвордса) описана у 1960 р. Каріотип 47, 18+. За даними різних авторів частота цієї хвороби коливається від 1:4500 до 1:6500. Смерть настає у 2-3-місячному віці. Зовнішній метод хворих настільки своєрідний, що дозволяє поставити діагноз до цитологічного аналізу. Череп незвичайної форми: вузький лоб і широка з виступом потилиця, дуже низько розташовані, деформовані вуха, постійна ознака – недорозвинення нижньої щелепи. Пальці рук широкі і короткі, характерна аномалія кисті – поперечна складка долоні.
Аномалія статевих хромосом. При дозріванні статевих клітин у людини може спостерігатися порушення розходження і статевих хромосом. Є дані, що це відбувається у 0,3% всіх гамет. Внаслідок цього у яйцеклітині замість однієї Х-хрмосоми можуть виявитися дві або не буде жодної. При заплідненні таких аномальних яйцеклітин нормальними сперматозоїдами будуть утворюватися зиготи, у яких змінена кількість статевих хромосом. Подібне явище може спостерігатися і при сперматогенезі. Аномалії кількості статевих хромосом бувають у вигляді моно- і полісомій.
Моносомія-Х (синдром Шерешевського–Тернера). Каріотип 45, Х0, фенотип жіночий. Це єдина сумісна з життям моносомія. Частота появи цієї аномалії 1:4000 – 1:5000, вона уперше була описана у 1925 р. ендокринологом Н. А. Шерешевським, а потім вивчалася Г. Тернером (1938). Проте причини цієї аномалії стали зрозумілими тільки в світлі досягнень цитогенетики. Основна патологічна ознака при цьому синдромі – недорозвинення яєчників. Своєрідна диспропорція тіла: більше розвинена верхня частина: широкі плечі і вузький таз, нижні кінцівки вкорочені. Зріст завжди нижчий від середньої норми (135-145 см). Характерні зовнішні ознаки: коротка шия зі складками шкіри, які йдуть від потилиці («шия сфінкса»), низький ріст волосся на потилиці, «антимонголоїдний» розріз очей (внутрішні кути очей розташовуються вище, ніж зовнішні).
Трисомія-Х. Каріотип 47, ХХХ. При такому комплексі народжується дівчинка, частота синдрому 1:1000 (0,1%). Фенотип різний. Більшість жінок має ряд певних відхилень у фізичному розвитку, порушення функції яєчників, передчасний клімакс, інтелектуальну неповноцінність, хоч у частини хворих ці ознаки і не проявляються.
Синдром Клайнфельтера спостерігається у осіб з чоловічим фенотипом. Каріотип 47, ХХY. Частота синдрому 1:1000 (0,1%). Характерною особливістю є недорозвинення сім’яників і відсутність сперматогенезу. Ця ендокринна недостатність визначає й інші ознаки фенотипу: розвивається астенічний, євнухоподібний тип будови тіла: вузькі плечі, широкий таз, відкладання жиру за жіночим типом, мало розвинена мускулатура, тобто спостерігається стирання статевих відмінностей, проявляються деякі конституційні ознаки протилежної статі.
Кожний синдром характеризується специфічним комплексом окремих аномалій. Точне підтвердження діагнозу дає аналіз каріотипу.
Метод гібридизації соматичних клітин
Соматичні клітини містять увесь обсяг генетичної інформації. Це дає можливість вивчати багато питань генетики людини, які неможливо досліджувати на цілому організмі. Соматичні клітини людини отримують із різних органів (шкіра, кістковий мозок, клітини крові, тканини ембріонів). Найчастіше використовують клітини сполучної тканини (фібробласти) і лімфоцити крові. Культивування клітин поза організмом дозволяє отримувати достатню кількість матеріалу для дослідження, який не завжди можна взяти у людини без шкоди для здоров’я.
У 1960 р. французький біолог Ж. Барський, вирощуючи поза організмом у тканинній культурі клітини двох ліній мишей, виявив, що деякі клітини за своїми морфологічними і біохімічними ознаками були проміжними між вихідними батьківськими клітинами. Ці клітини виявилися гібридними. Таке спонтанне злиття клітин у культурі тканини відбувається досить рідко. Згодом виявилося, що частота гібридизації соматичних клітин підвищується при введенні у культуру клітин РНК-вмісного вірусу парагрипу Сендай, який змінює властивості клітинних мембран і робить можливим злиття клітин. Вірус Сендай попередньо опромінювався ультрафіолетом. Такий інактивований вірус втрачав свої вірулентні властивості, але зберігав здатність впливати на злиття клітин. Під впливом такого вірусу у змішаній культурі двох типів клітин утворюються клітини, які містять у спільній цитоплазмі ядра обох батьківських клітин – гетерокаріони. Більшість гетерокаріонів гине, але ті, які містять тільки два ядра, часто продовжують свій розвиток, розмножуються поділом. Після мітозу і наступного поділу цитоплазми із двоядерного гетерокаріону утворюється дві одноядерні клітини, кожна з яких являє собою синкаріон – справжню гібридну клітину, яка має хромосоми обох батьківських клітин.
Гібридизація соматичних клітин проводяться у широких межах не тільки між різними видами, але і типами: людина миша, людина комар, муха курка тощо. Наприклад гібридні клітини людини і миші мають 43 пари хромосом: 23 – від людини і 20 від миші. Згодом при розмноженні цих клітин доля вихідних геномів різна. Відбувається поступова елімінація (зникнення) хромосом того організму, клітини якого мають повільніший темп розмноження. За допомогою цього методу проводиться картування хромосом у людини.
Використання методу гібридизації соматичних клітин дає можливість вивчати механізми первинної дії генів і взаємодію генів. Культури соматичних клітин використовуються для визначення мутагенної дії факторів навколишнього середовища.
Методи моделювання
Теоретичну основу біологічного моделювання у генетиці дає закон гомологічних рядів спадкової мінливості М. І. Вавілова, за яким генетично близькі види і роди характеризуються подібними рядами мутацій. Виходячи із цього закону можна передбачити, що у межах класу ссавців (і навіть за його межами) можна виявити багато мутацій, які викликають такі самі зміни фенотипу, як і у людини. Для моделювання певних спадкових аномалій людини підбирають і вивчають мутантні лінії тварин, які мають подібні порушення.
На сьогодні відомо близько трьохсот мутантних ліній кролів, пацюків і собак. Було описано і вивчено багато генетичних мутацій у тварин, які подібні до відповідних аномалій людини. Гемофілія А і В зустрічається у собак і зумовлена, як і у людини, рецесивними генами, локалізованими в Х-хромосомі. У ховрахів і пацюків виявлені патологічні мутації, які проявляються як гемофілія, цукровий діабет, ахондроплазія та деякі інші.
Мутантні лінії тварин точно не відтворюють спадкових хвороб людини. проте навіть часткове моделювання, тобто відтворення не всієї хвороби у цілому, а тільки патологічного процесу або навіть його фрагменту, дозволяє у ряді випадків виявити механізми первинного відхилення від норми. Поряд з біологічним моделюванням останнім часом використовуються методи математичного моделювання. Ці методи використовуються у популяційній генетиці (моделі популяцій).
У ряді випадків використовуються додаткові методи вивчення генетики людини: імунологічні, фізіологічні. Вивчаються особливості електроенцефалограм, швидкість утворення умовних рефлексів, реакції поведінки, використовуються психологічні тести.
Лекція 10 Генетика популяцій
Дарвінівське вчення утвердило в науці уявлення, що кожний вид – категорія історична, якісний етап еволюції. Кожний вид виник з іншого і існує, доки не зміняться умови. При нових умовах вид або загине, або змінюючись, дасть початок якісно новому чи новим видам.
Вид – сукупність особин, які схожі між собою за морфологічними і фізико-біохімічними ознаками, каріотипом, мають спільне походження, схрещуються між собою і дають плідне потомство та заселяють певну територію (ареал).
Біологічний вид, який складається з чисельних особин, що мають генетичні родинні зв’язки, але різняться за комбінаціями спадкових ознак, складає цілісну біологічну макросистему. Генофонд – сукупність усіх генів (генотипів) популяції, виду у певний період часу.
Особини будь-якого виду поширені у своєму ареалі не рівномірно, а окремими стійкими скупченнями – популяціями. Популяція – сукупність особин одного виду, що заселяють певну територію, схрещуються між собою і в певному ступені за певних причин ізольовані від інших аналогічних сукупностей.
Кожна популяція має визначений ареал, віковий і статевий склад; численність особин у популяції може коливатися від кількох сотень до кількох тисяч. Чим менше популяція, тим більша загроза її вимирання або загибелі від будь-яких випадкових причин.
Не тільки види тварин і рослин складаються з популяцій. Людською популяцією називають групу людей, які займають одну територію і вільно вступають у шлюб. Факторами, що обмежують людей у взятті шлюбу, можуть бути географічні, соціальні, релігійні тощо. Великі популяції людей складаються не з однієї, а із кількох антропологічних груп, які відрізняються за походженням, і розселені на великі території.
Малі популяції, чисельність яких не перевищує 1500 – 4000 осіб, називають демами. Вони характеризуються великою частотою споріднених шлюбів (80 – 90%). Ще менші людські популяції з чисельністю не більше 1500 осіб називають ізолятами, у яких споріднені шлюби становлять більше 90%.
Генетичні процеси у популяціях. У ряді випадків до складу популяції одночасно можуть входити особини як з домінантними, так і з рецесивними ознаками, які не знаходяться під контролем природного добору. Виникає питання: чому рецесивний алель не витісняється домінантним? Це питання чисто математично для ідеальної популяції розв’язали у 1908 р. незалежно один від одного математик Дж. Харді і лікар В. Вайнберг. Виявлена ними закономірність називається законом Харді–Вайнберга.
Ідеальна популяція характеризується такими особливостями: безмірністю, вільним схрещуванням (панміксією), відсутністю мутацій за даним геном, відсутністю міграцій у популяції, відсутністю добору (за ознакою, яка кодується даним геном). У ідеальній популяції співвідношення генотипів домінантних гомозигот (АА), гетерозигот (Аа) і рецесивних гомозигот (аа) залишаються постійним.
Якщо частоту гена А позначити через р, а частоту гена а через q, то q буде дорівнювати 1–р. У F2 і наступних поколіннях частота генотипів АА, Аа і аа буде визначатися за формулою біному Ньютона
(р+ q)2 = р2 + 2рq + q2,
Де р – частота домінантного алеля А; q – частота рецесивного алеля а; р2 – частота домінантних гомозигот АА; 2рq – частота гетерозигот Аа; q2 – частота рецесивних гомозигот аа. Величини р2, 2рq, і q2 залишаються постійними. Цим пояснюється той факт, що за наявності умов, які властиві ідеальній популяції, особини з рецесивними ознаками зберігаються поряд з особинами, які несуть домінантні ознаки.
У реально існуючих популяціях наведені вище умови нездійснені: реальні популяції мають обмежену чисельність, панміксія ніколи не буває абсолютною, відбуваються міграції особин і мутаційний процес. Проте це не зменшує значення закону Харді-Вайнберга. Він встановлює цілком визначені співвідношення між алелями у популяції. Використання формул закону Харді-Вайнберга дозволяє розрахувати генетичний склад популяції у даний час і визначити тенденції до його змін.
Використовуючи закон Харді – Вайнберга, можна вирахувати насиченість популяції певними генами, розрахувати частоти гетерозиготного генотипу у людей.
Розглянемо використовування його на конкретному прикладі. У населеному пункті при обстеженні на резус-фактор виявилося 16% особин з резус-негативним фактором і 84% – з резус-позитивним. Відомо, що позитивний резус-фактор успадковується практично моногенно, аутосомно, за домінантним типом. Якщо ген резус-фактора позначити С, то носії Rh+ будуть мати генотип СС і Сс. Але яка частина із них гомо- і гетерозиготна (тобто, яка концентрація рецесивного алеля)? Використаємо формулу
(р+ q)2 = р2 + 2рq + q2 = 1 (або 100%).
Гомозиготи за рецесивним алелем q2 відомі, вони становлять 16%. Отже, q2 =0,16, звідси q = 0,4 (або 40%), тобто із загальної кількості генів популяції, які визначають резус-належність, 40% рецесивних.
Яка ж частка домінантного алеля? Оскільки р+q=1, а q = 0,4, то р = 0,6, тобто у популяції 60% домінантних алелей.
Для того, щоб знайти процент поєднань у популяції генів СС і Сс, вираховуємо: р2 = (0,6)2 = 0,36, тобто 36% мають генотип СС, 2рq = 2 (0,6) х (0,4) = 0,48, тобто 48% мають генотип Сс. Отже, у досліджуваній групі людей, з 84% особин з резус-позитивною кров’ю було: 36% з генотипом СС і 48% з генотипом Сс, а 16%, які мали генотип сс, були резус-негативними.
При медико-генетичних дослідженнях популяцій подібні розрахунки досить широко поширені. Але в тих випадках, коли популяції обмежені за чисельністю, закон Харді–Вайнберга не діє, бо базується на статистичних заномірностях, які не виконуються у разі малих чисел. Основна маса людства складається із великих популяцій, у яких за законом Харді–Вайнберга підтримується рівновага генетичного складу. Проте ця рівновага постійно порушується мутаційним процесом, міграціями, дрейфом геном і іншими факторами.
Популяція як елементарна еволюційна структура. У ареалі будь-якого виду особини поширені нерівномірно. Ділянки великої концентрації особин чергуються з просторами, де їх мало або зовсім немає. У результаті виникають більш або менш ізольовані популяції, у яких систематично відбувається випадкове вільне схрещування (панміксія). Схрещування (тобто обмін генами) з представниками інших популяцій, якщо і відбувається, то значно рідше і нерегулярно. Завдяки ізоляції в кожній популяції створюється властивий для неї генофонд, який відрізняється від інших популяцій. Саме популяцію і необхідно визнати елементарною одиницею еволюційного процесу.
Поняття про мікроеволюцію
Мікроеволюцією називають еволюційний процес, який відбувається всередині виду, веде його до диференціації і може завершитися утворенням нового виду. Але найчастіше результатом мікроеволюції є утворення генетичного поліморфізму (перевага гетерозигот в популяції над гомозиготами, при якому обидва алелі зберігаються у популяції з проміжною частотою й популяція знаходиться в стані рівноваги). Мікроеволюція здійснюється у короткий історичний час і доступна для безпосереднього вивчення. Вчення про мікроеволюцію складає основу синтетичної теорії еволюції. Це вчення ґрунтується на точних методах дослідження еволюційного процесу в популяціях (генетичні, екологічні, математичні, експеримент з моделюванням). За синтетичною теорією еволюції, популяція – елементарна одиниця еволюції. Елементарним еволюційним матеріалом є мутація. Видоутворення починається з елементарного еволюційного явища – стійкості і напрямленої зміни генетичного складу (генетичної конституції або генофонду) популяції.
Властивості популяції: величина, ступінь ізольованості від суміжних популяцій, периферичне або центральне розташування популяції в межах ареалу виду, статевий і віковий склад популяції, внутрішньопопуляційний поліморфізм.
Явища і процеси, які змінюють генофонд популяції, називаються елементарними еволюційними факторами. Розрізняють ненапрямлені і напрямлені еволюційні фактори. До ненапрямлених елементарних еволюційних факторів належать: мутаційний процес, популяційні хвилі, ізоляція, міграція, дрейф генів. Напрямленим елементарним еволюційним фактором є єдиний фактор – природний добір. Він спрямовує еволюційний процес у бік вироблення нових і вдосконалення існуючих пристосувань.
Проблема генетичного вантажу у людини має велике значення для медицини. Для медико-генетичного консультування важливо мати уявлення про насиченість генами спадкових хвороб населення на тих чи інших територіях.
Мутації як елементарний еволюційний матеріал.
Гени, які у цілому константні, періодично змінюються шляхом мутацій. Еволюція організмів пов’язана із заміною одних генотипів іншими. Більшість мутацій шкідливі: вони можуть бути летальними і напівлетальними, викликати безпліддя або знижувати життєві функції. Деякі із мутацій за тих умов, що існує дана популяція організмів, виявляються більш-менш нейтральними. Зрештою, невелика частина мутацій в якійсь мірі корисна для життя організму, для існування виду. Увесь поліморфізм людства – результат мутаційних змін. Вважається, що у людини на один гаплоїдний набір за покоління виникає від 1 до 10 нових мутацій, а на диплоїдний набір їх у два рази більше. Крім того, людство несе у собі генетичний вантаж раніше виниклих мутацій, серед яких немало рецесивних, летальних, напівлегальних, що спричиняють ряд спадкових хвороб.
Другим фактором еволюції є популяційні хвилі, тобто зміни чисельності особин у популяції. Встановлено, що немає жодної популяції де чисельність особин є постійною. У деяких видів (комахи, риби тощо) чисельність популяцій може змінюватися у десятки і сотні разів. Ці коливання С. С. Четверіков образно називав хвилями життя. Зростання або зменшення чисельності особин у популяціях може бути позасезонним, так і не сезонним, повторюватися через різні проміжки часу і бути зв’язаним, наприклад, із змінами кількості кормів.
У малих популяціях частота алелів може змінюватися без участі природного добору – внаслідок випадкових процесів. Зміни частот алелів у малих популяціях американський генетик Стьюел Райт назвав дрейфом генів. Інтенсивність дрейфу генів залежить від розміру популяції. Встановлено, що у невеликих популяціях гетерозиготні особини рано чи пізно зникають і зростає генетична однорідність. Вся популяція стає гомозиготною. Прикладом наслідку дрейфу генів є кількість резус-негативних людей у Європі (14%) і у Японії (1%). Якщо у генофонді популяції є летальні гени, це може мати фатальні наслідки – настане вимирання всієї популяції.
З дрейфом генів зв’язаний принцип засновника. Терміном «принцип засновника» Е. Майр позначив виникнення нової популяції від кількох засновників (навіть від однієї заплідненої самки в екстремальному випадку). Більшість самців птахів, успішно вселених у Північну Америку, Австралію і Нову Зеландію, а також майже всі завезені комахи були нащадками лише кількох особин і несли лише декілька генів з усього генофонду вихідної популяції.
У зміні генофонду людських популяцій не останню роль відіграють міграції. З ними пов’язані порушення попередніх обмежень шлюбів, поява змішаних шлюбів. Міграції ведуть до змін складу генів як у популяціях, із яких населення емігрувало, так і у тих, куди емігрувало.
Завдяки ізоляції і дрейфу генів у популяціях зростає гомозиготність, відмічаються підвищена загибель плодів, мертвонародження, природжені аномалії і спадкові хвороби.
Ізоляція – відмежування, виключення вільного схрещування між особинами, внаслідок чого відбувається обмежена чи повна відсутність обміну генами між групами особин, популяціями чи видами.
Причини ізоляції у людських популяціях можуть бути різними:
- географічні (острови, гірські поселення);
- національні;
- расові;
- релігійні;
- соціальні.
За природою факторів серед усіх живих істот розрізняють такі форми ізоляції:
- географічна (просторова) – роз’єднання популяцій простором:
- фізико-географічним бар’єром (гори, ріки, моря, пустелі);
- відстанню (віддалені селища);
- екологічна – пристосування організмів до різних умов середовища:
- біотопічна – близькі види існують на одній території, але для життя займають різні місця (біотопи), не зустрічаються між собою і не схрещуються;
- сезонна – особини існують на одній території, зустрічаються між собою, але розмножуються в різні календарні терміни (так, в озері Севан (Вірменія) існують 5 рас форелі, які розмножуються у різні місяці на різних глибинах);
- біологічна ізоляція:
- морфофізіологічна – досягається відмінностями в будові органів розмноження, що унеможливлює процес копуляції між різними видами;
- етіологічна (поведінкова) – пов’язана з особливостями шлюбної поведінки тварин: ритуалами залицяння, звуковими, зоровими і хімічними подразниками, на які реагують особини того самого виду і не реагують особини інших видів (комахи, птахи, ссавці);
- генетична (репродуктивна) – несумісність гамет (неможливе запліднення), нежиттєздатність зигот, гібридів F1, стерильність гібридів (порушений гаметогенез). Таким чином, ізолюючі механізми являють собою серію бар’єрів. Якщо був подоланий один бар’єр, на шляху до панміксії стає наступний.
Усі елементарні еволюційні фактори взаємозв’язані між собою під контролем природного добору: мутаційний процес, популяційні хвилі є факторами-постачальниками еволюційного матеріалу; ізоляція – фактор, який підсилює генетичні відмінності між групами особин. Механізм цих факторів різний, але спільними є ненаправленість і випадковість їх дії. Спрямованості еволюційному процесу надає природний добір. Добір зажди має направлений характер. Добір, як і еволюція в цілому, має пристосувальний характер. Він приводить генофонди у відповідність з критерієм пристосованості. Результатом творчої ролі добору слугує процес органічної еволюції, який в цілому відбувається шляхом прогресивного морфофізіологічного ускладнення, а на окремих етапах – шляхом спеціалізації.
Лекція 11 Організм та середовище. Математичне моделювання в екології. Екосистеми
Екологія – наука, що вивчає зв’язки організмів між собою та з навколишнім середовищем. Порушення екологічної рівноваги може спричинити екологічну небезпеку, екологічну кризу та екологічну катастрофу. Перед людством постають різноманітні екологічні проблеми:
- забруднення природного середовища відходами промислового та сільськогосподарського виробництва;
- потепління клімату;
- забруднення атмосфери кислотними опадами;
- запустелювання великих територій;
- зниження біологічного різномаїття;
- вимирання видів;
- втрата цілих екосистем.
Крім зазначених проблем, в екології людини виникають питання заповнення екологічних ніш небажаними організмами (паразити, шкідники, збудники нових захворювань, такі як вірус пташиного грипу, пріони тощо); перенаселення Землі, демографічні катаклізми; погіршення середовища у міській і сільській місцевостях.
Вперше визначив екологію як «науку, що вивчає взаємовідносини тварин з органічною та неорганічною природою» у 1870 р. німецький вчений Е. Геккель у своїй праці «Загальна морфологія організмів». У 1895 р. датський ботанік Е. Вармінг застосував цей термін до представників рослинного світу. І лише у ХХ ст. екологія сформувалася як самостійна біологічна наука зі своїми методами і предметом вивчення.
Об’єктами досліджень в екології можуть бути окремі організми, популяції, угрупування, екосистеми та вся біота нашої планети. Тому в екології виділяють три рівні вивчення: 1) популяційно-видовий; 2) екосистемний; 3) еволюційно-історичний.
Популяційно-видовий рівень передбачає вивчення індивідуальних реакцій окремих організмів, популяцій або виду в цілому на дію чинників навколишнього середовища.
Екосистемний рівень досліджень передбачає вивчення процесів, спричинених взаємним впливом організмів чи популяцій різних видів. На цьому рівні виконуються надзвичайно складні й трудомісткі дослідження, спрямовані на з’ясування закономірностей, які визначають існування у просторі та часі зв’язків між різними видами автотрофів і гетеротрофів.
Еволюційно-історичний рівень досліджень передбачає встановлення стратегічних можливостей розвитку об’єктів досліджень під впливом глобальних (історичного масштабу) коливань параметрів навколишнього середовища, таких як вікові коливання погодних умов, зміни ґрунтів або рельєфу тощо.
У сучасній екології застосовуються такі методи досліджень:
- емпіричні;
- експериментальні;
- моделювання.
До емпіричних методів належать численні способи реєстрації прямих спостережень, вимірювання за допомогою будь-яких технічних засобів. Але не завжди вистачає лише спостережень для проведення зваженого аналізу даних, тому часто застосовуються експерименти.
Експерименти здійснюються на основі теорій, які вимагають інтерпретації результатів досліджень. До експериментальних належать методи, що передбачають контроль та керування умовами, в яких перебуває об’єкт дослідження. Основним у виконанні експериментів є перевірка гіпотез, припущень, котрі мають принципове практичне значення.
До методів моделювання в екології належать: предметне моделювання, при якому модель певним чином відтворює параметри оригіналів, і знаково-математичне моделювання, при якому об’єкт відтворюється віртуально (комп’ютерне моделювання) або у вигляді графіків, гістограм, схем тощо.
З метою передбачення змін у природних екосистемах, зумовлених природними процесами або діяльністю людини, здійснюють екологічне прогнозування. Виділяють такі рівні прогнозування:
- локальне (поширюється на невелику територію);
- національне (поширюється на окрему державу);
- регіональне (поширюється на декілька країн, материк, океан тощо);
- глобальне.
Екологічний прогноз передбачає можливі зміни в екосистемах, основуючись на об’єктивних наукових даних про закономірності функціонування живих систем та їхні реакції на вплив певних чинників.
Навколишнє середовище – це сукупність чинників, які здатні прямо чи опосередковано впливати на життєдіяльність організмів. Часто їх називають екологічними чинниками. Виділяють чотири основні типи середовища: водне, повітряне, ґрунтове і внутрішнє середовище організмів.
За ступенем впливу на організми зовнішні чинники поділяються на:
- життєво необхідні (світло, вода, мінеральні солі, кисень);
- факультативні активні (дим, радіаційне випромінювання);
- факультативні нейтральні (інертні гази).
За походженням виділяють:
- біотичні;
- абіотичні;
- антропогенні.
За характером впливу чинники поділяються на
- прямі (безпосередньо впливають на конкретні показники стану організму чи популяції);
- непрямі (впливають опосередковано).
Абіотичні чинники поділяються на елементарні (температура, вода, атмосферний тиск, повітря, електромагнітне поле) та комплексні (хімічний склад субстрату (розчину, газу), агрегатний стан субстрату, сонячне світло).
Біотичні чинники являють собою дію живих організмів один на одного. Вони поділяються на фітогенні, зоогенні, мікробіогенні та мікогенні.
Антропогенними чинниками вважають будь-які впливи, які спричинені діяльністю людини в природних екосистемах. Відповідно до характеру діяльності антропогенні чинники бувають: сільськогосподарські, транспортні, військові, промислові та природоохоронні.
Кожен окремо взятий чинник може бути охарактеризований певними параметрами, значення яких можна зареєструвати візуально або інструментально. Параметри кожного чинника можуть змінюватися в просторі і часі. Такі коливання параметрів позначаються на життєдіяльності організмів.
Залежно від діапазону, в якому організм чи популяція здатні сприймати коливання параметрів окремих чинників, виділяють еврибіонтні та стенобіонтні системи. Еврибіонтами називають організми (системи), здатні існувати і
відтворюватися в широких межах дії чинника. Наприклад, рудий тарган (прусак) може існувати і розмножуватися в умовах екстремальних температур, освітленості, хімічного складу атмосфери тощо. Стенобіонтами є організми (системи), які можуть забезпечувати повноцінне існування лише в умовах вузького діапазону дії окремих чинників. Так, усі ендопаразити є стенобіонтами відносно температури (личинка людської аскариди), хімічного складу рідини, що оточує паразита (печінковий сисун), або інших чинників.