
Курс лекцій з основ біології для самостійної роботи студентів спеціальності 090804
| Вид материала | Курс лекцій |
- Тематичний план аудиторної та самостійної роботи 7 Перелік тем та питань самостійної, 209.58kb.
- Робоча навчальна програма для студентів спеціальності 060100 "Правознавство" Київ 2009, 1909.37kb.
- Робоча навчальна програма для студентів спеціальності 060100 "Правознавство" Затверджено, 742.86kb.
- Методичні рекомендації до практичних занять та самостійної роботи з "Основ екології", 726.43kb.
- Робоча навчальна програма для студентів спеціальності 0304 "Міжнародні відносини" напряму, 503.7kb.
- Робоча навчальна програма для студентів спеціальності 030301 журналістика Затверджено, 191.2kb.
- Робоча навчальна програма для студентів спеціальності 030300 Історія. Затверджено, 245.88kb.
- Робоча навчальна програма для студентів спеціальності 030303 видавнича справа та редагування, 220.01kb.
- Робоча навчальна програма для студентів спеціальності 030302 „Реклама І зв’язки з громадськістю, 446.01kb.
- Робоча навчальна програма для студентів спеціальності „Реклама та зв’язки із громадськістю, 261.8kb.
Неповне домінування – така форма взаємодії, коли у гетерозиготного організму (Аа) домінантний ген (А) не повністю пригнічує рецесивний ген (а), внаслідок чого проявляється проміжна між батьківськими ознака:
А – червоні квіти P: ♀АА X ♂аа
а – білі квіти F1: Аа (рожеві)
Наддомінування – коли ген у гетерозиготному стані виявляється сильнішим, ніж у гомозиготному.
АА – норм. тривалість життя;
Аа – подовжена тривалість;
аа – летальний наслідок (у дрозофіл).
Гетерозис у рослин і тварин - "гібридна сила" – використовується в селекції (життєздатність, швидкий ріст, вага, плодючість).
Кодомінування – одночасний прояв алелей у гетерозиготі за відсутності тенденцій до будь-якого домінування одного над іншим. Класичним прикладом є система груп крові АВ0, коли еритроцити людини з ІV групою крові несуть на поверхні антигени, що контролюються обома алелями.
Системи груп крові
Властивості груп крові за системою АВ0
Людина за цією системою має чотири фенотипи: група І (0), група ІІ (А), група ІІІ (В), група ІV (АВ). Кожний з цих фенотипів відрізняється специфічними білками-антигенами, які містяться в еритроцитах, і антитілами, що зосереджуються у сироватці крові (табл. 2). Антиген – речовина, переважно білкової природи, яка при введенні в організм викликає утворення антитіл і здатна реагувати з антитілами в різних реакціях. Антитіло – захисний білок (субстанція), який продукується в організмі імунною системою ссавців (В-лімфоцитами) при потраплянні в організм антигену. Антитіла специфічним чином зв’язують антигени й нейтралізують їх шкідливу дію. Фенотип І (0) групи зумовлений відсутністю у еритроцитах антигенів А і В і наявністю у сироватці крові антитіл α і β. Фенотип ІІ (А) характеризується наявністю в еритроцитах антигена А і антитіл β у сироватці. Фенотип ІІІ (В) пов'язаний з наявністю у еритроцитах антигена В, а у сироватці крові – антитіл α. Фенотип ІV (АВ) зумовлений наявністю в еритроцитах антигенів А і В та відсутністю у сироватці крові антитіл α і β.
Таблиця 2 Властивості груп крові за системою АВ0
| Фенотип | Антиген | Антитіло | Генотип |
| І (0) | – | , | І0 І0 |
| ІІ (А) | А | | ІА ІА, ІА І0 |
| ІІІ (В) | В | | ІВ ІВ, ІВ І0 |
| ІV (АВ) | А, В | – | ІА ІВ |
Кодомінування має місце і при успадкуванні групи крові за системою MN, відкритою у 1927 р. Ця система визначається двома алелями: ІМ і ІN. Обидва алелі кодомінантні, тому існують люди з генотипом ІМІМ (у фенотипі вони мають фактор М), ININ (у фенотипі вони мають фактор N), IМIN (у фенотипі у них обидва фактори М і N). У сироватці крові людей з тим чи іншим фенотипом за цією системою груп крові немає антитіл до відповідних антигенів, як це має місце у системі АВ0. Тому звичайно при переливанні крові ця система може не враховуватися. Серед європейців генотип ІМІМ зустрічається приблизно у 36%, ININ – у 16% і IМIN – у 48%.
Важливою є резус-система груп крові.
У 1940 р. із еритроцитів макак-резус було виділено антиген, який назвали резус-фактором (Rh-фактор). Згодом він був знайдений і у людей:. близько 85% європейців мають його, тобто є резус-позитивними (Rh+), а у 15% резус-негативних (rh-) він відсутній.
У нормі в осіб з резус-негативною кров’ю не виробляються антитіла до резус-фактора, але вони почнуть вироблятися у результаті переливання резус-позитивної крові як захисна реакція проти чужорідного гена.
Резус–позитивний фактор зумовлений домінантними генами. При шлюбі жінки з резус-негативною кров’ю і чоловіка з наявністю резус-фактора за умови гомозиготності батька всі діти будуть резус-позитивними, а при гетерозиготності буде спостерігатися розщеплення у відношенні 1:1.
Якщо у жінки з резус-негативною кров’ю дитина, що народиться, успадковує резус-фактор, перша вагітність може завершитись цілком нормально. Але при цьому у кров’яному руслі матері утворюються антитіла до Rh+-фактора. При наступній вагітності ці антитіла проникають у кров плода і викликають руйнування еритроцитів, які мають антиген Rh. З кожною наступною вагітністю, несумісною за антигенами, кількість антитіл до Rh-фактора у тілі матері зростає і обумовлює резус-конфлікт.
Резус-конфлікт (Rh-несумісність) – несумісність Rh-антигенів rh--матері і Rh+-плода, або Rh+-донора і rh--реципієнта, що призводить до розвитку важких форм гемолітичної хвороби новонароджених (гемолітична жовтяниця викликана руйнуванням еритроцитів) або трансфузійних ускладнень (через 1-2 години після гемотрансфузії). Гемотрансфузія – процес переливання крові від донора до реципієнта. Переливання несумісної за цим фактором крові дівчатам і жінкам зовсім недопустиме, бо може викликати безпліддя. У реципієнта уражаються нирки і печінка, розвиваються жовтяниця й анемія.
Взаємодія неалельних генів
Неалельні гени – гени, що розміщені в різних локусах однієї хромосоми або в не гомологічних хромосомах.
Розрізняють такі види взаємодії неалельних генів: комплементарність, епістаз, полімерія.
Комплементарність – вид взаємодії неалельних генів, коли один домінантний ген доповнює дію іншого неалельного домінантного гена, і вони разом визначають нову ознаку, яка була відсутня в батьків. Цей тип у природі дуже поширений.
У запашного горошку колір віночка квітки зумовлений наявністю двох домінантних генів (А і В), за відсутності одного з них – квітки білі. Тому при схрещуванні рослин з генотипами ААbb і aaBB, які мають білі віночки, у першому поколінні рослини мають забарвлення, а у другому поколінні розщеплення відбувається у співвідношенні 9 забарвлених до 7 незабарвлених (3А_bb, 3 ааВ_, 1 ааbb). Таке розщеплення пояснюється тим, що синтез пігменту антоціану з речовини-попередника проходить через стадію проміжного з’єднання, і обидва біохімічних процеси обумовлюються ферментами, що кодуються неалельними генами А і В. Тільки гетерозигота А_В_ здатна синтезувати пігмент. Самі по собі гени А і В не мають самостійного зовнішнього прояву.
У мишей сірий колір також зумовлений двома домінантними генами: з них ген А необхідний для утворення пігменту, тому рецесивні гомозиготи (аа) не мають пігменту (альбіноси). Ген В зумовлює нерівномірний розподіл пігменту, а рецесивний його алель (b) кодує рівномірний розподіл пігменту по всьому волоссі, і рецесивні гомозиготи (bb) за присутності гена А мають чорний колір. Домінантний алель (В) кодує відкладення пігменту зонами, біля верхівки і основи волосини, і його носії за присутності гена А мають сірий колір. При схрещуванні дигетерозигот (АаВb) спостерігається розщеплення у співвідношенні 9:3:4. Два домінантні гени у генотипі А_В_ дають сірий колір (9 частин), носії домінантного гена А і рецесивного b – чорні (3А_bb), за відсутності гена А (3ааВ_ і ааbb) – білі.
Комплементарну взаємодію генів у людини можна проілюструвати на таких прикладах. Нормальний слух зумовлюється двома домінантними неалельними генами D і F, з яких один визначає розвиток завитка, а інший – слухового нерва. Домінантні гомозиготи і гетерозиготи за обома генами мають нормальний слух, рецесивні гомозиготи за одним з цих генів або за двома генами – глухі.
Епістаз – (від грецької "зупинка, перешкода") – така взаємодія неалельних генів, за якої один ген пригнічує дію іншого неалельного гена. Пригнічення можуть викликати як домінантні так і рецесивні гени, і залежно від цього розрізняють епістаз домінантний і рецесивний.
Ген, який виконує пригнічуючу функцію, називається супрессором або інгібітором.
Наприклад, у курей домінантний алель гена С зумо-
влює розвиток пігменту, але домінантний алель іншого гена І є його супресором, «пригнічувачем». Тому навіть ті кури, які мають домінантний алель гена кольору, за присутності супресора виявляються білими. Особини, які мають у генотипі гени І_С_ – білі, а з генотипом ііСС і ІІСс – пігментовані. Таким чином, білий колір курей може бути зумовлений як відсутністю домінантного алеля гена пігментації, так і наявністю домінантного алеля гена, який викликає явище епістазу гена пігментації. Отже, якщо схрестити двох білих птахів з генотипами ІІСС і іісс, всі особини першого покоління виявляться білими ІіСс, але у другому поколінні (F2) при спорідненому схрещуванні особин першого покоління відбудеться розщеплення за фенотипом у співвідношенні 13:3. 3/16 птахів будуть пігментовані (ііСС і ііСс), бо у них ген-супресор знаходиться в рецесивному стані і є домінантний ген пігментації.
Полімерія - така взаємодія неалельних генів, при якій декілька генів зумовлюють розвиток однієї ознаки, причому вони не проявляють однакову дію. Полімерні гени прийнято позначати однією літерою латинського алфавіту з цифровим індексом, наприклад: А1А1 і а1а1; А2А2 і а2а2 і т.д. Уперше одночасні фактори були виявлені шведським генетиком Нільсоном-Еле (1908 р.) при вивченні успадкування кольору насіння у пшениці. Було встановлено, що ця ознака залежить від двох полімерних генів, тому при схрещуванні домінантних і рецесивних дигомозигот – забарвленої (А1А1 А2А2) з безколірною (а1а1а2а2) в F1 всі рослини дають забарвлене насіння, хоча воно помітно світліше, ніж батьківські екземпляри, які мають червоне насіння. При схрещуванні особин першого покоління в F2 виявляється розщеплення за фенотипом у співвідношенні 15:1, бо безколірними є лише рецесивні дигомозиготи (а1а1а2а2). У пігментованих екземплярів інтенсивність кольору дуже варіює залежно від кількості отриманих ними домінантних алелів: максимальна у домінантних дигомозигот (А1А1 А2А2) і мінімальна у носіїв одного з домінантних алелів (А1а1а2а2 і а1а1А2а2).
Важлива особливість полімерії – сумація (адитивність) дії неалельних генів на розвиток кількісних ознак. Якщо при моногенному успадкуванні ознаки можливі три варіанти «доз» гена у генотипі АА, Аа, аа, то при полігенному кількість їх зростає до чотирьох і більше. Сумація «доз» полімерних генів забезпечує існування безперервних рядів кількісних змін. Так, наприклад, у людини зріст визначається двома не алельними генами (домінантні гени А1 і А2 визначають в генотипі низький зріст, а рецесивні а1 і а2 – високий зріст). Залежно від кількості домінантних і рецесивних генів в генотипі людини ділять на дуже низький зріст – А1А1А2А2, низький зріст – А1А1А2а2, або А1а1А2А2, середній зріст – А1А1а2а2, а1а1А2А2 чи А1а1А2а2, високий зріст – А1а1а2а2 або а1а1А2а2, та дуже високий а1а1а2а2. У шлюбі дуже низької людини з дуже високою нащадки в F1 будуть всі середнього зросту:
Р: ♀ А1А1А2А2 X ♂ а1а1а2а2
F1: А1а1А2а2.
Якщо ж батьки будуть середнього зросту за фенотипом і дигетерозиготами за генотипом, то розщеплення у F2 буде спостерігатися таким:
Р: ♀ А1а1А2а2 X ♂ А1а1А2а2
F2
| Гамети ♂ ♀ | А1А2 | А1а2 | a1А2 | а1а2 |
| А1А2 | А1А1А2А2 | А1А1А2а2 | А1а1А2А2 | А1а1А2а2 |
| А1а2 | А1А1А2а2 | А1А1а2а2 | А1а1А2а2 | А1а1а2а2 |
| a1А2 | А1а1А2А2 | А1а1А2а2 | а1а1А2А2 | а1а1А2а2 |
| а1а2 | А1а1А2а2 | А1а1а2а2 | а1а1А2а2 | а1а1а2а2 |
Народження дуже низьких дітей буде дорівнювати 1/16, низького зросту 4/16, середнього 6/16, високого 4/16 і дітей дуже високого зросту 1/16. Дане розщеплення за фенотипом можна побачити в даній решітці Пенета.
Біологічне значення полімерії полягає ще у тому, що ознаки, які кодуються цими генами, більш стабільні, ніж ті, що кодуються одним геном. Організм без полімерних ознак був би дуже нестійким: будь-яка мутація, або рекомбінація приводила б до різкої мінливості, а це у більшості випадків невигідно.
У тварин і рослин є багато полігенних ознак, серед них і цінні для господарства: інтенсивність росту, скороспілість, яйценосність, кількість молока, його жирність, вміст цукристих речовин і вітамінів тощо.
Багато морфологічних, фізіологічних і патологічних особливостей людини визначається полімерними генами: зріст, маса тіла, величина артеріального тиску. Розвиток таких ознак у людини підпорядковується загальним законам полігенного успадкування і дуже залежить від впливу умов середовища.
При некумулятивній полімерії ознака розвивається повністю при наявності хоча б одного з полімерних генів, а при кумулятивній полімерії ступінь розвитку ознаки залежить від кількості даних алелів у генотипі, оскільки кожен із генів сам по собі дає слабку дію, а дія декількох генів сумується. Кумулятивна полімерія характерна для кількісних ознак.
Якісна ознака – це одна з альтернативних ознак, що чітко відрізняється між собою (наприклад, праворукість – ліворукість).
Кількісна ознака – ознака, ступінь вираження якої варіює, потребує змін, підрахунку. Кількісні ознаки характеризуються безперервною мінливістю. Організми за кількісними ознаками не можуть бути розмежовані на чіткі фенотипові класи, оскільки такий поділ завжди носить умовний характер із-за умовності меж (наприклад: зріст – високий, середній, низький; вага організму, колір шкіри, кров’яний тиск, об’єм легень, жирність молока тощо).
У більшості випадків окремі гени самостійно не визначають ознак. У явищах комплементарності, епістазу і полімерії фенотипічно проявляється молекулярна взаємодія генів.
Лекція 6 Мінливість ─ властивість живого. Генотип і фенотип. Проблеми мутагенезу
Мінливістю називають відмінності між особинами одного виду – предками і нащадками, які виникають внаслідок змін спадкового матеріалу або впливу умов зовнішнього середовища. Мінливість, як і спадковість, властива всій живій природі.
У генетичній інформації закладена здатність розвитку окремих властивостей і ознак. Ця здатність реалізується тільки у певних умовах середовища. Одна і та сама спадкова інформація у змінених умовах може проявлятися по-різному. Так, у примули забарвлення квіток (червоне або біле), у кролів гімалайської породи та сіамських котів характер пігментації волосяного покриву на різних частинах тіла визначається зовнішньою температурою (на більш охолоджених ділянках шерсть темна, бо у цих організмів є мутантний фермент тирозиназа). Отже, успадковується не готова ознака, а певний тип реакції на дію зовнішнього середовища.
Діапазон мінливості, у межах якої залежно від умов середовища один і той самий генотип здатний давати різні фенотипи, називають нормою реакції. У примули генотип такий, що червоний колір квіток з’являється при температурі 15 – 200С, білий – при більш високій температурі, але ні при якій температурі не спостерігається блакитні, сині, фіолетові або жовті квітки. Така норма реакції цієї рослини за ознакою пігментації квіток.
У ряді випадків у одного і того самого гена залежно від усього генотипу і зовнішніх умов можлива різна форма вияву фенотипу: від майже повної відсутності контрольованої геном ознаки до повної її присутності.
Ступінь прояву ознаки при реалізації генотипу у різних умовах середовища називають експресивністю. Під експресивністю розуміють вираженість генотипного вияву гена. Вона пов’язана з мінливістю ознак у межах норми реакції. Експресивність може проявитися у зміні морфологічних ознак, біохімічних, імунологічних, патологічних та інших показників. Так вміст хлору у поті людини звичайно не перевищує 40 ммоль/л, а при спадковій хворобі муковісцидозі (при одному і тому самому генотипі) коливається від 40 до 150 ммоль/л. Спадкова хвороба фенілкетонурія (порушення амінокислотного обміну) може мати різний ступінь прояву (тобто різну експресивність): від легкої розумової відсталості до глибокої імбецильності (тобто здатності тільки до елементарних навичок самообслуговування).
Одна і та сама ознака може проявлятися у одних організмів і не проявлятися у інших, які мають той самий ген. Кількісний показник фенотипового вияву гена називають пенетрантністю. Вона характеризується співвідношенням особин, у яких даний ген проявляється у фенотипі, до загальної кількості особин, у яких ген міг би проявитися (якщо враховується рецесивний ген, то у гомозигот, якщо домінантний – то у домінантних гомозигот і гетерозигот). Якщо, наприклад, мутантний ген проявляється у всіх особин, говорять про 100% – ну пенетрантність, у решті випадків – про неповну і вказують процент особин, у яких проявляється ген. Так, успадкованість груп крові у людини за системою АВО має стопроцентну пенетрантність, спадкові хвороби – неповну: епілепсія – 67%, цукровий діабет – 65%, природжений вивих стегна – 20% пенетрантності тощо.
Терміни «експресивність» і «пенетрантність» введені у 1927 р. М. В. Тимофеєвим-Ресовським. Експресивність і пенетрантність підтримуються природним добором. Обидві особливості необхідно мати на увазі при вивченні спадковості у людини. Необхідно пам’ятати, що гени, які контролюють патологічні ознаки, можуть мати різну пенетрантність і експресивність, тобто проявлятися не у всіх носіїв аномального гена, і що у хворих рівень хворобливого стану неоднаковий. Змінюючи умови середовища, можна впливати на прояв ознак.
Плейотропія (грец. pleison – повний, tropos – спосіб) – залежність кількох ознак від одного гена, тобто множинна дія одного гена. Це явище було уперше виявлене Менделем, хоча він його спеціально не досліджував. За його спостереженнями, у рослин з пурпуровими квітками черешки листків завжди мали червоний колір, а шкірка насіння була білою. Ці три ознаки визначалися дією одного гена. У людини відома спадкова хвороба – арахнодактилія («павукові пальці» – дуже тонкі і довгі пальці), або хвороба Марфана. Ген, який відповідає за цю хворобу, викликає порушення розвитку сполучної тканини і одночасно впливає на розвиток кількох ознак: довжину пальців, порушення будови кришталика ока, аномалії в серцево-судинній системі.
Плейотропічна дія гена може бути первинною і вторинною. При первинній плейотропії ген проявляє і свою множинну дію. Наприклад, змінений білок взаємодіє з цитоплазмою різних клітинних систем або змінює властивості мембран у клітинах кількох органів.
При вторинній плейотропії є один первинний фенотиповий прояв гена, услід за яким розвивається ступінчастий процес вторинних проявів, які приводять до множинних ефектів. Так, при серпоподібноклітинній анемії у гомозигот спостерігається кілька патологічних ознак: анемія, збільшена селезінка, ураження шкіри, серця, нирок і мозку. Тому гомозиготи гинуть, як правило, у дитячому віці. Першопричиною, безпосереднім фенотиповим проявом дефектного гена є аномальний гемоглобін і еритроцити серпоподібної форми. Внаслідок цього відбуваються інші патологічні процеси: злипання і руйнування еритроцитів, анемія, дефекти у нирках, серці, мозку. Ці патологічні ознаки є вторинними. Більше розповсюдження має вторинна плейотропія.
Той факт, що один і той самий генотип може стати джерелом розвитку різних фенотипів, має суттєве значення для медицини. Це значить, що обтяжена спадковість не обов’язково має проявлятися. Багато залежить від тих умов, у яких знаходиться людина. У ряді випадків хворобу як фенотиповий вияв спадкової інформації можна відвернути дотриманням дієти або використанням лікарських препаратів. Реалізація спадкової інформації знаходиться у прямій залежності від середовища. Їхню взаємозалежність можна сформулювати у вигляді певних положень.
- Оскільки організми є відкритими системами, які є єдине ціле з умовами середовища, то і реалізація спадкової інформації відбувається під контролем середовища.
- Один і той же генотип здатний дати різні фенотипи, що визначається умовами, в яких реалізується генотип у процесі онтогенезу особини.
- У організмі можуть розвиватися тільки ті ознаки, які зумовлені генотипом. Фенотипова мінливість у межах норми реакції відбувається за кожною конкретною ознакою.
- Умови середовища можуть вплинути на ступінь вираженості спадкової ознаки в організмах, які мають відповідний ген (експресивність), або на кількість особин, які проявляють відповідну ознаку (пенетрантність).
Мінливість, як і спадковість, властива всім живим організмам. Розрізняють спадкову (генотипічну) мінливість і неспадкову (фенотипічну) мінливість.
Неспадкова (фенотипічна) мінливість поділяється на 2 види: модифікаційну й онтогенетичну.
Модифікаційна мінливість – це неспадкова зміна ознак, що виникає в результаті дії зовнішнього середовища на соматичні клітини і не впливає на спадковий апарат. Модифікаційна мінливість фенотипу відбувається в межах норми реакції. Зміни ознак, що виникають унаслідок різних умов існування, називаються модифікаціями.
Модифікації носять адаптивний (пристосувальний) характер і мають значення для виживання організмів, виконуючи певну роль у збереженні виду. Модифікаційна мінливість – явище масове. Вона торкається не одного, а сукупності об’єктів, і тому її ще називають груповою, або визначеною.
Під впливом зовнішніх умов змінюються ріст тварин і рослин, їхня вага, колір, тощо. Виникнення модифікацій пов’язане з тим, що умови середовища впливають на ферментативні реакції, які відбуваються в організмі і певним чином змінюють їх хід (підсилення пігментації під впливом ультрафіолету, розвинення м'язево-кісткової системи під впливом фізичного навантаження).
Онтогенетична мінливість спостерігається під час індивідуального розвитку особини, у період від запліднення до смерті.
Онтогенез – це повний цикл індивідуального розвитку кожної особини, в основі якого лежить реалізація спадкової інформації на всіх стадіях існування у певних умовах навколишнього середовища; він починається утворенням зиготи (при статевому розмноженні) і закінчується смертю.
Спадкова мінливість – це здатність до змін самого генетичного матеріалу, а неспадкова – здатність організму реагувати на умови зовнішнього середовища, змінюючись в межах норми реакції, заданої генотипом.
Спадкова (генотипічна) мінливість у свою чергу поділяється на комбінативну і мутаційну.
Комбінативна (рекомбінативна) мінливість, яка залежить від перекомбінування алелів у генотипах нащадків порівняно з генотипами батьків, призводить до появи нових ознак у дітей.
Існує 3 механізми комбінативної мінливості:
- Незалежне розходження хромосом при мейозі.
- Рекомбінація генів завдяки кросинговеру.
- Випадкове поєднання при заплідненні.
Явище комбінативної пояснює причину поєднання
в дітей та онуків ознак родичів по батьківській і материнській лініях.
Мутаційна мінливість.
Термін «мутація» запровадив Г. де Фріз (1901) для характеристики випадкових генетичних змін в Oenotera lamarkiana.
Мутаційна мінливість виникає раптово в результаті взаємодії організму і середовища без схрещування. Мутації виникають раптово, що іноді різко відрізняє організм від вихідної форми. У результаті мутацій з'являються і успадковуються аномалії в будові тіла, спадкові хвороби людини. З мутаційною мінливістю пов’язана еволюція – процес утворення нових видів, сортів, порід.
За характером змін генетичного апарату розрізняють мутації, які зумовлені:
а) зміною кількості хромосом (геномні);
б) зміною структури хромосом (хромосомні
аберації);
в) зміною молекулярної структури гена (генні).
Геномом називається сукупність усього спадкового матеріалу організму – ДНК ядра і органел). Мутації, які спричиняють зміну кількості хромосом, називають геномними. До них належать поліплоїдія і гетероплоїдія (анеуплодія).
Поліплоїдія – збільшення диплоїдної кількості хромосом шляхом додавання цілих хромосомних наборів у результаті порушення мейозу.У поліплоїдних форм спостерігається збільшення числа хромосом, кратне галоїдному набору: 3n – триплоїд, 4n – тетраплоїд, 5n – пентаплоїд. Еволюція квіткових рослин йшла шляхом поліплоїдії. У селекційній практиці з метою отримання поліплоїдів на рослини діють критичними температурами, іонізуючим випромінюванням, хімічними речовинами.
Гетероплоїдія виникає в результаті порушення мейозу і мітозу, кількість хромосом може змінюватися і ставати некратною галоїдному. Явище, коли будь-яка хромосома у генотипі має не 2 , а 3 гомологічні хромосоми, називається трисомією, а організм називається трисоміком (2n+1). Три- сомиками є люди з синдромом Дауна.
Протилежне трисомії явище, тобто втрата однієї хромосоми з пари у диплоїдному наборі, називається моносомією, а організм – моносоміком, його каріотип 2n-1 (синдром Шерешевського-Тернера 45, Х0).
Хромосомні аберації виникають в результаті перебудови хромосом.
Розрізняють 4 основні типи хромосомних аберацій:
- Делеція (нестача) – виникають внаслідок утрати хромосомою тієї чи іншої ділянки.
- Дуплікація (подвоєння) – пов’язана з включенням зайвого дублюючого відрізка хромосоми.
- Інверсії – повернення ділянки хромосоми на 180 градусів.
- Транслокація – ділянка хромосоми з однієї пари прикріплюється до негомологічної хромосоми (варіант синдрому Дауна).
Генні мутації змінюють структуру самого гена. Мутації можуть змінювати ділянки молекули ДНК різної довжини.
Чинники, які здатні проникати в соматичні або зародкові клітини живих організмів і викликати мутації, називаються мутагенами. вони можуть бути хімічної, фізичної або біологічної природи.
До найбільш активних фізичних мутагенів належить іонізуюче випромінювання та ультрафіолетові промені.
Хімічних мутагенів надзвичайно багато. Мутагенну активність проявляють 4 умовних групи хімічних факторів зовнішнього середовища: пестициди, промислові отрути, харчові домішки, ліки. Мутагенами є смоли, що містяться в тютюновому димі.
Біологічні мутагени – це віруси, бактерії, найпростіші, гельмінти. Вони спричиняють зміни такого самого характеру, як фізичні та хімічні мутагени.
Учені відслідковують винаходи і поширення спадкової патології, використовуючи так званий генетичний моніторинг. Це комплексне обстеження певних груп населення, спрямоване на реєстрацію подій генетичного характеру, виявлення нових мутацій, успадкованих від попередніх поколінь.
Мутаційний процес – явище, властиве всій живій природі. людина не є винятком. Цей процес проходить постійно й інтенсивно на генному, хромосомному і геномному рівнях. Близько 20% усіх спадкових хвороб у кожному з поколінь після народження – це хвороби, зумовлені новими мутаціями.
Антимутагени. Відомо понад 500 сполук, в яких доведено антимутагенний вплив – здатність їх до захисту геному від дії мутагенів. Речовини з таким спрямуванням підвищують стійкість клітин до негативного впливу мутагенів, знижують кількість пошкоджених мутагеном клітин. Так, прийом -каротину (25 мг), вітаміну С (100 мг), вітаміну Е (280 мг) зменшує пошкодження ДНК у лімфоцитах периферичної крові. Поліфенольні антиоксиданти, які містяться у зеленому чаї, звичайно зменшують частоту сестринських хроматичних обмінів у лімфоцитах периферичної крові у робітників, які контактують з кам’яно-вугільними смолами та вугільним пилом.
Застосування антимутагенів спрямоване на максимальну стійкість клітин людини та захист геному людини від мутагенних впливів.
Лекція 7 Хромосомна теорія спадковості. Зчеплення зі статтю
Хромосоми відіграють велику біологічну роль і мають пряме відношення до передачі спадкових властивостей. Роль хромосом у передаванні спадкової інформації була доведена завдяки: а) відкриттю генетичного визначення статі; б) встановленню груп зчеплення ознак, які відповідають кількості хромосом; в) побудові генетичних, а потім і цитологічних карт хромосом. Обґрунтування хромосомної теорії дано у роботах Т. Моргана К. Бріджеса і А. Г. Стертеванта.
У всіх прикладах схрещування, наведених вище, мало місце незалежне комбінування генів, що належать до різних алельних пар. Це можливо лише тому, що названі гени локалізовані у різних парах хромосом. Проте кількість генів значно перевищує кількість хромосом. Отже, у кожній хромосомі локалізовано багато генів, які успадковуються разом. Гени, локалізовані в одній хромосомі, називаються групою зчеплення. Природно, що у кожного виду організмів кількість груп зчеплення дорівнює кількості пар хромосом, тобто у гороху – 7, у кукурудзи – 10, у томатів – 12, але у організмів, які мають гетеросоми, кількість груп зчеплення буде більше на 1, ніж кількість пар хромосом. Так, у дрозофіли 4 пари хромосом, а груп зчеплення буде 5, у людини 22 пари аутосоми та 2 статеві Х і Y, тому 24 групи зчеплення.
Проте виявилось, що гени, які знаходяться в одній хромосомі, зчеплені не абсолютно. Під час мейозу, при кон’югації хромосом гомологічні хромосоми обмінюються ідентичними ділянками. Цей процес називають кросинговером, або перехрещенням. Кросинговер може відбуватися у будь-якій ділянці хромосоми, навіть у кількох місцях однієї хромосоми. Чим більша відстань між окремими локусами, тим частіше між ними відбувається перехрестя і обмін ділянками.
Морган вдало обрав для дослідження муху-дрозо-філу, яка згодом стала класичним об’єктом для генетичних експериментів. Дрозофіл легко утримувати в лабораторії, вони мають значну плодючість, швидку зміну поколінь (через 1,5 – 2 тижні), невелике число хромосом, що спрощує спостереження.
Явище зчепленого успадковування Т. Морган встановив на такому досліді (рис.7).
У дрозофіли гени довжини крил (М – нормальні, м – короткі) і кольору тіла (В – сірий, в – чорний) локалізовані в гомологічних хромосомах, тобто належать до однієї групи зчеплення. Якщо муху, яка має обидва рецесивні алелі (bbmm), схрестити з гомозиготою за домінантними алелями (ВВММ), то в першому поколінні все потомство виявляється гетерозиготним з домінантними виявами ознак у фенотипі (ВbМт). Тут ще немає ніякої відмінності від звичайного дигібридного схрещування. Для виявлення якості гамет першого покоління необхідно провести аналізуюче схрещування, взявши гібридну самку і рецесивного за обома ознаками самця (тобто чорного короткокрилого).
Якщо два гени, які належать до різних алельних пар, локалізовані у різних хромосомах, то дигетерозигота буде утворювати чотири типи гамет: 25% ВМ, 25% Вт, 25% bМ і 25% bm. При аналізуючому схрещуванні такі дигетерозиготи дадуть чотири типи нащадків: сірих довгокрилих, сірих короткокрилих, чорних довгокрилих і чорних короткокрилих у співвідношенні 1:1:1:1.
У нашому експерименті такого співвідношення потомків не буде: В і М знаходяться в одній групі зчеплення, обидва домінантні алелі локалізовані в одній хромосомі, тому гени В і М незалежно один від одного не комбінуються. При абсолютному зчепленні обох генів необхідно очікувати тільки два типи гамет: 50% ВМ і 50% – bm. При аналізуючому схрещуванні має утворитися половина мух сірих довгокрилих, а половина – чорних короткокрилих.
Фактично ж гібридна самка при аналізуючому схрещуванні дає таке потомство:
41,5% сірих довгокрилих;
8,5% чорних довгокрилих;
41, 5% чорних короткокрилих;
8,5% сірих короткокрилих.
Переважання сірих довгокрилих і чорних короткокрилих мух вказує на те, що алелі ВМ і bm тісно зчеплені. Особини таких фенотипів утворюються з гамет, у яких ці хромосоми не перехрещуються. З іншого боку, поява сірих короткокрилих і чорних довгокрилих свідчить про те, що у певній кількості випадків відбувається порушення зчеплення між алелями В і М та b і m. Це наслідок обміну (кросинговеру) ідентичними ділянками хромосом.
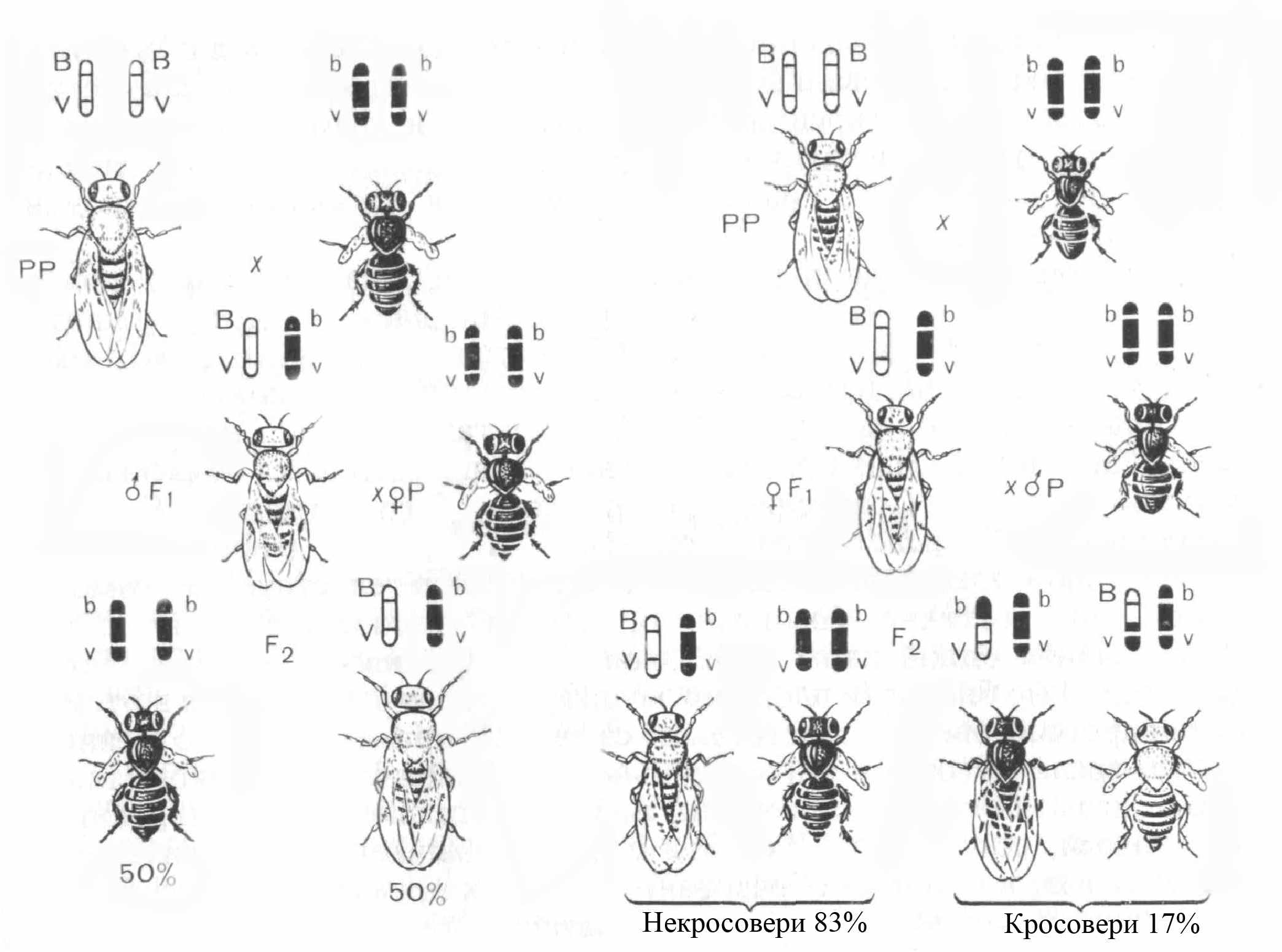
Рисунок 7 – Дослід Т. Моргана
На підставі отриманих даних Морган припустив, що гени, які визначають забарвлення тіла і форму крил, розміщені в одній хромосомі, але у процесі мейозу під час утворення гамет гомологічні хромосоми можуть обмінюватися ділянками – відбувається кросинговер.
Обмін ділянками між гомологічними хромосомами має велике значення для еволюції, тому що значно збільшує можливості комбінування мінливості. Внаслідок перехресту добір у процесі еволюції йде не за цілими групами зчеплення, а за групами генів і навіть окремими генами. Адже в одній групі зчеплення можуть знаходитися гени, які кодують поряд з адаптивними (пристосувальними) і не адаптивні стани ознак. У результаті перехресту «корисні» для організму алелі можуть бути відділені від «шкідливих» і, отже, виникнути більш корисні для існування види генів, комбінації генів – адаптивні.
Лінійне розташування генів. Генетичні карти еукаріотів
Існування кросинговеру дозволило школі Моргана розробити у 1911-1914 рр. принцип побудови генетичних карт хромосом. У основу цього принципу покладене уявлення про розміщення генів вздовж хромосоми лінійно. За одиницю відстані між двома генами домовились брати 1% перехрещення між ними. Цю величину називають сантиморганідою, на честь генетика Т. Г. Моргана.
Припустимо, що до однієї групи зчеплення належать гени А і В. Між ними виявлене перехрещення у 10%. Отже, ці гени знаходяться на відстані 10 одиниць (сантиморганід). Припустимо також, що до цієї самої групи зчеплення належить ген С. Щоб знайти його місце у хромосомі, треба з’ясувати, який процент перехрещення він дає з кожним з двох уже відомих генів. Наприклад, якщо з А він дає 3% перехрещення, то можна припустити, що ген С знаходиться або між А і В, або у протилежному кінці, тобто А розташований між С і В. Якщо між В і С виявиться перехрещення у 7%, то на хромосомі їх необхідно розташувати у такому порядку, як показано на верхній схемі .
Якщо ж між В і С перехрещення 13%, то розміщення генів у хромосомі має бути таким, як на нижній схемі.
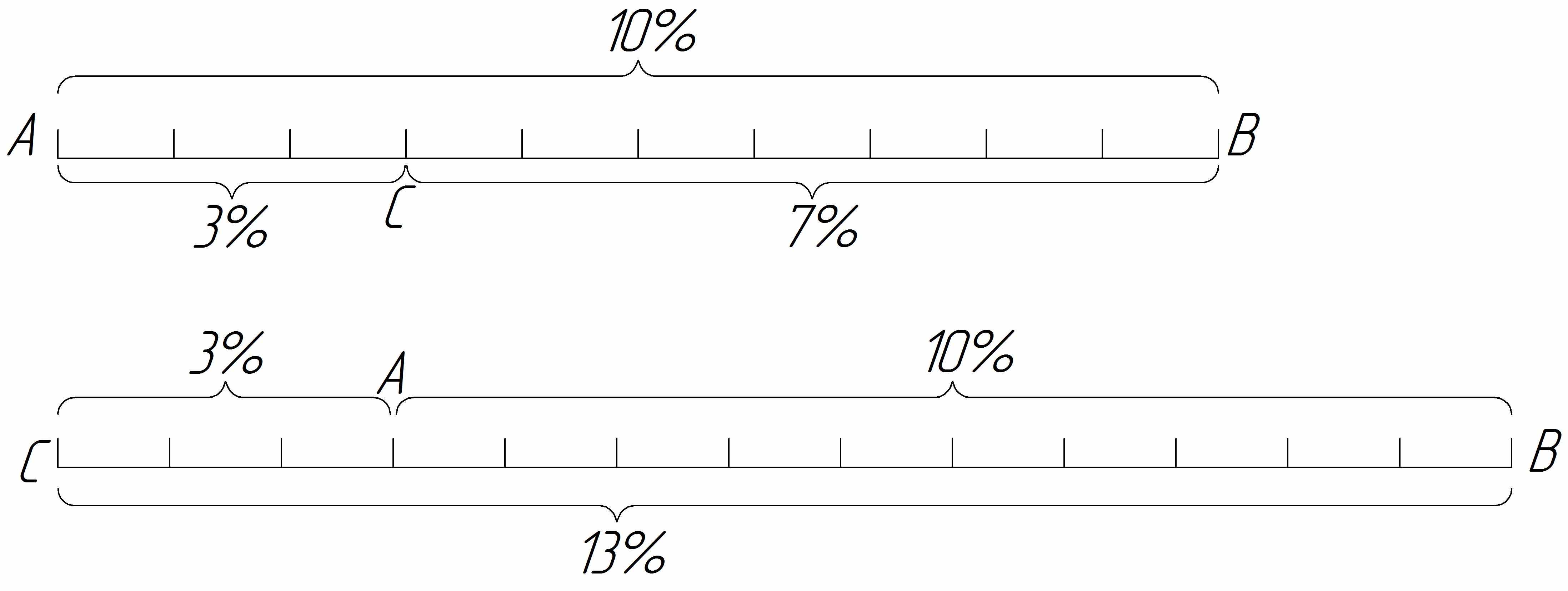
У загальному вигляді цю закономірність можна виразити такою формулою: якщо гени А, В, С належать до однієї групи зчеплення і відстань між генами А і В дорівнює k одиницям, а відстань між В і С дорівнює m одиницям між А і С може бути або k + m або k - m.
Найбільш детальні карти хромосом складені для дрозофіли, яка давно стала класичним генетичним об’єк-том. Із рослинних об’єктів порівняно добре у цьому відношенні вивчені кукурудза і томати, з тварин – кури і миші. Складені карти хромосом людини для всіх 24 груп зчеплення.
Генетичні карти хромосом будують на базі гібридологічного аналізу. Проте, для дрозофіли використовують значно інший спосіб побудови генетичних карт хромосом. Справа в тому, що в клітинах слинних залозах личинок мух знаходяться гігантські (політенні) хромосоми, які перевищують розмір хромосом з інших клітин у 100-200 разів і містять у 8000 разів більше хромонем. Виявилося, що в тих випадках, коли гібридологічним методом виявлялися які-небудь порушення спадкування, відповідні їм зміни мали місце і у гігантських хромосомах. У результаті співставлення генетичних і цитологічних даних стало можливим побудувати цитологічні карти хромосом. Це відкриття підтверджує вірність тих принципів, що були покладені в основу побудови генетичних карт хромосом.
Карти хромосом прокаріотів
Розроблені два способи складання карт хромосом прокаріот. Вони базуються на існуванні кон’югації у бактерій. В основі першого способу лежить можливість часткового переходу хромосоми із бактерії-донора у бактеріюреципієнт, який триває близько 2 год. Бактерії під час кон’югації сполучені дуже слабо, і розрив їх найчастіше відбувається до повного переходу хромосоми. Ця особливість використовується для з’ясування порядку розміщення генів у хромосомі. Мається на увазі, що послідовність генів і відстань між ними пропорціональна часові, протягом якого здійснювалась кон’югація. Штучно припиняючи кон’югацію через певні відрізки часу і з’ясовуючи, які гени за цей час перейшли у реципієнтну клітину, можна встановити порядок їх розташування.
Другий метод полягає в тому, що в результаті кон’югації у бактерії-реципієнта частина хромосоми подвоюється. Протягом кількох поколінь бактерій ця ділянка залишається подвоєною (диплоїдною). Такі особини використовуються для з’ясування того, який із генів є домінантним, а який – рецесивним. Як правило, після кількох поділів низка генів, що локалізовані у ділянці хромосоми із донорської бактерії, шляхом кросинговеру включаються у гомологічні локуси хромосоми бактерії-реципієнта і замінюють алельні гени, а інші гени донора елімінуються (зникають). Утворену рекомбінантну хромосому можна використовувати для вивчення локалізації генів за принципом, який зроблений для еукаріот.
Закономірності, які встановлені школою Моргана, а потім підтверджені і поглиблені на численних об’єктах, відомі під загальною назвою хромосомної теорії спадковості.
Основні її положення такі:
- Гени розміщені у хромосомах по довжині в лінійному порядку; різні хромосоми містять неоднакову кількість генів.
- Алельні гени займають певні й ідентичні локуси (місця) гомологічних хромосом.
- Гени, розташовані в одній хромосомі, утворюють групу зчеплення, завдяки чому має місце зчеплення деяких ознак, які разом передаються нащадкам.
- Між генами гомологічних груп зчеплення, завдяки кросинговеру, можуть відбуватися рекомбінації.
- Сила зчеплення між генами обернено пропорційна відстані між ними. Чим ближче розміщені гени в одній хромосомі, тим менше частота кросинговеру і тим сильніше їх зчеплення. Відстань між генами вимірюється у процентах кросинговеру.
- Кожний біологічний вид характеризується специфічним набором хромосом – каріотипом.