
Щербицкий Евгений Павлович в 1965 г окончил кгпи. Доцент кафедры экономической географии. Отличник народного просвещения рф, награжден грамотой морф
| Вид материала | Документы |
- Султанов Адгам Яхиевич кандидат физ мат наук, доцент, отличник народного образования,, 70.81kb.
- Методика преподавания математики в высшей школе. Телефон кафедры, 21.08kb.
- Формирование культуры здорового образа жизни учащихся в условиях здоровьесберегающей, 55.5kb.
- Автобиография Комаров А. А. учитель технологии гоу сош №1973, 25.42kb.
- Профессорско-преподавательский состав кафедры экономики, финансов и бухгалтерского, 325.83kb.
- А. С. Пушкина Составитель: Смирнова, 71.42kb.
- Учебная программа для специальности: 1-31 02 01 География направление 1- 31 02 01-02, 159.07kb.
- Учебная программа для специальности: 1-31 02 01 География по направлению 1-31 02 01-02, 311.94kb.
- Донец Любовь Алексеевна имеет педагогический стаж работы 35 лет, по должности учитель, 29.25kb.
- Программа элективного курса по литературе «Драма как род литературы. Русский драматический, 195.8kb.
2.Гнездовая экология популяций некоторых широко
распространенных видов птиц на разных высотах Саянских гор
При сравнении были рассмотрены данные по экологии воробьиных видов птиц, популяции которых имеют широкое распространение как на равнине, так и в высокогорном ландшафтном ярусе Саян.
2.1. Фенология прилета и отлета некоторых видов птиц
на разных высотах Саян
Фенологическая картина появления перелетных птиц в местах гнездования в пределах горных систем представляется достаточно сложной. Ее понимание зачастую бывает затруднено транзитно пролетающими особями из популяций других широт. Особенности положения горной области или ее морфология еще больше усугубляют эту задачу. Известно, что ранней весной и на равнине в умеренных широтах погодные факторы (низкие температуры, осадки, ветер) могут периодически оказывать влияние на репродуктивный цикл птиц, задерживая размещение самцов по гнездовым территориям, формирование пар и гнездование (Птушенко, 1962). В горах на значительных высотах эти процессы довольно обычны и могут происходить не только весной, но и в течение всего репродуктивного периода. Они ярче выражены, чем на равнине (Ирисов, 1992а).
В горах умеренных и высоких широт раннеприлетные виды в первое время локализуются в пониженных элементах рельефа (долинах, поймах, седловинах), на южных склонах хребтов и отдельных массивов, у ручьев и рек, в местах, освободившихся от снега и защищенных от ветра (Ирисов, 1997). Достигшие мест размножения многие горные и особенно высокогорные птицы накануне и в ходе репродуктивного цикла оказываются в неблагоприятных условиях из-за частых в этот период времени снежных буранов, холодных ветров, заморозков и снегопадов.
За время исследований в различных районах гор Южной Сибири был зарегистрирован ряд случаев массовой гибели птиц из-за неблагоприятных метеорологических условий в течение всего летнего периода. Иногда при понижении температуры воздуха (до –5… –7С) и обильных снегопадах глубина снежного покрова на вершинах хребтов достигала 40-60 см и сохранялась на протяжении 2-3 дней. В это время наблюдалось резкое увеличение численности птиц в предгорьях и поймах рек за счет откочевывающих из верхнего пояса гор насекомоядных видов: трясогузок, коньков, соловьев, чеканов, завирушек, среди которых отмечался наибольший процент гибели (за один час экскурсии – 4-6 погибших птиц). Такие ситуации были зафиксированы в середине июня 1991 г. на хр. Цаган-Шибэту (Тыва), в конце июня 1991 г. на хр. Сайлыг-Хем-Тайга (Западный Саян), в начале июня 1992 г. в северо-западной части Восточного Саяна (Кутурчинское белогорье) и в середине августа 1996 г. на Агульских белках (Восточный Саян).
Таблица 2
Проявление сезонных аспектов у некоторых видов птиц на
малых (верхняя строка*) и больших (нижняя строка**) высотах в центральной и
северо-западной частях Восточного Саяна (в среднем за весь период исследований)
| Вид | Весна | Осень | |||
| дата первого появления | массовый прилет | начало отлета | массовый отлет | даты последних встреч | |
| Лесной конек | 18/IV* | 30/IV-15/V | | 29/VIII-12/IX | 23/IX |
| 16/V** | 28/V-10/VI | 18/VIII | 24-30/VIII | 8/IX | |
| Горная трясогузка | 26/IV | 10-25/V | | 30/VIII-15/IХ | 26/IX |
| 7/V | 18-30/V | 18/VIII | 25-30/VIII | 15/IX | |
| Черноголовый чекан | 9/V | 16-27/V | | 28/VIII-15/IX | 2/X |
| 17/V | 25/V-5/VI | 9/VIII | 18-27/VIII | 14/IХ | |
| Краснозобый и чернозобый дрозды | 12/IV | 19-24/IV | | 10-22/IX | 9/X |
| 5/V | 21-30/V | 15/VIII | 25-31/VIII | 14/IХ | |
| Обыкновенная чечевица | 8/V | 15-25/V | | 1-18/IХ | 24/IХ |
| 19/V | 28/V-7/VI | 19/VIII | 25-30/VIII | 11/IХ | |
Аналогичные ситуации, как характерное явление в горных областях, описывали и другие авторы (Козлов, 1899; Сушкин, 1938; Ковшарь, 1965; Потапов, 1966б; Иванов, 1969; Ирисов, 1968; Ирисов, Ирисова, 1986; Забелин, 1984; Лаптенок, 1991 и др.).
По мере освобождения пространств от снежного покрова и повышения температуры воздуха птицы вновь перемещаются в высокогорный пояс. Сроки и характер этих перемещений не у всех видов одинаковые, что обусловлено различием реакций на возникновение неблагоприятных погодных условий.
Анализ фенологических показателей свидетельствует о том, что прилет равнинных популяций приходится на более ранние сроки по сравнению с высокогорными (табл.2).
Осенние миграционные перемещения птиц из высокогорного пояса начинаются раньше, чем у предгорных популяций (табл.2). Так, отлет черноголового чекана начинался с середины III декады августа и заканчивался во II декаде сентября, тогда как в лесостепных районах одиночные птицы еще встречались в первых числах октября (Ким, 1996). Первые кочующие стайки лесных коньков и горных трясогузок встречались на Канском, Пезинском и Кутурчинском белогорьях уже 18 августа (массовый отлет с 27-30 августа), в предгорных районах северо-западной части Восточного Саяна массовый отлет начинался с 10-15 сентября. Краснозобый и чернозобый дрозды постепенно откочевывают из высокогорного пояса в нижележащие ландшафты уже с середины августа, а уже в начале сентября они обычны в долинных лесах предгорий и многочисленны в лесостепных районах, местами достигая численности 35-40 особей/км2. В середине октября эти птицы уже не встречались. В отдельные годы чернозобых дроздов встречали зимой: в феврале и марте 1969 г. на водоразделах рек Чибижек, Джебь, Осиповка, в декабре 1968 г. – в районе Красноярской ГЭС (Ким,1996).
2.2. Особенности вокализации некоторых видов птиц на разных высотах
В связи с экологической спецификой высокогорного пояса существуют некоторые адаптивные особенности в активности и интенсивности пения птиц. Изучение суточной активности и интенсивности пения проводилось у 2 видов, встречающихся в различных биотопах предгорий и высокогорного пояса Саян (лесной конек и обыкновенная чечевица), а также у горного конька и гималайской завирушки в гольцовом поясе.
Поскольку широта местности и продолжительность дня существенно не меняются, у птиц, наблюдавшихся на разной высоте над уровнем моря, продолжительность пения в течение суток одинакова (за исключением гималайской завирушки) и составляет 16 ч. (рис. 1-3).
Утренний пик интенсивности пения лесного конька в предгорье наступает в 10 ч, затем плавно снижается к 15 ч и, резко поднимаясь, достигает максимума к 17 ч. В оставшуюся часть суток интенсивность пения падает. Форма кривой интенсивности пения лесного конька в высокогорье однотипна с предгорной. Однако с 9 до 18 ч она держится на почти неизменно высоком уровне. На общем уровне интенсивности едва возвышаются дневной пик между 11 и 12 ч и вечерний – в 18 ч.
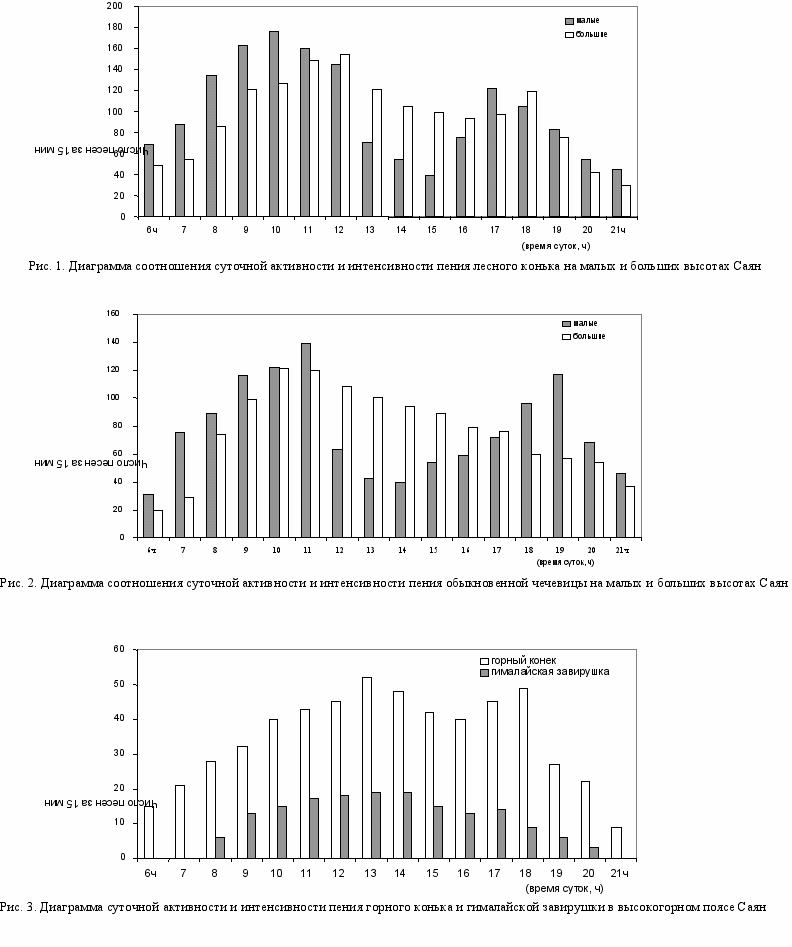
У обыкновенной чечевицы, так же, как и у лесного конька, в предгорье наблюдалось два четко выраженных пика интенсивности пения (рис. 2). В высокогорном поясе у нее был только один пик вокальной интенсивности – с 10 до 12 ч, а затем наблюдалось плавное снижение.
Оба вида в своем распространении в высокогорном поясе достигают его нижней части– субальпийских редколесий и кустарников – биотопов, сохраняющих определенное экологическое сходство с лесными местообитаниями предгорий. В связи с этим интенсивность пения в горах у них не намного ниже по сравнению с предгорными популяциями и относительно равномерно рассредоточена в течение дня, что определяется частой сменой погоды на больших высотах (рис. 1-2).
Совершенно иная ситуация складывается в верхней части гольцового пояса Саян – в каменистых тундрах, на скальных гребнях у снежников и ледников. Такие виды, как гималайский и сибирский вьюрки, большая чечевица, поют упрощенные песни, состоящие, как правило, из короткого щебета и мелодичной позывки. У горного конька и гималайской завирушки наблюдается снижение вокальной интенсивности (рис. 3).
Причина такого рода различий заключается в том, что звуковая среда в лесах, будь то на равнине или в горах, характеризуется насыщенностью биогенными звуками и широким их спектром (от 400 до 50000 Гц и выше), тогда как открытые пространства высокогорий отличаются наименьшей насыщенностью биогенными звуками, спектр которых смещен в сторону высоких и средних частот (Ильичев, 1964). Следовательно, слуховой анализатор птиц в лесах, где ограничено действие зрительного анализатора, должен функционировать в приоритетном режиме, влияя, таким образом, на совершенствование вокальных способностей лесных птиц. В условиях открытых пространств высокогорий приоритетная роль принадлежит зрительному анализатору (Ирисов, 1997). Логично ожидать, что звуковая гамма песни лесных птиц должна быть с богатой тональной структурой, а вокальная активность – большей продолжительностью. Напротив, птицы, обитающие в открытом ландшафте, должны уступать им по всем этим показателям, тем более что погодные условия в гольцовой зоне характеризуются крайней нестабильностью (частые дожди, снегопады, более низкая, чем на равнине, среднесуточная температура).
Таким образом, интенсивность пения у горных популяций изученных видов певчих птиц в нижней части высокогорья меняется незначительно и не уступает таковой у предгорных форм, вместе с тем, она выше, чем у видов, обитающих исключительно в верхней части высокогорного пояса.
Интенсивность пения в течение сезона размножения также зависит от ряда причин, но главной, по-видимому, является соответствие той или иной фазе репродуктивного цикла. Так, у изученных видов в предгорье первый максимум интенсивности пения предшествовал кладке, а второй наступал в период массового выкармливания птенцов и, по-видимому, обозначал подготовку ко второй кладке. В высокогорном поясе первый максимум непосредственно совпадал с кладкой, а второй пик начинался после того, как в большинстве гнезд выкармливание заканчивалось и начинался вылет молодых. Судя по характеру интенсивности пения, первая кладка в предгорьях бывает несколько более растянутой, чем в высокогорье.
Рассмотренная специфика пения птиц в высокогорье, по-видимому, носит адаптивный характер (Ирисов, 1997).
2.3. Расположение и устройство гнезд
При выявлении особенностей гнездостроения у воробьиных птиц в условиях предгорий и высокогорий особое внимание обращалось на характер расположения гнезд, их размеры и строительный материал.
В предгорных биотопах краснозобый и чернозобый дрозды для устройства гнезд (n=29) использовали в 15 случаях пихту, в 6 – сосну, в 3 – кедр, в 5 – буреломные пни. Гнезда находились на высоте от 0,5 до 8 м, чаще – от 1,5 до 3 м (Ким, 1996). Гнезда (n=27), найденные в высокогорном поясе, располагались на высоте 1,5-5 м (от 1 до 7 м) обычно на кедре и лиственнице, реже на пихтовом стланнике (субальпийское редколесье), в поясе субгольцовых кустарников и ерниковой тундре на ольхе, карликовой березке и иве в пределах высот 0,5-1,5 м от земли. Независимо от высоты местности для строительства гнезд дрозды использовали одни и те же материалы (сухие травянистые растения, тонкие веточки, хвою пихты и кедра, мох, глину).
Обыкновенная чечевица в предгорье строит гнезда (n=19) на подросте ели, сосны, кедра, березе, черемухе, реже на кустарниках, в интервале высот от 0,5 до 7,5 м (в среднем 0,8-2 м). В высокогорных условиях ее гнезда (n=11) в большинстве случаев (62,8%) были устроены на таволге, жимолости, рододендронах, карликовой иве, реже на кедре, лиственнице, пихтовом стланнике на высоте 0,5-1,6 м от земли.
Гнезда лесного конька и черноголового чекана, найденные в предгорных (соответственно n=21 и n=16) и горных местах обитания (n=11 и n=9), размещались на земле. Они представляли собой небольшие углубления, расположенные у основания кочки или под кустом, и состояли из сухих стеблей травянистых растений (осоки, злаков), лоток был выстлан пуховыми перьями, реже сухим мхом, шерстью и волосом млекопитающих.
У горной трясогузки большинство гнезд на малых (n=25) и больших (n=16) высотах располагались по берегам ручьев и рек, вдоль лесных троп, под надежным прикрытием камней, упавших деревьев, корневищ пней, обрывов. В высокогорных биотопах для устройства гнезд горная трясогузка очень часто использует различные формы нанорельефа, чего практически не наблюдалось в предгорье. Так, в горах был зафиксирован ряд случаев расположения гнезд этого вида в расщелинах скал (n=3) и нагромождениях камней (n=6).
Типичный околоводный вид оляпка на малых (n=8) и больших (n=6) высотах устраивал свои гнезда, как правило, под прикрытием обрывистого берега либо скальной ниши в непосредственной близости от воды (от 0,5 до 3,5 м). Гнезда обычно представляли собой массивные сооружения шаровидной или эллипсовидной формы из мха, корешков и стеблей трав с входным отверстием, обращенным в сторону воды.