
Эколого-морфологический анализ хирономид рода cryptochironomus kieffer (diptera, chironomidae) палеарктики 03. 00. 16 экология 03. 00. 09 энтомология
| Вид материала | Автореферат диссертации |
- Хирономиды (diptera, chironomidae) бассейна нижнего амура. Фауна, систематика, распространение, 387.83kb.
- Общая характеристика работы актуальность, 882.91kb.
- Эколого-фаунистический анализ паразитофауны рыб вислинского залива, рек преголи и прохладной, 419.14kb.
- Экология бактерий рода Rhodococcus из глубоководных битумных построек озера Байкал, 306.35kb.
- Тема. Медицинская энтомология, 52.97kb.
- Фауна, биология и экология рода anopheles mg. (Сем. Culicidae) в прибрежных экосистемах, 1214.31kb.
- Региональные программы по решению эколого-экономических задач. Эколого-экономические, 68.04kb.
- Рабочая программа на 2007-2008 учебный год по дисциплине: «энтомология» Специальность:, 172.45kb.
- Характер изменения функционально активных участков и компактности политенных хромосом, 364.37kb.
- Программа «Экология комнатных растений», 44.17kb.
На правах рукописи
Морозова Елена Евгеньевна
ЭКОЛОГО-МОРФОЛОГИЧЕСКИЙ АНАЛИЗ
ХИРОНОМИД РОДА CRYPTOCHIRONOMUS KIEFFER
(DIPTERA, CHIRONOMIDAE) ПАЛЕАРКТИКИ
03.00.16 – экология
03.00.09 – энтомология
Автореферат диссертации на соискание ученой степени
доктора биологических наук
Саратов - 2008
Работа выполнена в Государственном образовательном учреждении высшего профессионального образования «Саратовский государственный университет им. Н.Г. Чернышевского» на кафедре морфологии и экологии животных
Научные консультанты: доктор биологических наук, профессор
Заслуженный деятель науки РФ
Шляхтин Геннадий Викторович
доктор биологических наук, профессор
Зинченко Татьяна Дмитриевна
Официальные оппоненты: доктор биологических наук, профессор,
член-корреспондент РАН
Малахов Владимир Васильевич
доктор биологических наук,
старший научный сотрудник
Петрова Нинель Алексеевна
доктор биологических наук, профессор
Сачков Сергей Анатольевич
Ведущая организация: Институт проблем экологии и эволюции
им. А.Н. Северцова РАН
Защита состоится « 14 » ноября 2008 г. в 10.00 часов на заседании диссертационного совета Д 212.243.13 при Государственном образовательном учреждении высшего профессионального образования «Саратовский государственный университет им. Н.Г. Чернышевского» по адресу: 410012 г. Саратов, ул. Астраханская, 83, учебный корпус № 5, аудитория № 61, факс (8452) 511635, e-mail: biosovet@sgu.ru
С диссертацией можно ознакомиться в Зональной научной библиотеке имени В.А. Артисевич ГОУ ВПО «Саратовский ГУ»
Автореферат разослан « 10 » октября 2008г.
Ученый секретарь
диссертационного совета С. А. Невский
Общая характеристика работы
Актуальность исследования. Личинки хирономид (Diptera, Chironomidae) – являются важной составляющей сообществ макрозообентоса большинства пресноводных и многих солоноватых водоёмов. Обладая высокой экологической пластичностью, они играют значительную роль в жизни водных экосистем: участвуют в процессах биологического самоочищения, являются объектом питания ценных промысловых рыб, относятся к индикаторам экологического состояния и типологической классификации водоемов (Константинов, 1977; Балушкина, 1987; Макарченко, 2006; Сергеева, 2006; Зинченко и др., 2007; Wiederholm, 1978; Saether, 1979; Rosenberg, Resh, 1993). Имея в ядрах клеток слюнных желез гигантские политенные хромосомы, личинки хирономид широко используются в цитогенетических исследованиях (Кикнадзе и др., 1991, 2005; Петрова, 1992; Шобанов, 2000; Сергеева, 2006).
Особый интерес представляет исследование мало изученных хирономид подсемейства Chironominae, а именно, видов рода Cryptochironomus Kieffer, 1918 широко распространенных в Палеарктике и Неарктике (Шилова, 1976; Saether, 1977; Mason, 1985). В России и сопредельных странах они обычны для водоемов различного типа, включая водохранилища, реки, пруды, горные озера и слабосолоноватые водоемы (Шилова 1966, 1976; Макарченко, 1999; Зорина, 2000; Зинченко, 2002). Трофическая специализация личинок рода Cryptochironomus разнообразна: от всеядных до типично хищных форм (Константинов, 1958; Шилова, 1959, 1976; Луферов, 1961; Извекова, 1975, 1980).
Виды этого рода имеют четкую структуру политенных хромосом, привлекая внимание вариабельностью диплоидных чисел (2n = 4 и 2n = 6), разнообразием функционально активных зон (ядрышек, колец Бальбиани, пуфов), высоким уровнем хромосомного полиморфизма инверсионного типа (Петрова и др., 1981; Белянина, 1983; Кикнадзе и др., 1991; Истомина, 1992). Исследования экологии и систематики видов р. Cryptochironomus из водоемов бассейна Нижней Волги связаны, в основном, с изучением функционально-энергетических характеристик хирономид этого рода (Константинов, 1958; Мисейко, 1966; Шилова, 1976; Белянина, 1983; Каширская, 1986; Балушкина, 1987; Зинченко, 2004) в условиях отсутствия видовой диагностики личинок. В этой связи проведение целенаправленных исследований (систематики, морфологии, биологии, экологии) видов р. Cryptochironomus в водоемах Волжского бассейна позволяет объективно оценить ценотическую роль хирономид в водных экосистемах разного типа.
Цель и задачи исследования. На основе комплексных исследований (систематики, морфологии, биологии, экологии) палеарктических видов р. Cryptochironomus и анализа многолетней динамики структуры сообществ макрозообентоса водоемов бассейна Нижней Волги выявить эколого-морфологические особенности хирономид данного рода и установить закономерности формирования и развития их ценоза в условиях разнотипных водных объектов. Для достижения поставленной цели решались следующие задачи:
- оценить изменения многолетней структурной организации сообществ макрозообентоса и дать количественную оценку различий видовой структуры ценоза хирономид Волгоградского водохранилища и сопредельных водоемов на основании собственных данных и литературных сведений.
- проанализировать современное состояние систематики и диагностической принадлежности (палеарктических и неарктических) видов рода Cryptochironomus. Выявить морфологические признаки личинок, куколок и имаго, имеющих определяющее значение для идентификации видов.
- представить сравнительную характеристику данных о структуре кариотипов хирономид р. Cryptochironomus фауны Палеарктики. Выявить особенности кариотипической изменчивости личинок массовых видов Cryptochironomus obreptans (Walker, 1856) и Cryptochironomus defectus (Kieffer, 1913) из разнотипных водных объектов.
- на основе анализа многолетних данных о видовой структуре ценоза хирономид р. Cryptochironomus как структурообразующего литорального комплекса, выявить состав, распространение, биологические и экологические особенности видов (на примере Волгоградского водохранилища и водоемов бассейна Нижней Волги).
- провести количественную оценку меры воздействия экологических факторов, определяющих формирование ценоза хирономид с доминированием C. оbreptans и C. defectus. Установить индикаторную значимость видов р. Cryptochironomus для оценки состояния гидросистем и районирования модельных водных объектов (на примере участков Волгоградского водохранилища и сопредельных водоемов).
Научная новизна и теоретическая значимость. Изучена систематика, морфологическая и кариотипическая изменчивости видов р. Cryptochironomus. Исследована биология массовых видов и выявлены их экологические особенности (на примере водоемов бассейна Нижней Волги). Впервые исследован состав ценоза хирономид р. Cryptochironomus мелководной зоны Волгоградского водохранилища и водоемов урбанизированной территории.
Впервые обобщен и проанализирован в сравнительном аспекте материал по морфологии 20 палеарктических и неарктических видов хирономид р. Cryptochironomus; получены диагностические данные личинок, куколок, имаго 11 палеарктических видов. Приводится описание имаго самки С. defectus и вида C. andreevi Morozova, sp.n. Дан анализ кариотипов 9 видов р. Cryptochironomus. Выявлен хромосомный полиморфизм в популяциях C. оbreptans и C. defectus в условиях изменения экологических факторов разнотипных водных объектов. Впервые составлена цитофотокарта C. defectus. Выявлены закономерности многолетней динамики формирования ценоза хирономид и биоиндикационной значимости видов р. Cryptochironomus в соответствии с изменением основных факторов среды в водоемах бассейна Нижней Волги как информационная основа для проведения мониторинговых исследований.
Практическая значимость. Определительные таблицы могут использоваться для диагностики личинок и куколок р. Cryptochironomus при систематических, экологических и энтомологических исследованиях. Осуществлена пространственная экологическая дифференциация и районирование модельных водных объектов (на примере участков Волгоградского водохранилища и сопредельных водоемов) на основе многомерного массива разнородных параметров. Количественные характеристики морфологической и кариотипической изменчивости личинок С. оbreptans и C. de-fectus и динамика многолетних показателей численности являются базовой основой в использовании полученных данных для типологической оценки водоемов при проведении мониторинга экологического состояния водных объектов бассейна Нижней Волги.
Материалы по питанию личинок С. оbreptans и C. defectus позволяют объективно определить их роль в трофической структуре донных сообществ. Виды С. оbreptans и C. defectus рекомендованы для использования в биоиндикационных исследованиях при оценке экологического состояния водоемов бассейна Нижней Волги. Полученные данные биоиндикационной роли видов р. Cryptochironomus используются при мониторинговых исследованиях в рамках реализации задач региональной экологической программы «Возрождение Волги». Материалы нашли применение в учебных программах теоретических курсов (биологических факультетов) по экологии, зоологии и энтомологии в Саратовском государственном университете им. Н.Г. Чернышевского, являются составной частью тематического плана научных исследований кафедры морфологии и экологии животных СГУ: «Изучение надорганизменных биологических систем», «Разработка методов контроля за состоянием экосистем» (гос. регистрационный № 8107305), «Комплексное изучение функционирования надорганизменных биологических систем при антропогенном воздействии» (гос. регистрационный № 21071305).
Апробация результатов. Основные результаты исследований представлены на: Всесоюзном совещании «Систематика хирономид и их значение в жизни водных экосистем» (Паланга, 1989); Всероссийских совещаниях по проблемам изучения хирономид (Борок, 1990, 1992, 1995, 2000); V Всесоюзном диптерологическом симпозиуме (Новосибирск, 1990); I Всесоюзной конференции по генетике насекомых (Москва, 1991); II и III Всесоюзных симпозиумах по кариосистематике беспозвоночных животных (Чебоксары, 1991; Москва, 1996); Всероссийской научной конференции «Стратегия природопользования и сохранения биоразнообразия в XXI веке» (Оренбург, 1996); XIII, XIV, XV Международных симпозиумах по хирономидам (Германия, 1997; Бразилия, 2000; США, 2003); VI Всероссийском диптерологическом симпозиуме (Санкт-Петербург, 1997); XI, XII и XIII Съезде Русского энтомологического общества (Санкт-Петербург, 1998, 2002, 2007); Международной научной конференции «Малые реки: Современное экологическое состояние, актуальные проблемы» (Тольятти, 2001); VIII съезде гидробиологического общества РАН (Калининград, 2001); Всероссийской научной конференции «Фундаментальные и прикладные аспекты функционирования водных экосистем: проблемы и перспективы гидробиологии и ихтиологии в ХХI веке» (Саратов, 2001); Международной конференции «Биоразнообразие и динамика экосистем в Северной Евразии» (Новосибирск, 2002); Всероссийской научно-практической конференции «Экологические, морфологические особенности и современные методы исследования живых систем» (Казань, 2003); II Международной конференции «Разнообразие беспозвоночных животных на Севере» (Сыктывкар, 2003); VI Международной научной конференции «Эколого-биологические проблемы бассейна Каспийского моря» (Астрахань, 2003); конференции по хирономидам, посвященной столетию А.А. Черновского (Санкт-Петербург, 2004), Любищевских чтениях (Ульяновск, 2007), научных конференциях и семинарах Саратовского государственного университета им. Н.Г.Чернышевского (1995-2008гг.).
Публикации. По теме диссертации опубликованы 52 работы, в том числе 9 - в изданиях перечня ВАК РФ.
Декларация личного участия автора. Автор лично проводил экспериментальные исследования, выявлял закономерности формирования и развития ценоза хирономид р. Cryptochironomus в условиях разнотипных водных объектов. Анализ полученных данных, их интерпретация и оформление выполнялись автором самостоятельно. В совместных публикациях вклад соискателя составил 50-80%.
Объем и структура диссертации. Диссертация изложена на 305 страницах и включает 125 рисунков и 60 таблиц. Работа состоит из введения, 5 глав, заключения, выводов и приложения. Библиографический указатель включает 355 источников, из которых 135 на иностранных языках.
Основные положения, выносимые на защиту
1. Ценоз хирономид р. Cryptochironomus, формируемый в Волгоградском водохранилище и сопредельных водоемах, характеризуется изменением видового разнообразия и снижением численности популяций ряда видов в результате воздействия природных и антропогенных факторов.
2. В масштабе разнотипных экотопов Волгоградского водохранилища (участки открытых, защищенных мелководий и прудов урбанизированной территории) численность популяций массовых видов хирономид р. Cryptochironomus (C. оbreptans и C. defectus) находится в зависимости от температурного режима водоема и содержания основных биогенных элементов.
3. Для массовых видов хирономид р. Cryptochironomus (С. obreptans и C. defectus) характерен хромосомный полиморфизм, обусловленный различными условиями существования личинок в водоемах разного типа (открытые и защищенные мелководья Волгоградского водохранилища, пруды).
4. В многолетней динамике формирования ценоза хирономид р. Cryptochironomus выделяются виды (C. оbreptans и C. defectus), способные поддерживать максимальную численность во временном и пространственном масштабе при воздействии разных факторов среды.
5. Диагностика близкородственных видов хирономид р. Cryptochironomus, разработанная на основе естественной классификации семейства Chironomidae, возможна с использованием морфологических признаков личинок, куколок, имаго и привлечением кариотипических данных.
Благодарности. Автор признателен и благодарен д.б.н. Г.В. Шляхтину (СГУ), д.б.н. Т.Д. Зинченко (ИЭВБ РАН), д.б.н. Л.А. Чубаревой, д.б.н. Н.А. Петровой, (ЗИН РАН), д.б.н. В.Ф. Шуйскому (СПбГУ), д.б.н. Е.А. Макарченко, к.б.н. О.В. Зориной (Биолого-почвенный институт ДВО РАН), д.б.н. Н.А. Шобанову, д.б.н. А.И. Шиловой, к.б.н. Н.И. Зеленцову, к.б.н. Г.Х. Щербине (ИБВВ РАН); д.б.н. Л.И. Гундериной, к.б.н. А.Г. Истоминой (Новосибирск), д.б.н. В.В. Аникину (СГУ), к.б.н. Э.И. Извековой (МГУ им М.В.Ломоносова), д.б.н. Беляниной С.И., д.б.н. И.В. Сергеевой, д.б.н. Н.В. Полуконовой, к.б.н. Дурновой (СГМУ), к.х.н. Л.А. Выхристюк (ИЭВБ РАН), к.б.н. Ю.А. Малининой (Сар.отд. ГОСНИОРХ), Х. Валлендууку (Голландия) и П. Лангтону (Англия) за творческую поддержку, неоценимую помощь, советы и консультации.
Содержание работы
Во Введении обосновывается актуальность исследования, его практическая и теоретическая значимость; сформулированы основная цель и задачи исследования, а также пути их реализации.
Глава 1. СОВРЕМЕННОЕ СОСТОЯНИЕ СИСТЕМАТИКИ
ХИРОНОМИД ВИДОВ РОДА CRYPTOCHIRONOMUS И ИХ
ЭКОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА (обзор литературы)
Приводятся сведения, отражающие современное состояние систематики и диагностики видов р. Cryptochironomus Kieffer, 1918. Этот род принадлежит к подсемейству Chironominae, трибе Chironomini, входит в состав комплекса Harnischia (Beck and Beck, 1969). Выделен новый подрод Miscellanea Zorina, subgen. n. в составе р.Cryptochironomus (Зорина, 2000). Несмотря на то, что диагностика видов р. Cryptochironomus разрабатывается длительное время, в большинстве гидробиологических и экологических работ до сих пор указываются группы Cryptochironomus gr. defectus (Шилова, 1966) и Cryptochironomus gr. fulvus (Beck and Becк, 1964; Saether, 1977), включающие в себя несколько видов, морфологически трудно различимых на стадии личинки, что не позволяет дать адекватную экологическую характеристику видов. В составе р. Cryptochirono-mus выделено 14 палеарктических видов (Ashe, Cranston, 1990; Saether et. al., 2000): С. albofasciatus (Staeger); С. crassiforceps Goetghebuer; С. defectus (Kieffer); С. denticulatus (Goetghebuer); С. obreptans (Walker); С. psittacinus (Meigen); С. redekei (Kruseman); С. rostratus Kieffer; С. supplicans (Meigen); С. ussouriensis (Goetghebuer); С. tamayoroi Sasa et Ichimori; C. tokaracedeus Sasa et Suzuki; C. hentonensis Hasegawa et Sasa; C. tamaichimori Sasa. Виды C. tokaracedeus Sasa et Suzuki; C. hentonensis Hasegawa et Sasa; C. tamaichimori Sasa в настоящее время считаются не валидными (Макарченко, Зорина, не опубликованные данные). Обобщены сведения по биологии и экологии хирономид (распространению, особенностям питания, циклам развития и др.) наиболее изученных видов р. Cryptochironomus, обитающих в водоемах Волжского бассейна. Анализируется имеющийся опыт использования хирономид в качестве биоиндикаторов в мониторинговых исследованиях поверхностных вод. Приводятся примеры индикационной значимости хирономид на разных уровнях организации биологических систем.
Глава 2. СОВРЕМЕННАЯ ЭКОЛОГИЧЕСКАЯ ОЦЕНКА
РАЙОНА ИССЛЕДОВАНИЯ И ФАУНИСТИЧЕСКИЙ
ОБЗОР ХИРОНОМИД
В главе приводится подробная характеристика районов исследования, включая Волгоградское водохранилище (мелководье и русловую часть), пруды урбанизированной территории г. Саратова и Саратовской области (рис.1). Дана оценка трофического статуса районов исследования, степени загрязнения участков водохранилища и прудов (на основании собственных данных и материалов литературных и архивных источников) с использованием индекса ИЗВ. Все исследованные водные объекты по своему трофическому статусу относятся к эвтрофному типу; по степени загрязнения выделяются: «умеренно-загрязненные», «загрязненные» и «грязные». Представлены результаты многолетних исследований (1994-2002 гг.) динамики численности и биомассы сообществ макрозообентоса в различных экотопах Волгоградского водохранилища.
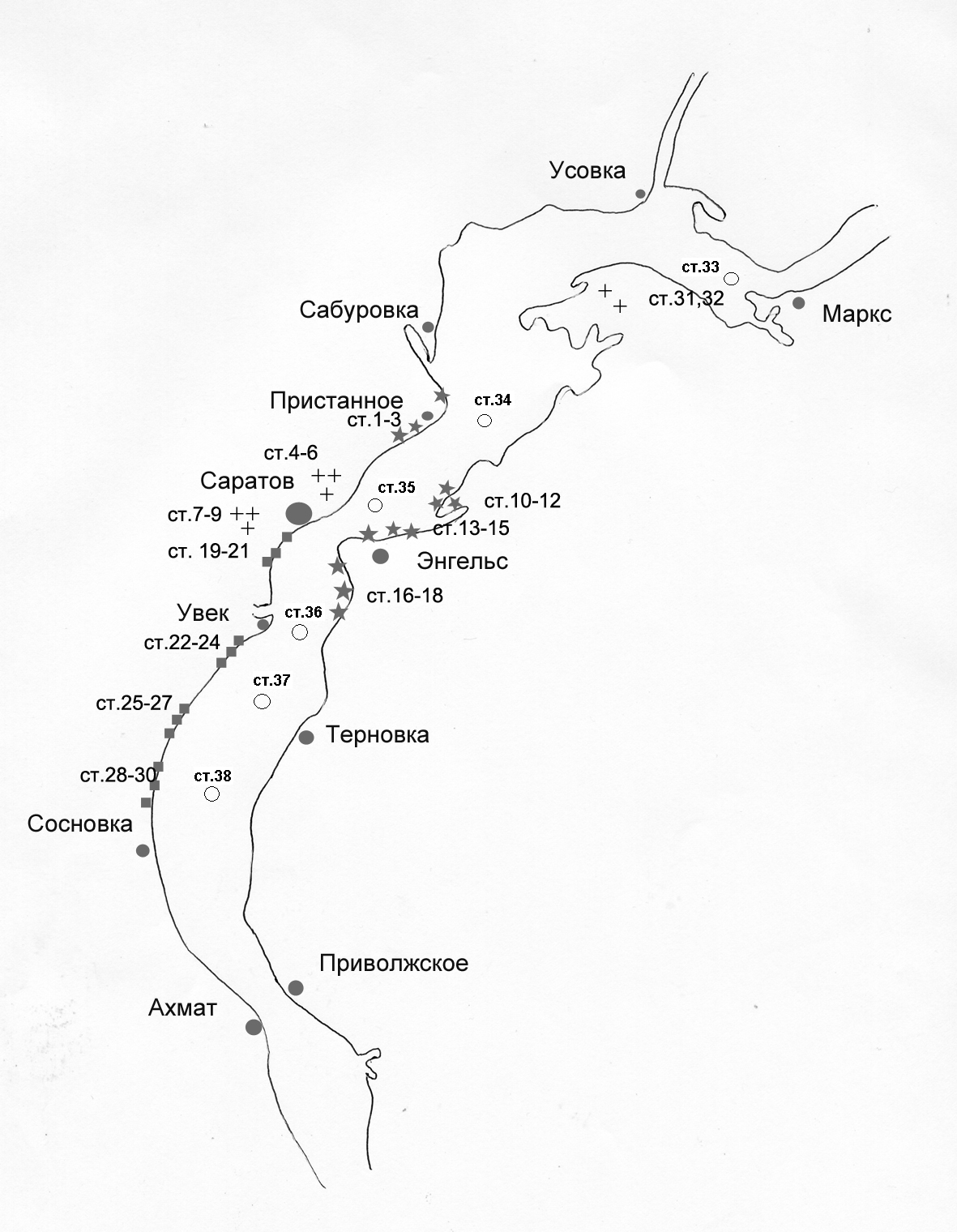
Рис. 1. Карта-схема Волгоградского водохранилища. Обозначения мест отбора проб:
 - защищенные мелководья; ■ - открытые мелководья; + - пруды; ○ – русло
- защищенные мелководья; ■ - открытые мелководья; + - пруды; ○ – руслоПриведены данные пространственного распределения сообществ макрозообентоса и хирономид. В многолетней динамике донных сообществ Волгоградского водохранилища в результате их трансформации, изменяется видовое разнообразие (для сообществ макрозообентоса соответственно Нn=1,2 -0,5 бит/экз.), преобладающими по биомассе становятся аборигенные олигохеты, инвазионных моллюски и ракообразные. Вселившиеся полихеты натурализовались и имеют достаточно высокую численность на мелководье и в русловой части (133-208 экз./м2). Доля хирономид в общей биомассе «мягкого» бентоса варьирует от 0,1 до 40% и составляет 11- 47% от общей численности бентоса. Биомасса хирономид изменяется в отдельные годы от 0,4 до 1,1 г/м2, при численности личинок – от 20 до 850 экз./м2 .
Фауна хирономид Волгоградского водохранилища включает 56 видов и личиночных групп (Мисейко, 1966; Нечваленко, 1974; Филинова, 2003). Ранее (Константинов, 1953) отмечалось значительное видовое разнообразие хирономид не зарегулированной р. Волга, после чего, при формировании водохранилища, выявлено заметное обеднение видового состава хирономид (Белявская, 1962, Мисейко, 1966, Белянина, Константинов, 1972; Нечваленко, 1977). Ведущими в ценозе хирономид по частоте встречаемости являлись - Chironomus plumosus и Chironomus annula-rius. На участках защищенного мелководья преобладали личинки Cladotanytarsus mancus, Parachironomus tener, Polipedilum bicrenatum, Рolipe-dilum nubeculosum, Procladius ferrugineus, С. gr. defectus (Филинова, 1986, 1991, 2001). Исследования мелководной и глубоководной зон водохранилища (летом 2002 г.) свидетельствуют о том, что наибольшим числом видов среди представителей макрозообентоса характеризуются хирономиды - 31 вид. Рассчитанные индексы доминирования для ценоза хирономид выявили структурные особенности комплекса, которые заключаются в формировании доминирующего ядра хирономид, с учетом их численности, представленного эврибионтными – Polypedilum nubeculosum (dn-21,0), Cladotanytarsus mancus (dn-14,1) и др. Субдоминантами по биомассе, наряду с другими видами хирономид, являются C. gr. defectus (dn-1,2; dв-3,1).
Глава 3. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объект исследования – сообщества макрозообентоса и ценоз хирономид р. Cryptochironomus. Основу исходного материала составили оригинальные данные, полученные в ходе многолетних исследований хирономид (1988-2007гг.) в разнотипных водоемах (см. рис. 1), отличающихся разнообразием биотопов, гидрохимическими и гидрологическими параметрами (Волгоградское водохранилище: районы защищенного и открытого мелководья; пруды).
Для выявления признаков надежной видовой диагностики при проведении морфологического анализа видов р. Cryptochironomus был собран с использованием общепринятых методов (Панкратова, 1970; Шилова, 1976; Шилова и др., 1982; Макарченко, 1985, 1999, 2006) многолетний материал, основу которого составили сборы имаго, куколок и личинок. Видовая идентификация хирономид проведена на основе материалов выведения имаго из личинок и куколок. Обработаны коллекционные материалы Зоологического института РАН и частных коллекций X. Vallenduuk, P. Langton.
Отбор количественных и качественных проб бентоса и их обработка проводились с использованием общепринятых методов гидробиологических исследований (Константинов, 1950; Методы, 1975; Руководство…, 1983). Анализ гидробиологического материала проведен с использованием архивных данных института экологии Волжского бассейна РАН и литературных сведений по многолетней динамике зообентоса Волгоградского водохранилища (Белявская, 1966, 1969; Нечваленко, 1976, 1980; Филинова, 2003; Шашуловский и др., 2005). Оценка качества воды и степени загрязнения водоемов проведена на основе гидрохимического анализа воды и грунта в аккредитованной лаборатории аналитической химии и химической экологии Саратовского государственного университета им. Н.Г. Чернышевского. Трофический статус исследуемых участков водохранилища и других водных объектов определялся по: И.С.Трифоновой (1990); M.Sakamoto (1966); B.W. Tayler et al. (1980).
Для интегральной характеристики сообществ бентоса и ценоза хирономид использовали показатели: численность (экз./м2 ), биомасса (г/ м2), число видов, частота встречаемости (%), индекс видового разнообразия Шеннона (Shannon, Weaver, 1949), индекс доминирования (Палий, 1961; Kownacki, 1971), биотический индекс Вудивисса (Woodowiiss, 1964; Вудивисс, 1977). Общий объем материала составил 800 количественных и 500 качественных проб (для отбора личинок и куколок хирономид) из водоемов разного типа. При проведении комплексного анализа видовой принадлежности хирономид р. Cryptochironomus исследовано свыше 17 тыс. личинок.
Для оценки биоиндикационной значимости видов хирономид р. Cryptochironomus, обитающих в условиях разных водоемов, были проведены многолетние мониторинговые исследования (1998-2003 гг.) на 9 станциях Волгоградского водохранилища и прудов. Рассматриваемые станции (экотопы) статистически значимо отличаются по показателям: БПК5, общей минерализации, ToC воды, содержанию кремния. В качестве «модельных» объектов выбраны виды C. obreptans и C. defectus. Объем обработанного материала представлен в табл. 1.
При морфологическом описании вида за основу были взяты общепринятые показатели и обозначения (Шилова, 1966; Saether, 1980). Для личинок введены новые отличительные признаки: индекс S3/A1 (расстояние между фронтальными щетинками S3, отнесенное к длине первого членика антенны); форма зубцов на коготках передних подталкивателей; форма и расположение бороздок на вентроментальных пластинках (для рода ранее не использовались). Морфологические особенности личинок C. obreptans и C. defectus IV возраста и степень их изменчивости изучались путем измерения 11 признаков: BL - длина тела (мм); CL - длина головной капсулы (мкм); BL/CL -отношение длины тела к длине головной капсулы; BL/CW - отношение длины тела к ширине головной капсулы; CL/CW - отношение длины головной капсулы к ее ширине; CW - ширина головной капсулы (мкм); OW - ширина затылочного отверстия (мкм); CW/OW - отношение ширины головной капсулы к ширине затылочного отверстия; MW - ширина ментума (мкм); AL-длина антенны (мкм); LMО - расстояние от затылочного склерита до вершины срединного зубца ментума (мкм). Статистическая обработка морфологических признаков проведена общепринятыми методами (среднеарифметические значения признака, дисперсия, среднеквадратическое отклонение, стандартная ошибка, достоверность отличий по критерию Стьюдента, коэффициент корреляции).
Таблица 1
Характеристика станций мониторинговых исследований видов
( C. obreptans и C. defectus) и объем обработанного материала
| Водоем | Вид | Морфологический анализ, n | Кариотипический анализ, n | Анализ содержимого кишечника, n |
| Волгоградское водохранилище, участки защищенного мелководья: ст. 1,13,14 | C. obreptans | 2100 | 372 | 375 |
| C. defectus | 1350 | 349 | 375 | |
| Волгоградское водохранилище, участки открытого мелководья: ст. 25,28,29 | C. obreptans | 4100 | 402 | 600 |
| C. defectus | 1676 | 396 | 600 | |
| Пруды урбанизированной территории г. Саратова: ст. 7,8,9 | C. obreptans | 1350 | 383 | 225 |
| C. defectus | 1800 | 406 | 225 | |
| Всего (экз.) | | 12376 | 2308 | 2400 |
Проведен сравнительно-морфологический анализ видовых признаков личинок, куколок и имаго самцов 18 видов р. Cryptochironomus, а так-же двух представителей нового подрода Miscellanea - С. (Miscellanea) dilatatus (Zorina, 2000), C.(M.) rectus (Zorina, 2000) и вида Demicryptochironomus vulneratus Zett. Родственные связи видов определяли с применением методов иерархического кластерного анализа (Айвазян и др., 1989; Компьютерная биометрика, 1990; Шитиков и др., 2003). В качестве меры сходства и различия исследуемых морфологических признаков применяли нормированные метрики: евклидово расстояние и его квадрат, манхеттенское расстояние. Дендрограммы родственных связей строили, используя ряд алгоритмов иерархического кластерного анализа: «средней связи», «медианной связи», «ближайшего соседа», «дальнего соседа», а также алгоритмы Уорда и центроидный. При использовании указанных метрик и алгоритмов объединения получали серии дендрограмм, которые анализировались по степени сходства. Решение о близости видов основывали на тех вариантах дендрограмм, которые дублировались наибольшим количеством используемых сочетаний метрик и алгоритмов.
Кариотипическое изучение видов р. Cryptochironomus проводили на препаратах политенных хромосом слюнных желез личинок, приготовленных по стандартной ацет-орсеиновой методике (Кикнадзе и др., 1991). При изучении генетической структуры популяций C. obreptans и C. defectus оценивались процент личинок с гетерозиготными инверсиями на популяцию (число изученных личинок из одной фиксации) и среднее число гетерозиготных инверсий на особь.
При выяснении числа генераций учитывали сезонные изменения численности личинок на единицу площади (экз./м2) и анализировали возрастной состав их популяций с учетом фаз развития личинок IV возраста (Ильинская, Иордан, 1975, 1977). На основе размерных характеристик личинок C. obreptans и C. defectus III и IV возраста построены гистограммы размерных спектров популяций. Питание личинок р. Cryptochironomus изучалось путем анализа содержимого их кишечников (Константинов, 1958, 1961; Луферов, 1961; Извекова, 1975, 1980; Шилова, 1976).
С целью установления временной и пространственной динамики численности популяций хирономид C. obreptans и C. defectus проведен двухфакторный дисперсионный анализ полученных данных (Шеффе, 1980; Шитиков и др., 2003). В ходе анализа статистическая достоверность долей дисперсии оценивалась с помощью F-критерия Фишера по уровню вероятности ошибочной оценки Р, который не должен превышать 0,05 (при принятом уровне значимости 5%). Проверку гипотезы о равенстве групповых средних осуществляли методом множественных сравнений (Шеффе, 1980). Для оценки нормальности распределения использовали W-критерий Шапиро-Уилка (для малых выборок). Дополнительно для оценки однородности выборок применялся непараметрический критерий Краскела-Уоллиса (Шитиков и др., 2003). Обработку материала проводили с использованием прикладных статистических программ (Компьютерная биометрика, 1990; Тюрин, Макаров, 1995).
Многолетняя динамика численности популяций C. obreptans и C. defectus рассмотрена в периоды: 1988-1992, 1993-1997, 1998-2003 гг. Пространственные изменения количественных показателей хирономид анализировались в условиях различных экотопов: защищенного (1), открытого (2) мелководий водохранилища и прудов (3). Для формализации индикаторной значимости видов использовали классический вариант дискриминантного анализа, основанный на определении канонических направлений в исходном пространстве признаков, удовлетворяющих критерию максимума отношения межгрупповой к внутригрупповой дисперсии (Айвазян и др., 1989; Шитиков и др., 2003). Группирующим фактором, на основе которого происходила классификация данных по численности хирономид, служила принадлежность к тому или иному экотопу.
Для изучения взаимосвязи численности рассматриваемых видов хирономид с основными параметрами среды был проведен факторный анализ по методу главных компонент (ГК) (Лиепа, 1980; Айвазян и др., 1989; Коросов, 1996). К рассмотрению принимались ГК (факторы), дисперсии которых были больше единицы (Иберла, 1980). Для получения окончательной матрицы факторных нагрузок использовали процедуру вращения осей координат факторного пространства методом «варимакс». Для оценки возможного комплексного влияния нескольких факторов на численность хирономид был проведен множественный регрессионный анализ. Отбор наиболее значимых предикторов осуществляли методом пошагового исключения (Дрейпер, Смит, 1987; Уланова, Забелин, 1990).
В целях выявления индикаторных свойств хирономид, с помощью дисперсионного анализа (Шеффе, 1980; Шитиков и др., 2003) проведена оценка различий численности личинок хирономид C. obreptans и C. defectus в водах разного уровня загрязнения.
Глава 4. МОРФОЛОГИЯ И СИСТЕМАТИКА ВИДОВ
ХИРОНОМИД РОДА CRYPTOCHIRONOMUS
4.1. Сравнительный морфологический анализ видов
хирономид рода Cryptochironomus
Среди представителей р. Cryptochironomus выделяют группы видов: Cryptochironomus gr. defectus и Cryptochironomus gr. fulvus (Шилова, 1966; Beck and Becк, 1964; Saether, 1977; Mason, 1985). Существование двух групп близких видов, обитающих в различных географических районах, обусловливает поиск информативных морфологических признаков, имеющих диагностическую ценность в пределах рода.
Сравнительно-морфологический анализ серии дендрограмм, составленных по морфологическим признакам личинок, куколок и имаго 20 видов р. Cryptochironomus и вида D. vulneratus, не выявил четкого разделения рода на две группы C. gr. defectus и C. gr. fulvus. (рис. 2,3,4). При этом имаго представителей нового подрода C. (Miscellanea) dilatatus, С. (Miscellanea) rectus и вида D. vulneratus образуют на анализируемых дендрограммах обособленную ветвь (рис. 4).
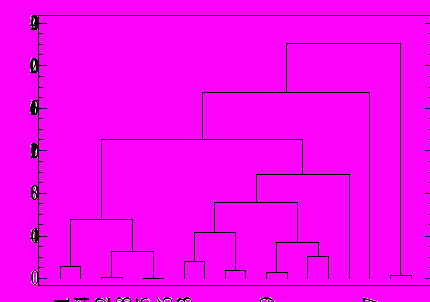
Рис. 2. Дендрограмма: личинки, алгоритм Уорда, по оси ординат – квадрат Евклидова расстояния: 1 - С. defectus; 2 - С. оbreptans; 3 - С. rostratus; 4 - С. supplicans; 5 - С. redekei; 6 - С. psittacinus; 7 - С. ussouriensis; 8 - С. andreevi; 9 - C.sorex; 10 - C. fulvus; 11- C.blarina; 12 - С. digitatus, 13 - C. conus,14 - C.curryi, 15 - C. eminentia, 16 - C. ramus, 17 - D. vulneratus, 18 - С. sp.А

Рис. 3. Дендрограмма: куколки, алгоритм Уорда, по оси ординат – квадрат Евклидова расстояния: 1 - С. defectus; 2 - С. оbreptans; 3 - С. rostratus; 4 - С. supplicans; 5 - С. redekei; 6 - С. psittacinus; 7 - С. ussouriensis; 8 - С. andreevi; 9 - C.sorex; 10 - C. fulvus; 11- C.blarina; 12 - С. digitatus, 13 - C. conus,14 - C.curryi, 15 - C. eminentia, 16 - C. ramus, 17 - D. vulneratus

Рис. 4. Дендрограмма: имаго, алгоритм Уорда, по оси ординат – квадрат Евклидова расстояния: 1 - С. defectus; 2 - С. оbreptans; 3 - С. rostratus; 4 - С. supplicans; 5 - С. redekei; 6 - С. psittacinus; 7 - С. ussouriensis; 8 - С. andreevi; 9 - C.sorex; 10 - C. fulvus; 11- C.blarina; 12 - С. digitatus, 13 - C. conus,14 - C.curryi, 15 - C. eminentia, 16 - C. ramus, 17 - D. vulneratus; 19 - С. (M) dilatatus ; 20 - C.(M.) rectus; 21- С. tamayoroi
Анализ морфологии 20 палеарктических и неарктических видов рода Cryptochironomus позволяет заключить, что для дифференциации личинок могут быть использованы следующие признаки: форма и окраска срединного зубца ментума; количество боковых зубцов ментума; количество зубцов премандибулы; особенности строения антенн и мандибул; пигментация гулярного склерита; для куколок – количество и форма оральных рожков, наличие сетчатости на поверхности брюшных сегментов; особенности строения органа дыхания; степень выраженности боковых выростов у задних углов II брюшного сегмента; расстояние между дугами поперечного ряда крючков на II тергите, форма выроста между анальными лопостями; для имаго – наличие фронтальных бугорков, особенности расположения, форма и опушение придатков гипопигия, форма гоностиля, наличие киля, окраска тела.
4.2. Морфологические особенности палеарктических видов
хирономид рода Cryptochironomus
С учетом литературных сведений (Шилова,1959, 1966, 1969; Родова, 1978; Панкратова, 1983; Кикнадзе и др., 1991; Kieffer, 1918; Lenz, 1926, 1941; Harnisch, 1923; Edwards, 1929; Goetghebuer, 1937; Saether, 1977, 1980; Pinder, 1978; Pinder, Reiss, 1983, 1986; Granston et al., 1989; Langton, 1991, 1994; Saether et al., 2000) и собственных данных представлен обобщенный диагноз р. Cryptochironomus на стадии личинки, куколки и имаго. Проанализированы морфологические характеристики 11 палеарктических видов данного рода (Шилова, 1966; Родова, 1978; Панкратова, 1983; Reiss, 1968; Pinder, 1978; Langton, 1991, 1994). На основе разработанной системы диагностических признаков составлены морфологические описания личинок, куколок и имаго С. defectus; С. obreptans; С. psittacinus; С. redekei; С. rostratus; С. supplicans; С. ussouriensis; C. andreevi. Представлены определительные таблицы личинок и куколок р. Cryptochironomus фауны Палеарктики, в которых придерживались трактовки видов C. redekei, C. psittacinus по А.И. Шиловой (1966), видов C. supplicans, C. аlbofasciatus по П. Лэнгтону (Langton,1991, 1994).
Определительная таблица личинок рода Cryptochironomus
IV возраста
1. Срединный зубец ментума светлый. Последние боковые зубцы ментума расположены выше первых боковых зубцов. Срединный зубец ментума без надрезов по бокам (рис. 5).……..……... ………………. 2
- Срединный зубец ментума темный. Последние боковые зубцы ментума расположены ниже первых боковых зубцов. Срединный зубец ментума с надрезами по бокам…… С. ussouriensis (Goetghebuer, 1933)
Вид известен только для России.
2. Гулярный склерит пигментирован. Пятно на гулярном склерите может быть большим и занимать более половины длины гулярного склерита, может быть слабо пигментировано. Основание затылочного склерита пигментировано также как вершинный зубец мандибулы….…...3
- Гулярный склерит светлый, не пигментирован. Основание затылочного склерита светло- или темно-желтое…...………………….…………4
3. …..……………………………………….… С. obreptans и С. supplicans
Виды трудно различимы по личинке. Для дифференциации видов можно использовать кариотипические данные и учитывать следующие морфологические параметры:
- S3-S3 (рис. 6) 45-65 мкм; индекс S3/A1 (отношение S3-S3 к длине
первого членика антенны) 0,63 -0,79. Зубцы на коготках передних подталкивателей тупые …….…...……..… С. obreptans (Walker, 1856)
б. S3-S3 60-80 мкм; индекс S3/A1 0,76-0,90; Зубцы на коготках
передних подталкивателей острые………С. supplicans (Meigen, 1830)
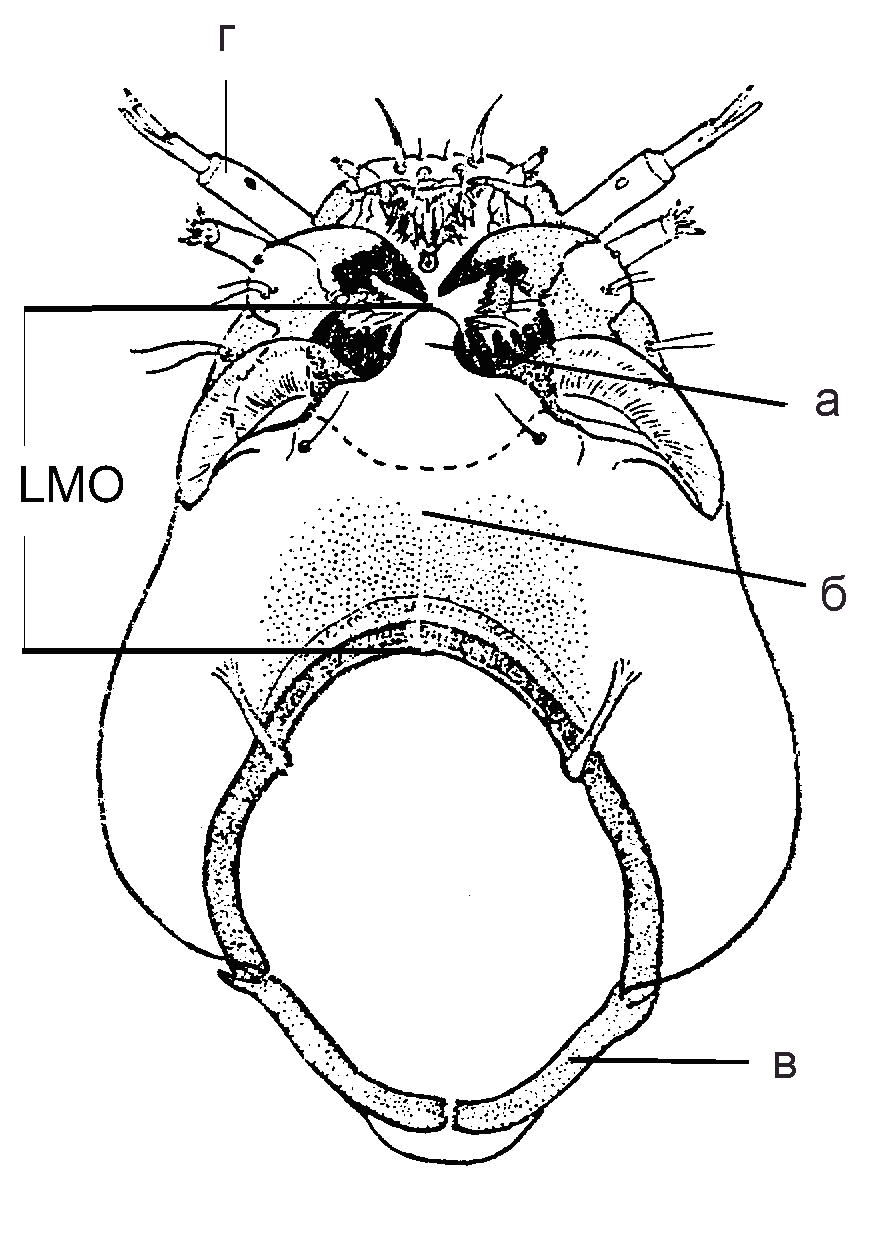 | 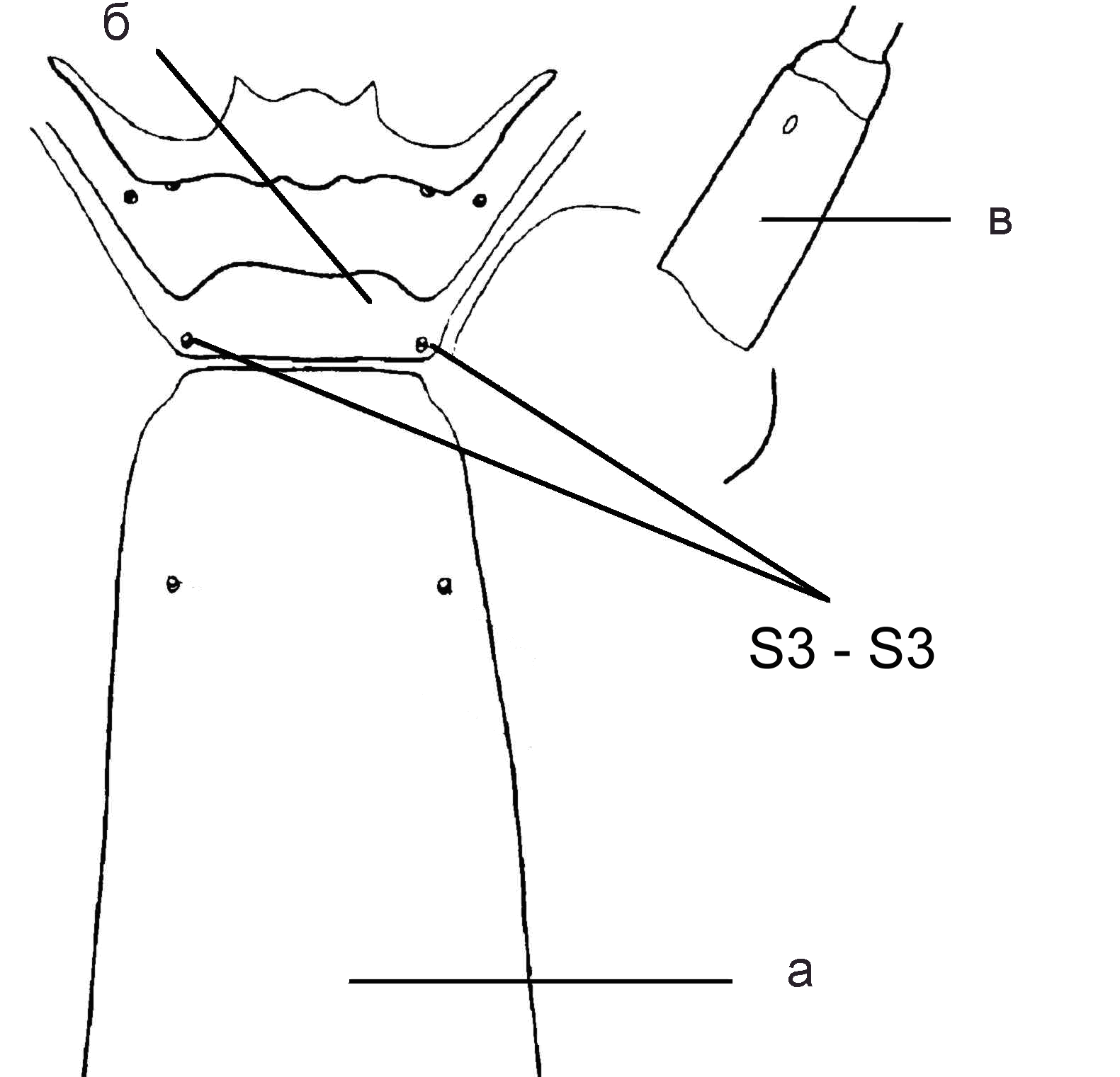 |
| Рис 5. Cryptochironomus obreptans. Голова, вентральная сторона: а – мен-тум; б - гулярный склерит; в - затылочный склерит; г – антенна (по: Кикнадзе и др., 1991) | Рис. 6. Cryptochironomus obreptans. Голова, дорзальная сторона: а – фронталь-ный склерит; б - латеральный склерит 1; в – антенна, первый сегмент |
4. Длина головы 395- 635 мкм. Ширина головы 265-435 мкм .…..….......5
- Длина головы 800-925 мкм. Ширина головы 530-720 мкм ........….…..7
5. LMO 180-240 мкм (рис.5). Длина вентроментальных пластинок 45-63 мкм. Длина головы 475-635 мкм. Бороздки на вентроментальных пластинках почти достигают края пластинок………....С. defectus (Kieffer, 1913)
- LMO 135-170 мкм. Длина вентроментальных пластинок 35-50 мкм. Длина головы 395-460 мкм. Бороздки на вентроментальных пластинках составляют только ¾ их длины ……….………….….…………... 6
6. Четвертый боковой зубец ментума выше пятого зубца. LMO 135-170мкм. Бороздки вентроментальных пластинок немного перекрывают друг друга. Длина головы 395-460 мкм. Ширина головы 265-325 мкм. Второй и третий сегмент антенны приблизительно равны. Щетинка SSd достигает верхнего зубца мандибулы ……………………..
…………………………………………………..С. rostratus Kieffer, 1921
- Четвертый боковой зубец ментума по высоте почти равен пятому зубцу. LMO 150 мкм. Бороздки вентроментальных пластинок крупные и относительно широко разделены друг от друга; некоторые линии бороздок могут сходиться в одну линию. Длина головы неизвестна. Ширина головы 380 мкм. Второй сегмент антенны короче, чем третий сегмент. Щетинка SSd достигает второго нижнего зубца мандибулы (вид редкий)… ………..….С. denticulatus (Goetghebuer, 1921)
7. Затылочный склерит светлый или темно-желтый. Длина головы 800-925 мкм. LMO 330-360 мкм. Длина вентроментальных пластинок 65-75 мкм. Длина мандибулы 225-270 мкм индекс S3/A1 1,05-1,2 ..……..
….……….... С. psittacinus (Meigen, 1830) и С. redekei (Kruseman, 1933)
Виды трудно различимы по личинке. Для дифференциации видов необходимо использовать кариотипические данные.
- Затылочный склерит коричневый как верхний зубец мандибулы.
Длина головы 600-820 мкм. LMO 238-250 мкм. Длина вентроменталь-
ных пластинок 55-65 мкм. Длина мандибулы - 215 мкм, индекс S3/A1
0,71-0,86 …………………………...…….... C. andreevi, Morozova sp.n.
Определительная таблица для диагностики личинок рода
Cryptochironomus и определения их возраста (III или IV):
1. Гулярный склерит пигментирован …..………………………………. 2
- Гулярный склерит без пигментации …..………………………...…....3
2. Длина тела 6,0 мм и более. Длина головы 480 мкм и более, ширина - 360 мкм и более…..……………………….... С. obreptans, возраст IV
- Длина тела 6,0 мм и менее. Длина головы 480 мкм и менее, ширина - 360 мкм и менее……………………………..С. obreptans, возраст III
3. Длина тела 6,5 мм и более. Длина головы 400 мкм и более, ширина головы 300 мкм и более………………….......... С. defectus, возраст IV
- Длина тела 6,5 мм и менее. Длина головы 400 мкм и менее, ширина головы 300 мкм и менее ………………………..С. defectus, возраст III
Определительная таблица куколок рода Cryptochironomus
1. Поперечный ряд крючков на II тергите прерван. Вырост между анальными лопастями фуркообразно разветвлен. В органе дыхания можно выделить основной ствол и 3 - 4 дополнительных пучка серебристых нитей…...………..………………………………...………...2
- Поперечный ряд крючков на II тергите сплошной. Вырост между анальными лопастями треугольный. В органе дыхания четко выделяются основной ствол и дополнительный пучок, разветвленный на 10 – 14 серебристых нитей ..…….С. ussouriensis (Goetghebuer, 1933)
2. Брюшные сегменты без сетчатой структуры. Следы сетчатой струк-
туры могут едва различаться на передних сегментах экзувиев кукол-
ки……….…………………………..………………………………….....3
- I - V брюшные сегменты с ясной сетчатой структурой, стенки которой тонкие или толстые……………...……………………………...…6
3. Головогрудь с передними вентральными боковыми выростами …..………..................................................................................................4
- Головогрудь с передними дорсальными и вентральными боковыми выростами ….………….……...……...…… С. obreptans (Walker, 1856)
4. Боковые выросты у задних углов II брюшного сегмента хорошо развиты. Грануляция головогруди мелкая, не более 8 мкм…….. …………………..………………….................С. defectus (Kieffer, 1913)
- Боковые выросты у задних углов II брюшного сегмента не развиты. Грануляция головогруди крупная, 10 мкм и более……………...…...5
5. Головные бугорки прямые, их длина 180-255мкм. Расстояние меж-
ду дугами поперечного ряда крючков на II тергите немного больше
или равно длине одной дуги. Грануляция передней части головогру-
ди плотная и образует кольцо на антипронотуме …………………….
…….………...………………….............. С. albofasciatus (Staeger, 1839)
- Головные бугорки загнуты, их длина 275-515. Расстояние между дугами поперечного ряда крючков на II тергите много меньше, чем длина одной дуги. Грануляция передней части головогруди менее плотная……..… …..………….………..….С. supplicans (Meigen, 1830)
6. Стенки сетчатой структуры брюшных сегментов толстые (рис.11). Боковые выросты у задних углов II брюшного сегмента, если выражены, то без щетинок ……..…...……………...……………………….7
- Стенки сетчатой структуры брюшных сегментов тонкие. Боковые выросты у задних углов II брюшного сегмента несут щетинки. Расстояние между дугами поперечного ряда крючков на II тергите равно длине одной дуги. Сетчатая структура развита на I-VI сегментах, менее ясная на VII и на VIII, с небольшими шипиками………. ……..………………………..……..С. denticulatus (Goetghebuer, 1921)
7. Боковые выросты у задних углов II брюшного сегмента хорошо выражены. Расстояние между дугами поперечного ряда крючков на II тергите равно или больше длины одной дуги ..……..…………....8
- Боковые выросты у задних углов II брюшного сегмента отсутствуют. Расстояние между дугами поперечного ряда крючков на II тергите много меньше длины одной дуги ……………….…...………....9
8. Поперечные ряды шипов у задних краев I-VII тергитов желто-коричневые. Стенки сетчатой структуры без шипиков ……………. …..……..………………….……..…………....С. rostratus Kieffer, 1921
- Поперечные ряды шипов у задних краев I-VII тергитов темно-коричневые, почти черные. Стенки сетчатой структуры с шипиками. Спинные выросты головогруди хорошо развиты ....…………….. ….………………………………......С. crassiforceps Goetghebuer, 1931