
Эколого-морфологический анализ хирономид рода cryptochironomus kieffer (diptera, chironomidae) палеарктики 03. 00. 16 экология 03. 00. 09 энтомология
| Вид материала | Автореферат диссертации |
- Хирономиды (diptera, chironomidae) бассейна нижнего амура. Фауна, систематика, распространение, 387.83kb.
- Общая характеристика работы актуальность, 882.91kb.
- Эколого-фаунистический анализ паразитофауны рыб вислинского залива, рек преголи и прохладной, 419.14kb.
- Экология бактерий рода Rhodococcus из глубоководных битумных построек озера Байкал, 306.35kb.
- Тема. Медицинская энтомология, 52.97kb.
- Фауна, биология и экология рода anopheles mg. (Сем. Culicidae) в прибрежных экосистемах, 1214.31kb.
- Региональные программы по решению эколого-экономических задач. Эколого-экономические, 68.04kb.
- Рабочая программа на 2007-2008 учебный год по дисциплине: «энтомология» Специальность:, 172.45kb.
- Характер изменения функционально активных участков и компактности политенных хромосом, 364.37kb.
- Программа «Экология комнатных растений», 44.17kb.
Использование критерия Шеффе позволяет утверждать, что средние значения численности C. defectus в третьем периоде (1998-2003 гг.) с 95%-ной вероятностью отличаются от аналогичных показателей в первые два периода многолетних исследований. Это подтверждается и графическим анализом средних значений и их доверительных интервалов (рис. 12).
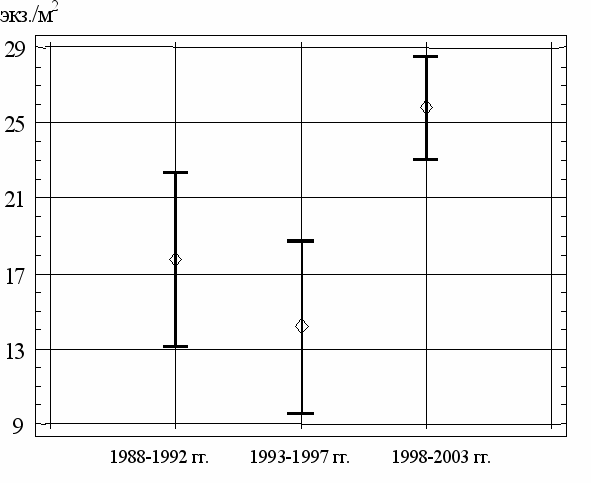
Рис. 12. Многолетние среднесезонные показатели численности (средние
значения и их 95%-й доверительный интервал) C. defectus в разные периоды
Влияние особенностей экотопов на численность C. defectus (см. табл. 6) проявляется в снижении плотности гидробионтов на грунтах открытых мелководий водохранилища (рис. 13). Популяции хирономид на участках защищенных мелководий и в прудах статистически не отличаются по чис-ленности. Неоднородность выборок подтверждается непараметрическим критерием Краскела-Уоллиса: для выборок по периодам KU=11,10 (Р=0,004) и экотопам - KU=48,43 (Р=3,04*10-11).
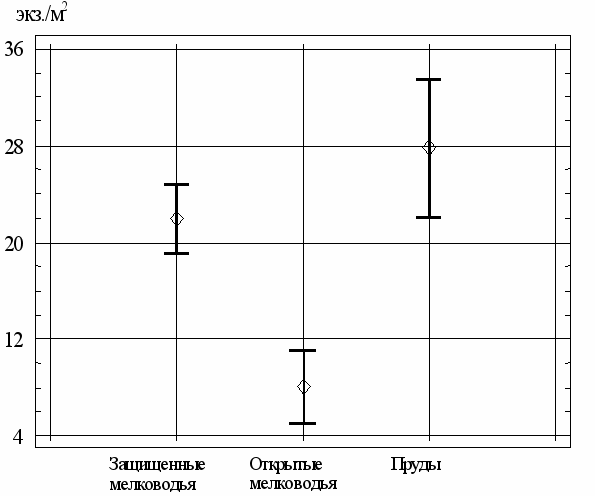
Рис. 13. Многолетние среднесезонные показатели численности (средние
значения и их 95%-й доверительный интервал) C. defectus в разных экотопах
Результаты дисперсионного анализа численности личинок двух видов хирономид свидетельствуют о том, что популяция C. obreptans поддерживает максимальную численность на открытых и защищенных мелководьях водохранилища, тогда как показатели численности популяции C. defectus максимальны в прудах и на участках защищенного мелководья водохранилища.
Одной из составных частей при оценке экологического состояния водоемов является изучение видового состава гидробиоценозов как наиболее важной биоиндикационной характеристики, которая непосредственно определяется условиями окружающей среды и существенно влияет на все остальные количественные ценотические показатели. Для обоснования различных подходов к оценке биоиндикационной значимости различных видов хирономид была проведена сравнительная оценка роли видов р. Cryptochironomus - C. obreptans и C. defectus как биоиндикаторов разнотипных водных объектов.
Факторный анализ по методу главных компонент (ГК) (Лиепа, 1980; Айвазян и др., 1989) позволил: 1) структурировать изученные показатели (численность C. defectus и C. obreptans, численность и биомасса бентоса, температура воды (июль), значение рН, содержание растворенного кислорода, БПК5, перманганатная и бихроматная окисляемость, общая минерализация, концентрации растворенного общего азота и фосфора, железа, кремния) на основе их максимальных взаимосвязей; 2) выделить наиболее важные из них, т.е. ГК (факторы); 3) оценить удельный вес изменчивости, приписываемой каждому из выделенных главных факторов; 4) количественно оценить взаимосвязь исходных показателей в рамках кластера того или иного главного фактора. Первый этап анализа выявил, что 5 выделенных ГК имеют собственные числа выше единицы и заключают в сумме 87,9% дисперсии исходных параметров. В этой связи необходимо отметить большую вариабельность массива рассматриваемых показателей, поскольку обычным считается (Коросов, 1996), что основная доля дисперсии исходных признаков должна распределяться между 2-3-мя (реже 4-мя). Высокая вариабельность в нашем случае объясняется охватом разнотипных групп водоемов и соответствующих им абиотических и биотических параметров.
В дальнейшем получена матрица факторных нагрузок (табл. 7), величины которых можно интерпретировать как коэффициенты корреля-ционной связи исходных параметров с выделенными главными факторами. Абсолютное значение нагрузки выше 0,7 принимается за существенную связь, ниже 0,7 – предполагается недоказанность связи (Джефферс, 1981). Наличие существенной связи указывает на согласованное изменение исходных параметров в рамках того или иного главного фактора.
Таблица 7
Факторные нагрузки исходных показателей для выделенных
главных факторов
| Показатели | Факторы | ||||
| 1 | 2 | 3 | 4 | 5 | |
| Численность C.defectus | -0,357 | 0,701 | -0,285 | -0,363 | 0,082 |
| Численность C.obreptans | 0,087 | 0,127 | -0,079 | -0,936 | -0,111 |
| Общая численность бентоса | 0,922 | -0,122 | 0,280 | 0,095 | -0,006 |
| Общая биомасса бентоса | 0,795 | -0,135 | 0,291 | 0,045 | -0,008 |
| Численность мягкого бентоса | 0,774 | 0,396 | 0,115 | 0,296 | 0,300 |
| Биомасса мягкого бентоса | 0,701 | 0,450 | 0,258 | 0,292 | -0,052 |
| Температура воды (июль) | -0,283 | 0,850 | -0,095 | 0,229 | 0,245 |
| рН | 0,072 | 0,133 | -0,172 | 0,016 | 0,941 |
| Концентрация кислорода в воде | 0,902 | -0,066 | -0,064 | -0,055 | -0,351 |
| БПК5 | 0,944 | 0,011 | 0,214 | 0,023 | 0,008 |
| Перманганатная окисляемость | 0,296 | 0,053 | 0,881 | -0,119 | -0,118 |
| Бихроматная окисляемость | 0,279 | -0,108 | 0,894 | 0,196 | -0,072 |
| Общая минерализация | 0,952 | -0,120 | 0,161 | 0,060 | -0,026 |
| Общий азот | 0,487 | 0,770 | 0,175 | -0,043 | -0,041 |
| Общий фосфор | 0,415 | 0,269 | -0,055 | 0,724 | -0,118 |
| Железо | 0,768 | -0,024 | 0,287 | 0,099 | 0,103 |
| Кремний | 0,804 | 0,159 | 0,128 | 0,034 | 0,388 |
| Доля дисперсии, % | 47,9 | 15,8 | 9,8 | 8,2 | 6,2 |
Примечание: жирным шрифтом выделены ГК, собственные числа которых более 1
В результате факторного анализа установлена связь динамики численности массовых хирономид с температурным режимом и биогенными элементами водоема: C. defectus -↔ ToC, Nобщ.; C. obreptans ↔ Pобщ. Выявленная связь обусловлена, возможно, увеличением нитратов, суммы минерального азота и кремния на различных участках Волгоградского водохранилища (Филинова, 2003), что является следствием непосредственного антропогенного воздействия на внутриводоемные биопродукционные процессы.
Для построения прогнозов изменения состояния экосистемы или «качества» окружающей среды является анализ причинно-следственных связей между эмпирическими индивидуальными и комплексными показателями. Для решения этой задачи используется регрессионный анализ. С целью оценки возможного комплексного влияния нескольких показателей на численность гидробионтов проведен множественный регрессионный анализ (Дрейпер, Смит, 1987; Уланова, Забелин, 1990). Значения коэффициентов в полученных регрессионных моделях приведены в табл. 8, из которой следует, что для изменения численности C. defectus статистически значимыми (р<0,05) являются 7 абиотических параметров, для C. obreptans – 4. Коэффициент множественной детерминации R2 модели для первого из видов составляет 0,915 (р=0,0001), для второго – 0,610 (р=0,011).
Таблица 8
Коэффициенты регрессионных уравнений зависимости показателей
численности C. defectus и C. obreptans от абиотических факторов
| C. defectus | C. obreptans | ||
| Показатели (Х1…Х7) | Коэффициенты | Показатели (Х1…Х4) | Коэффициенты |
| Температура воды (июль), оС | 17,75 | Температура воды (июль), оС | -10,85 |
| Содержание кислорода, мг О2/дм3 | 39,15 | Бихроматная окисляемость, мгО/дм3 | -36,88 |
| БПК5, мг О/дм3 | 117,15 | Общий азот, мг/дм3 | 65,72 |
| Общая минерализация, мг/дм3 | -0,548 | Общий фосфор,мг/дм3 | -734,13 |
| Общий азот, мг/дм3 | -33,00 | | |
| Общий фосфор, мг/дм3 | -490,79 | | |
| Железо, мг/дм3 | -127,34 | | |
| Свободный член | -871,1 | Свободный член | 571,8 |
Необходимо отметить, что в большинстве рассматриваемых гидрохимических параметрах существует как природная, так и антропогенная составляющая (исключением, по-видимому, являются кислородный режим и кремний), в результате чего выделить долю влияния последней на количественную составляющую гидробионтов сложно. Поэтому индикаторную значимость хирономид, более вероятно, рассматривать с точки зрения общего вектора динамики экосистем (например, эвтрофирования), а не конкретного влияния того или иного антропогенного фактора. В связи с чем, проведен анализ многолетних среднесезонных показателей численности популяций C. obreptans и C. defectus в соответствии с комплексом природных условий водоемов. Как следует из результатов дискриминантного анализа (табл. 9), уровень значимости позволяет признать статистическую состоятельность дискриминации данных по группирующему фактору.
Таблица 9
Результаты дискриминантного анализа показателей численности личинок двух видов хирономид по принадлежности к экотопу
| Основные характеристики | |||
| Дискриминантная функция | Собственные значения | Вклад в дисперсию, % | Канонические корреляции |
| 1 | 3,751 | 98,39 | 0,889 |
| 2 | 0,061 | 1,61 | 0,240 |
| Оценки уровня значимости | |||
| | Критерий Лямбда | Хи-квадрат | Р-значение |
| 1 | 0,1984 | 182,002 | <0,0001 |
| 2 | 0,9423 | 6,687 | 0,0097 |
При этом для решения классификационной задачи достаточно применение лишь одной первой дискриминантной функции, объясняющей более 98% дисперсии показателей численности. Графическое отображение результатов анализа представлено в виде диаграммы рассеивания наблюдаемых объектов на плоскости дискриминантных функций (рис. 14), из которой видно, что исследуемый ценоз донных организмов лимнических систем (пруды) образует достаточно самостоятельную группировку, практически не пересекающуюся с двумя другими классами. В то же время, ценозы мелководных участков водохранилища имеют область существенного взаимного перекрывания.
Информативность каждого из видов хирономид для дискриминации может быть оценена при рассмотрении стандартизированных коэффициентов первой дискриминантной функции. Для C. obreptans коэффициент равен –1,028, для C. defectus составляет 1,012. Близкие абсолютные значения коэффициентов указывают на сходную информативность численности двух видов для классификации по экотопам; индикаторная значимость видов приблизительно одинакова.
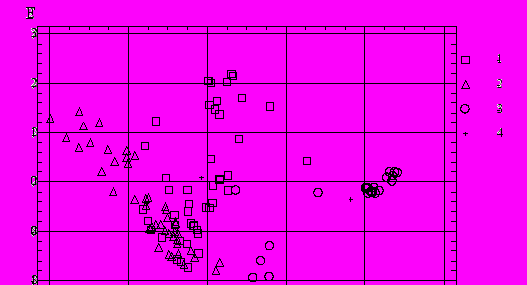
Рис. 14. Диаграмма рассеивания объектов на плоскости
двух дискриминантных функций: 1 – защищенные мелководья,
2 – открытые мелководья. 3 – пруды, 4 - центроиды классов
Полученная на основе рассматриваемой выборки табл. 10 результатов классификации исследованных экотопов свидетельствует о том, что корректная классификация проведена в целом для 81% наблюдений. Наиболее высока доля (93%) правильного выделения экотопа открытых мелководий, далее следуют экотопы прудов (83%) и защищенных мелководий (69%).
Таблица 10
Результаты классификации по экотопам, полученные
на основе обучающей выборки
| Фактический экотоп | Размер выборки | Прогнозируемый экотоп | Правильный прогноз, % | ||
| Защищенные мелководья | Открытые мелководья | Пруды | |||
| Защищенные мелководья | 48 | 33 | 14 | 1 | 68,8 |
| Открытые мелководья | 44 | 3 | 41 | - | 93,2 |
| Пруды | 24 | 4 | - | 20 | 83,3 |
| Всего | | | | | 81,0 |
Поскольку статистическая состоятельность дискриминации на обучающей выборке доказана, процентные значения корректной классификации могут быть интерпретированы как вероятностные оценки правильности прогноза по материалам вновь проводимых наблюдений. Прогнозная (индикаторная) модель, с помощью которой построена табл. 11, включает три линейных дискриминантных функции Фишера, параметры которых определены в рамках процедуры дискриминантного анализа. Каждая из этих функций определяет принадлежность наблюдения по численности личинок хирономид к одному из трех рассматриваемых экотопов. Порядок диагностики (индикации) биотопа с использованием классифицирующих функций заключается в том, чтобы на основе полученных данных по среднесезонной численности личинок двух видов хирономид рассчитать значения их откликов (d1, d2, d3). Наблюдение с определенной долей вероятности можно отнести к тому экотопу, для которого значение отклика классифицирующей функции будет максимальным.
Таблица 11
Прогнозная (индикаторная) модель
-
Экотоп
Классифицирующая функция
Защищенные мелководья -
d1 = -5,429 + 0,187Х1 + 0,025Х2
Открытые мелководья -
d2 = -6,857 + 0,264Х1 - 0,111Х2
Пруды -
d3 = -6,863 - 0,094Х1 + 0,311Х2,
Примечание: Х1 – среднесезонная численность C. obreptans, Х2 - среднесезонная численность C. defectus
Исследование многолетней динамики и закономерностей пространственного распределения сообществ макрозообентоса и доминирующих видов хирономид при их комплексном исследовании (систематика, морфология, биология, экология) являются в ряде случаев определяющими для выявления их индикаторной роли и оценки экологического состояния разнотипных водных объектов. В целях выявления индикаторных свойств гидробионтов проведена оценка различий показателей численности личинок хирономид в водах разного уровня загрязнения. Результаты анализа (табл. 12) показали, что изменения численности обоих видов в водах разного класса качества следует признать статистически достоверными (Р<0,001).
Таблица 12
Результаты дисперсионного анализа среднесезонных показателей численности
C. obreptans и C. defectus в водах разного класса качества
| Источник дисперсии | Сумма квадратов | Степень свободы | Средние квадраты | F-отноше-ние | Р-зна-чение |
| C.obreptans | |||||
| Между группами | 5291,7 | 2 | 2645,83 | 23,70 | <0,001 |
| Внутри групп | 3014,2 | 27 | 111,64 | | |
| Всего | 8305,9 | | | | |
| C.defectus | |||||
| Между группами | 5597,1 | 2 | 2798,53 | 13,49 | <0,001 |
| Внутри групп | 5600,4 | 27 | 207,42 | | |
| Всего | 11197,5 | | | | |
Вместе с тем, характер изменений численности для C. obreptans и C. defectus различен. Для C. obreptans весьма затруднительно говорить о наличии какой-либо тенденции (рис. 15), то в отношении C. defectus можно полагать, что с увеличением загрязненности вод численность гидробион-тов закономерно снижается (рис. 16, 17, 18).
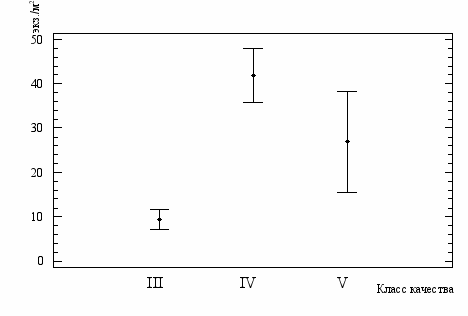
Рис.15. Среднесезонные показатели численность C.obreptans
в водах разного класса качества (средние значения и 95%-ный
доверительный интервал)
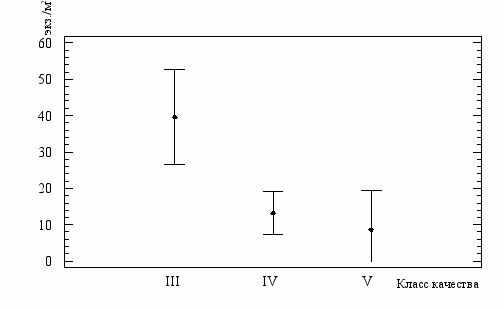
Рис.16. Среднесезонные показатели численности C.defectus
в водах разного класса качеств по ИЗВ (средние значения и
95%-ный доверительный интервал)
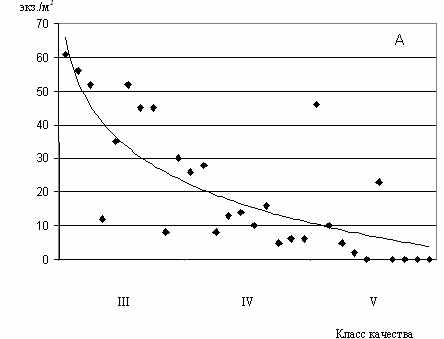
Рис.17. Зависимость среднесезонных показателей
численности C.defectus от класса качества вод
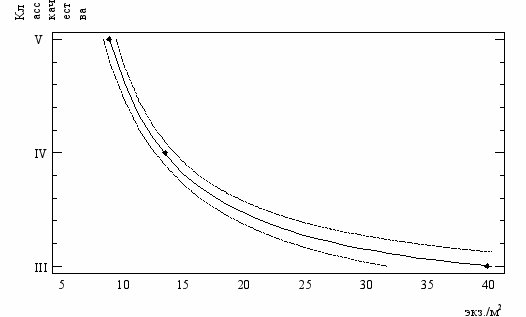
Рис.18. График регрессии класса качества вод по среднесезонным
показателям численности C. defectus (тонкими линиями обозначен
95%-ный доверительный интервал)
Формализация зависимости класса качества вод со средними значениями численности C. defectus, позволила получить регрессионное уравнение K = exp (0,958 + 5,615/Ndef), где K – цифровое обозначение класса качества воды, Ndef – среднесезонные показатели численности C. defectus экз./м2. При наличии всего трёх опорных точек, зависимость имеет коэффициент детерминации более 99% при уровне значимости Р=0,006.
Изменение соотношений видов при антропогенном воздействии определяется тем, что они по-разному реагируют на загрязнение. В настоящее время продолжается поиск видов-биоиндикаторов и обосновываются подходы к их изучению (Кожова, 1986; Манн, 1986; Щипанов, 1998; Криволуцкий и др., 1991; Баканов, 2000) с учетом знаний популяционной экологии видов и особенностей функционирования их популяций в среде обитания. По принципам функциональной организации популяций видов выделяют три группы: 1- способные изменять режим функционирования; 2 - способные обеспечить только восстанавливающую функцию; 3 - способные обеспечить контролирующую функцию. Виды 1-ой группы устойчивы в широком диапазоне колебаний качества среды, 2-ой - в динамической среде, а 3-ей - в стабильной (Щипанов, 1998, 2002). Результаты многолетнего анализа показателей численности видов C. obreptans и C. defectus говорят об изменчивости их функциональной организации и способности формировать группировки с различным типом функционирования в ответ на воздействие факторов среды.
ВЫВОДЫ
На основании комплексной оценки хирономид р. Cryptochironomus, включающей кариотипическое, морфологическое исследование палеарктических и неарктических видов, изучение биологических и экологических особенностей видов рода из водоемов бассейна Нижней Волги (на примере Волгоградского водохранилища и сопредельных водных объектов), можно сделать следующие выводы:
1. Для объективной диагностики личинок р. Cryptochironomus выделены следующие морфологические признаки: форма и окраска срединного зубца ментума, количество и расположение боковых зубцов ментума, количество зубцов премандибулы, длина и ширина головной капсулы, длина вентроментальных пластинок и мандибулы, индекс LMO, индекс S3/A1, форма зубцов на коготках передних подталкивателей, форма и расположение бороздок на вентроментальных пластинках, пигментация гулярного и затылочного склеритов; для куколок – количество и форма оральных рожек, наличие сетчатости на поверхности брюшных сегментов; особенности строения органа дыхания; степень выраженности боковых выростов II брюшного сегмента; расстояние между дугами поперечного ряда крючков на II тергите; количество выростов по бокам головогруди, форма выроста между анальными лопостями; для имаго – наличие фронтальных бугорков, особенности расположения, форма и опушение придатков гипопигия, форма гоностиля, наличие киля, окраска тела.
2. Установлены базовые кариотипические показатели для диагностики личинок р. Cryptochironomus: диплоидное число и морфологические отличия хромосом, степень морфологической выраженности центромерных районов, количество и расположение функционально активных районов хромосом.
3. Выявлена структурообразующая роль ценоза р. Cryptochironomus в сообществах макрозообентоса мелководной зоны водоемов бассейна Нижней Волги. Отмечена принадлежность видов р. Cryptochironomus к эвтрофным водоемам различной степени загрязнения.
4. Видовой состав хирономид р. Cryptochironomus в водоемах разного типа бассейна Нижней Волги включает: С. defectus (Kieffer, 1913); С. obreptans (Walker, 1856); С. psittacinus (Meigen, 1830); С. redekei (Kruseman, 1933); С. rostratus Kieffer, 1921; С. supplicans (Meigen, 1830); С. ussouriensis (Goetghebuer, 1933); С. andreevi, Morozova sp.n. Обнаружен многолетний (1988-2007 гг.) тренд снижения видового состава р. Cryptochironomus. Степень гетерогенности многолетней динамики видового состава р. Cryptochironomus на отдельных участках исследований неодинакова: наряду с массовыми (С. obreptans и С. defectus), в отдельные годы на мелководьях формируется ценоз с развитием видов C. supplicans, C. psittacinus, C. andreevi, C. rostratus, входящих в разряд второстепенных, сопутствующих массовым.
5. Массовые виды хирономид рода Cryptochironomus (С. obreptans и C. defectus), обитающие Волгоградском водохранилище и сопредельных водоемах, являются высоко полиморфным видам: 90,0-95,7% личинок С. obreptans и 56,0-70,8% личинок C. defectus из исследуемых популяций гетерозиготны по инверсиям. Наиболее высокие показатели хромосомного полиморфизм зарегистрированы в популяциях С. obreptans и C. defectus, обитающих в условиях широкого диапазона факторов среды литоральной зоны водохранилища.
6. В Волгоградском водохранилище и сопредельных водоемах виды С. obreptans, C. defectus, C. supplicans являются дицикличными, C. psittacinus, C. andreevi, C. rostratus C. redekei и С. ussouriensis имеют одну летнюю генерацию. Оценка многолетней динамики численности и биомассы хирономид и сообществ макрозообентоса в разнотипных водоемах при антропогенном воздействии показала, что вид С. obreptans развивает максимальную численность на участках защищенного и открытого мелководья водохранилища, С. defectus – в прудах и на защищенном мелководье. Вид C. obreptans способен поддерживать максимальную численность во временном масштабе при воздействии разных факторов среды, в то время как показатели численности C. defectus нестабильны во временном аспекте. Выявлено, что с увеличением загрязненности различных участков водохранилища численность C. defectus снижается.
7. Установлена связь динамики численности массовых видов хирономид с температурным режимом и биогенными элементами водных объектов: C. defectus -↔ ToC, Nобщ.; C. obreptans ↔ Pобщ. Изменения численности хирономид C. defectus обусловлено 7 статистически значимыми (р < 0,05) абиотическими параметрами (температура, общая минерализация воды, содержание растворенного кислорода, легкоокисляемое органическое вещество (БПК5), железо, азот, фосфор) , C. obreptans – 4 (температура воды, содержание трудноокисляемого органического вещества (ХПК), фосфор, азот).
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
* - публикации в изданиях перечня ВАК РФ
1. Белянина С.И. Логинова Н.В.(Полуконова), Морозова Е.Е. Фенотипические особенности хромосом двух видов хирономид Microtendipes pedellus и Fleuria lacustris из разных частей ареалов // Фенетика природных популяций. М.., 1990.С. 33-35.
2. Морозова Е.Е. Морфологические и кариотипические данные к диаг- ностике волжских видов Cryptochironomus gr. defectus Kieffer // Систематика, зоогеография и кариология двукрылых насекомых (Insecta: Diptera). Сб. научных работ. С.-Петербург, 1992.С.116-118.
3. Морозова Е.Е. Кариотипическая и морфологическая дифференциация видов Cryptochironomus gr. defectus Kieffer (Diptera, Chironomi-dae) // Кариосистематика беспозвоночных животных II. Сб. научных работ. С.- Петербург, 1993. С.71-75.
4*. Морозова Е.Е. Морфологические и кариотипические данные к диагностике волжских видов Cryptochironomus gr. defectus Kieffer (Dip-tera, Chironomidae) // Зоологический журнал. М., 1994. Т.73. Вып.3. С.72-82.
5. Морозова Е.Е. Кариотипическое и морфологическое изучение волжских видов Cryptochironomus ex gr. defectus Kieffer (Diptera, Chironomidae) // Кариосистематика беспозвоночных животных III. Сб. науч. работ. М.: Изд-во Ботанического сада Московского ун-та, 1996. С.49-51.
6. Morozova E.E. Diagnostic problems in species of Cryptochironomus ex. gr. defectus (Diptera:Chironomidae) from River Volga //Abstracts of 13 th International Symposium on Chironomidae in Freiburg, 5-9 September 1997. P.82.
7. Морозова Е.Е. Данные к экологии видов Cryptochironomus ex gr. defectus Kieffer (Diptera, Chironomidae) в районе Саратова // Место и роль двукрылых насекомых в экосистемах. Сб. науч. трудов VI Всерос. диптерологического симпозиума. С.-Петербург, 1997.С.83.
8. Морозова Е.Е. Видовая диагностика личинок волжских видов Crypto-chironomus ex gr. defectus Kieffer (Diptera, Chironomidae) // Проблемы энтомологии в России. Сб. науч. трудов XI Съезда Русского энтомологического общества. С.-Петербург, 1998. Том II. С.42-43.
9. Морозова Е.Е. Морфологическое и кариотипическое описание вида Cryptochironomus albofasciatus Staeg. (Chironomidae, Diptera) // Вопросы акмеологии. Межвуз. сб. науч. трудов. Саратов: Изд-во СГПИ, 1998. Вып. III. С.79-82.
10. Sergeeva I.V., Morozova E.E. Chironomidae from the Volga River // Chironomus, 1998. № 11. P.11-12.
11. Морозова Е.Е. История становления систематики рода Cryptochironomus Kieffer (1918) (Diptera, Chironomidae) // Вопросы акмеологии. Межвуз. сборник научных трудов. Саратов: Изд-во СГПИ, 1999. Вып.IV. С.161-165.
12. Морозова Е.Е. Взаимоотношения близкородственных видов Crypto-chironomus ex gr. defectus Kieffer (Diptera, Chironomidae): питание // Бюллетень Международной Академии психологических наук. Вопросы биологии. Межвуз. сборник научных работ. Саратов-Ярославль: Изд - во СГПИ, 1999. Вып.VII.С.163-166.
13. Белянина С.И., Сергеева И.В., Морозова Е.Е., Дурнова Н.А., Полуконова Н.В. Изменение видового состава некоторых групп хирономид Волги и придаточных водоемов в районе Саратова в связи с антропогенным воздействием // Гигиена окружающей среды и экология человека. Сб. науч. трудов Всерос. науч. конф. к 100-летию проф. Л. Л. Лося. Саратов: Изд-во СГМУ, 1999. С.13-17.
14. Морозова Е.Е. Видовой состав хирономид Cryptochironomus ex.gr. de-fectus (Chironomidae, Diptera) из Волги в районе Саратова // Стратегия природопользования и сохранения биоразнообразия в XXI веке. Сб. трудов Всерос. науч. конф. Оренбург: Изд-во ОГУ,1999. С.64-67.
15. Морозова Е.Е. Морфология Cryptochironomus defectus Kieffer (Diptera, Chironomidae) // Материалы Восьмых Страховских Чтений. Межвуз. сб. науч. трудов. Саратов: Изд-во СГПИ, 1999. С.50-54.
16*. Морозова Е.Е. Морфология самки Cryptochironomus defectus (Diptera, Chironomidae)//Зоологический журнал. М., 2000.Т.79.№7. С. 871-872.
17. Морозова Е.Е. Кариотипы трех волжских видов рода Cryptochironomus Kieffer (Diptera, Chironomidae)// Бюллетень Международной Академии психологических наук. Вопросы биологии. Межвуз. сб. науч. работ. Саратов: Изд-во СГПИ, 2000. Вып.VIII. С.259-262.
18. Морозова Е.Е. Экологические характеристики видов рода Cryptochi-ronomus Kieffer (Diptera, Chironomidae) // Вопросы акмеологии. Межвуз. сб. науч. работ. Саратов: Изд-во СГПИ, 2000. Вып.V. С.177-182.
19. Морозова Е.Е. Систематика и таксономия видов Cryptochironomus ex. gr. defectus (Diptera, Chironomidae) // Материалы Девятых Страховских Чтений. Межвуз. сб. науч. трудов. Саратов: Изд-во СГПИ, 2000. С.240-243.
20. Morozova E.E. Karyotype Features and Evolution of Cryptochironomus gen. species (Diptera, Chironomidae) // Biodiversity and dynamics of ecosystems in North Eurasia: Basic Problem of species and Ecosystemmmms Evolution.- Novosibirsk, 2000.Vol.1. Part.1. P.73-75.
21. Morozova E.E. The Volga region species of Cryptochironomus ex gr. defectus Kieffer (Diptera, Chironomidae): problems of diagnostics // Late 20 th Centure Research on Chironomidae: An antology from the 13 th International Symposium on Chironomidae (Freiburg, 5-8 September 1997). Fachen: Shaker, 2000. P. 193-196.
22. Шуйский В.Ф., Шувалов Ю.В., Перевозников М.А., Пашкевич М.А., Занцинская Т.П., Петров Д.С., Морозова Е.Е., Савченко А.В., Максимовская А.В., Маковский С.А., Петрова Т.А., Матюшонок М.Л. Сравнительная оценка аддитивной, мультипликативной и изоболической моделей многофакторных воздействий на биосистемы // Сб. науч. трудов ГосНИОРХ. С.-Петербург, 2000. Вып.326. С.118-130.
23. Морозова Е.Е. Виды рода Cryptochironomus Kieffer (Chironomidae, Dip-tera) как модельные объекты для познания закономерностей влияния условий среды на зообентос и биоту пресноводных водоемов // Бюллетень Международной Академии Психологических наук. Вопросы биологии. Межвуз. сб. науч. работ.Саратов-Ярославль: Изд-во СГПИ, 2000.Вып.9. С.256-262.