
Структурно-функциональные особенности запасных и защитных белков растений и их использование в генетических исследованиях
| Вид материала | Автореферат |
- Структурные и функциональные свойства цитокинин-связывающих белков растений Arabidopsis, 249.12kb.
- Педагогическая пресса в россии во второй половине XIX в.: Генезис, предметно-тематические, 302.96kb.
- Рабочая учебная программа по нормальной физиологии Электив «Структурно-функциональные, 112.81kb.
- Нп «сибирская ассоциация консультантов», 169.41kb.
- Календарно-тематический план практических занятий по биологии для студентов медико-биологического, 71.39kb.
- Названия экзотических растений в английском и русском языках (структурно-словообразовательный, 378.31kb.
- Программа 9 16 ноября 2010 г г. Москва Место проведения конференции: Учреждение Российской, 265.18kb.
- Добриева Зейнаб Израиловна структурно-семантические и функциональные особенности приставочных, 2630.63kb.
- Структурно-семантические и функциональные характеристики английского лингвокультурного, 562.68kb.
- Программа для поступающих в нввкус по специальности «Психология», 284.28kb.
Рис. 17. Аминокислотные последовательности некоторых дефензинов растений. †Последовательность установлена по структуре гена. Элементы вторичной структуры отмечены темно-серым (α-спирали) и серым (β-тяжи) (Lay, Anderson, 2005).
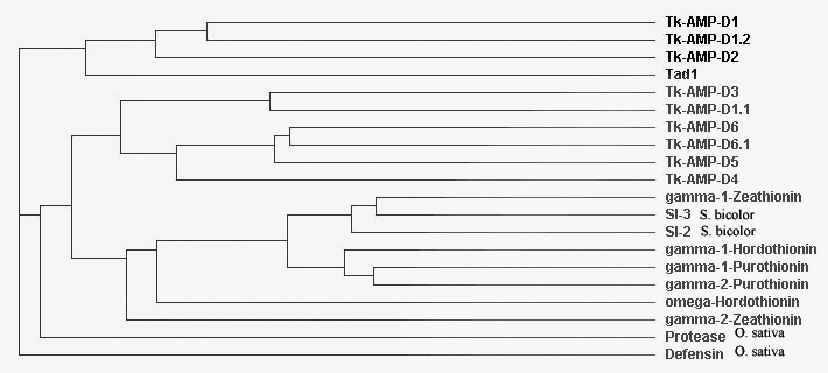
Рис. 18. Филогенетическое древо, отражающее эволюционные связи между представителями семейства злаковых, построенное на основе данных о первичной структуре дефензинов.
6.4. Локализация детерминант антифунгальной активности в молекулах дефензинов
Для локализация детерминант антифунгальной активности в молекулах дефензинов злаков было проведено сравнение аминокислотных последовательностей дефензинов пшеницы Tk-AMP-D1 и Tk-AMP-D6, обладающих низкой антифунгальной активностью к ряду фитопатогенов (A. consortiale, B. cinerea, H. sativum, F. сulmorum, C. graminicola и D. maydis) с дефензином ежовника обыкновенного Echinochloa crusgalli (L.) Beauv., обладающим высокой ингибирующей активностью по отношению к этим патогенам. Дефензины ежовника и пшеницы обладают высокой гомологией первичной структуры, особенно в N-концевой области молекулы (остатки 1-27), которая достигает 85%. Таким образом, можно предположить, что различия в антифунгальной активности между дефензинами Tk-AMP-D1 T. kiharae и Ec-AMP-D1 E. crusgalli связаны с различиями в аминокислотной последовательности в С-концевой области молекул.
Выявленная нами высокая гомология дефензинов в N-концевой области молекул свидетельствует о значительном консерватизме этого участка в эволюции, который сохранился почти неизменным после дивергенции триб Paniceae (E. crusgalli) и Triticeae (T. kiharae). Можно предположить, что консерватизм этой части молекул дефензинов связан с какими-то общими жизненно важными функциями (например, структурной), в то время как С-концевой участок молекул отвечает за функциональную специфичность.
6.5. Дефензины видов Triticum и Aegilops
Помимо дефензинов T. kiharae, были выделены и охарактеризованы также дефензины диплоидных видов родов Triticum и Aegilops, предполагаемых доноров А, В и D геномов. Было установлено, что структура D дефензинов высоко консервативна в эволюции. Кроме того, набор дефензинов у исследованных видов специфичен: каждый из них содержит лишь некоторые дефензины, обнаруженные у гексаплоидной пшеницы (табл. 7). В результате была определена локализация генов, кодирующих дефензины, в геномах полиплоидной пшеницы. Сходство в наборе дефензинов у устойчивых (T. kiharae) и восприимчивых форм пшеницы (T. aestivum сорта Хакасская и Родина) свидетельствует о том, что различие в устойчивости, по всей видимости, связано с уровнем экспрессии генов дефензинов.
Таблица 8.
Состав дефензинов в семенах T. kiharae и родственных видов
| Вид | Геномный состав | D1 | D1.1 | D1.2 | D2 | D3 | D3.1 | D4 | D5 | D6 | D6.1 |
| T. monococcum | AbAb | - | + | + | - | - | - | + | + | - | - |
| Ae. speltoides | BB | - | - | - | + | - | + | - | - | - | + |
| Ae. squarrosa | DD | - | - | - | + | + | - | - | - | + | - |
| T. timopheevii | AbAbGG | + | - | - | + | - | - | + | + | - | + |
| T. militinae | AbAbGG | - | - | + | + | - | - | + | + | - | + |
| T. aestivum сорт Родина | AuAuBBDD | + | + | - | + | + | - | + | + | + | + |
| T. aestivum сорт Хакасская | AuAuBBDD | + | + | - | + | + | - | + | + | + | + |
| T. kiharae | AbAbGGDD | + | + | - | + | + | - | + | + | + | + |
6.6. Исследование нового структурного типа 10-Cys гевеиноподобного пептида WAMP T. kiharae
Из пшеницы были выделены и просеквенированы новые гевеиноподобные пептиды WAMP-1a и WAMP-1b, различающиеся по наличию дополнительного С-концевого остатка аргинина у WAMP-1b (рис. 19).
WAMP-1a и WAMP-1b представляют собой новый структурный тип АМП. Они обладают гомологией с гевеиноподобными АМП, однако в их молекулах содержится 10 остатков цистеина, расположенных уникальным образом, поэтому их можно отнести к новому структурному типу 10-Cys гевеиноподобных пептидов с новым цистеиновым мотивом (рис. 19). Обращает на себя внимание сходство пептидов WAMP с хитин-связывающими доменами хитиназ класса 1 (рис. 19). Другой особенностью новых пептидов является замена консервативного остатка серина на глицин в углеводсвязывающем сайте, хотя способность связываться с хитином сохраняется.
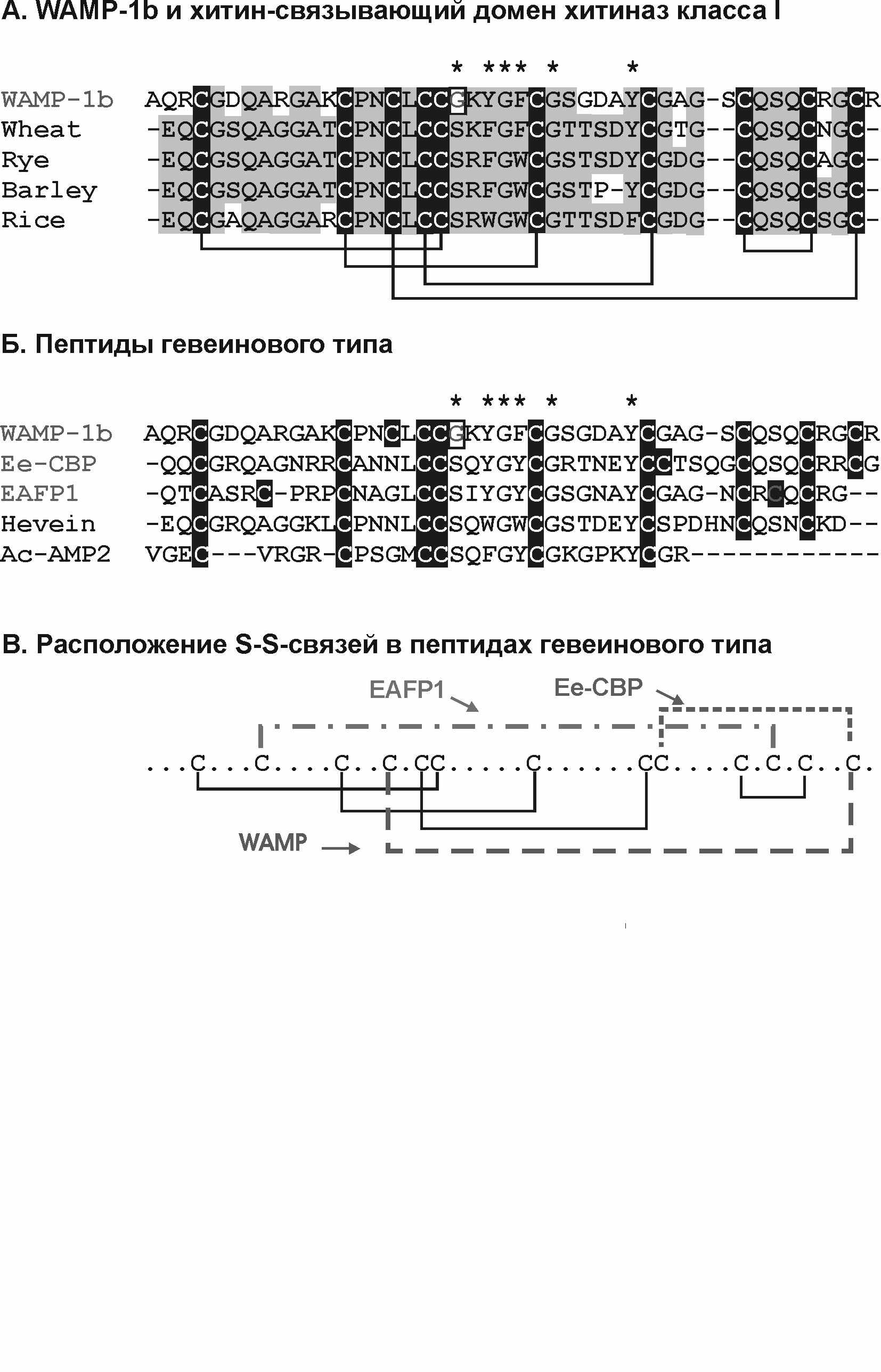
Pис. 19. Сравнение аминокислотных последовательностей пептидов WAMP с некоторыми гевеиноподобными АМП и хитин-связывающими доменами хитиназ класса I. Консервативные остатки цистеина выделены черным, идентичные или сходные аминокислотные остатки выделены серым цветом. Консервативные остатки углевод-связывающего сайта отмечены звездочками, как и остаток глицина, заменяющий консервативный остаток серина в хитин-связывающем сайте.
Исходя из гомологии с гевеином, мы предположили существование следующих дисульфидных связей в пептидах WAMP: Сys4-Сys19, Сys13-Сys25, Сys18-Сys32, Сys37-Сys41. дополнительная пятая дисульфидная связь образована между Сys16 и Сys44. Такое расположение дисульфидных связей подтверждается рентгеноструктурным анализом хитиназы класса 1 риса (Kezuka, Nishizawa, Watanabe, Nonaka, accession number 2DKV_A).
6.7. Получение рекомбинантного пептида WAMP-1a
Для того чтобы получить достаточное количество пептида для функциональных исследований, была проведена гетерологичная экспрессия синтетического гена этого пептида в клетках E. coli. Целевой пептид экспрессировали в составе гибридного белка с тиоредоксином, поскольку было известно, что в этом случае достигается высокий выход целевого пептида в нативной конформации. Используя данные по аминокислотной последовательности пептида WAMP-1a, из синтетических олигонуклеотидов методом ПЦР был сконструирован ген, кодирующий этот пептид. Этот ген был клонирован в экспрессионный вектор pET-32b и образовавшаяся плазмида (pET-32-M-WAMP-1a) была использована для трансформации клеток E. coli BL21(DE3). Образование гибридного белка Trx- WAMP-1a и его очистку контролировали с помощью электрофореза в полиакриламидном геле в присутствии ДДС-Na (pис. 20). Химерный белок обрабатывали бромцианом, и рекомбинантный пептид WAMP-1a очищали путем ОФ-ВЭЖХ (рис. 20). Рекомбинантный пептид обладал тем же временем удерживания на колонке, что и нативный WAMP-1a при ОФ-ВЭЖХ и имел ту же N-концевую аминокислотную последовательность. Его молекулярная масса, измеренная методом MALDI-TOF MS, также совпадала с массой нативного пептида. Выход очищенного рекомбинантного пептида WAMP-1a составил около 8 мг/л культуры.
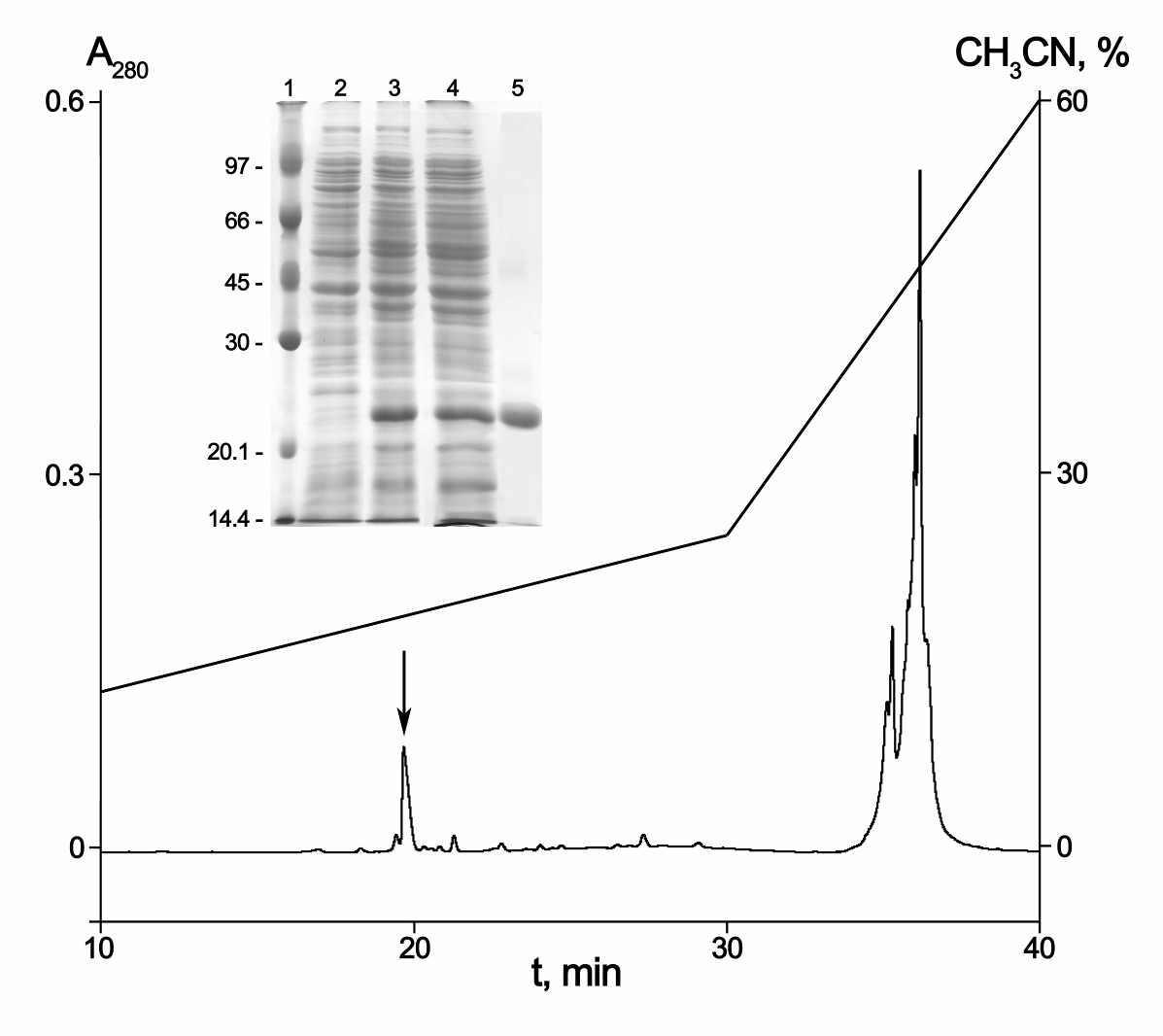
Рис. 20. Экспрессия и очистка гибридного белка Trx- WAMP-1a. ОФ-ВЭЖХ продуктов расщепления гибридного белка Trx-WAMP-1a бромцианом. Фракция, соответствующая WAMP-1a, отмечена стрелкой. На вставке - экспрессия и очистка гибридного белка Trx-WAMP-1a по данным электрофореза в ДДС-ПААГе (10%). (1) маркеры молекулярных масс, (2) лизат клеток E. coli BL21(DE3) с плазмидой pET-32b-M- WAMP-1a, до обработки IPTG, (3) после обработки 0,2 мМ IPTG, (4) растворимая фракция белков, (5) очищенный гибридный белок.
6.8. Антимикробная активность рекомбинантного пептида WAMP-1a
Определение биологической активности пептида в отношении ряда грибов, включая дейтеромицеты и аскомицеты, выявило существенное ингибирование прорастания спор при микромолярных концентрациях пептида, IC50 варьировала от 5 до 30 мкг/мл в зависимости от гриба.
Пептид также обладал способностью ингибировать рост фитопатогенных грамположительных ( C. michiganense) и грамотрицательных (P. syringae и E. carotovora) бактерий. В случае грамположительных бактерий эффект был более выражен. Антифунгальные свойства пептида WAMP-1a, по всей видимости, связаны с его хитинсвязывающей активностью, в то время как активность в отношении оомицета Ph. infestans и бактерий, которые лишены хитина, обусловлена каким-то иным механизмом.
6.9. Установление структуры предшественника пептида WAMP-1a
Была установлена полная нуклеотидная последовательность мРНК, кодирующей пептид WAMP. Методом 3’- и 5’-RACE с использованием вырожденных праймеров, подобранных по аминокислотной последовательности пептида WAMP-1a была определена последовательность его кДНК длиной в 670 нуклеотидов (рис. 21).
ggaggaggcaccaagatcatgaagccacacatgtccgctacggtactgag
agccccgagggtggcggccatcctcctggccgtggtcctggcggcggtgc
tcgccacggccgtgaacggcgcccagaggtgcggcgaccaggcccgcggc
gcaaagtgccccaactgcctctgctgcggtaaatacggcttctgcggcag
tggcgacgcctactgcggcgccggcagctgccagagccagtgccgcggct
gccgggacgacgtcgtggggcaggcgttgccggccgaaccgggttctaca
agagctactgcggcgtcctcggcgtcggccaggggattaaacttgactgc
tacaaccggaggcccttgaacggctgggctcgcgtcgcagtgaagacggc
cggtgactacatagagcgtgagccctggacgaataatctcgacaagtact
agtatgtgaaaaaaataaatggacgaatgatttcccatttgtcttgtctt
atcgtgcatcagcatctttttgtcttcaataagaaatgggacagagaact
tgggtctagccagccggtggtaaagacctcctgaaggggtgtgttaattt
cgtcaatatttagagctctgcaattcagtctttaatatacatttgaaaaa
aaaaaaaaaaaaaaaaaaaa
Рис. 21. Нуклеотидная последовательность кДНК пептида WAMP-1а. Кодоны инициации и терминации трансляции выделены жирным шрифтом.
Оказалось, что структура экспрессируемого предшественника (препропептида) состоит из сигнального пептида и зрелой части, которая включает последовательность зрелого пептида и С-концевой продомен:
MKPHMSATVLRAPRVAAILLAVVLAAVLATAVNGAQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGAGSCQSQCRGCRDDVVGQALPAEPGSTRATAASSASARGLNLTATTGGP
Установлено, что, помимо изучаемого пептида WAMP-1а, в пшенице экспрессируется мРНК двух высокогомологичных ему пептидов сходной структуры, отличающихся от него заменой Аla34 на Lys34 в зрелой части.
6.10. Влияние биотических и абиотических факторов на экспрессию гена WAMP-1а
Была исследована регуляция экспрессии гена пептида WAMP-1а стрессовыми факторами среды. Для оценки уровня экспрессии мРНК в ответ на стрессовые воздействия биотической и абиотической природы использовали метод полуколичественной ОТ-ПЦР. Биотический стресс создавали воздействием грибных патогенов при инфицировании ими прорастающих семян пшеницы. Для изучения воздействия абиотического стресса были выбраны температурный и солевой стресс разной интенсивности. При изучении воздействия температурного стресса достоверных различий обнаружено не было ни в случае гипотермического стресса, ни при воздействии повышенной температуры.
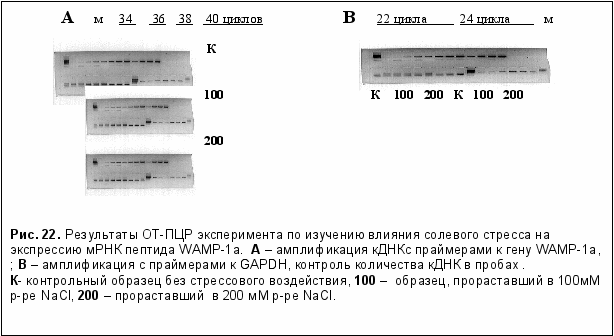
При изучении воздействия солевого стресса было выявлено значительное увеличение экспрессии мРНК пептида WAMP-1a (рис. 22). Под действием соли количество транскриптов изучаемого гена возрастает в 4,3 - 5,7 раза, зависимость от концентрации NaCl не выражена.
При изучении воздействия биотического стресса было установлено, что грибные патогены оказывают различное действие на экспрессию мРНК пептида WAMP-1a. При заражении зерновок непатогенным для пшеницы грибом A. niger не наблюдалось достоверных различий в уровне экспрессии между опытным и контрольным образцом. При заражении патогенными грибами F. oxysporum происходило достоверное увеличение уровня экспрессии мРНК пептида WAMP-1а. Максимальный уровень увеличения экспрессии, наблюдавшийся у зараженных растений по сравнению с контрольной группой, состоял в 5-кратном усилении экспрессии.
При изучении воздействия биотического стресса было установлено, что грибные патогены оказывают дифференцированное действие на экспрессию мРНК пептида WAMP-1a. При заражении зерновок непатогенным для пшеницы грибом A. niger не наблюдалось достоверных различий в уровне экспрессии между опытным и контрольным образцом. При заражении патогенными грибами F. oxysporum происходило достоверное увеличение уровня экспрессии мРНК пептида WAMP-1а. Максимальный уровень увеличения экспрессии, наблюдавшийся у зараженных растений по сравнению с контрольной группой, состоял в 5-кратном усилении экспрессии. Различия между контрольным образцом и образцами, пораженными F. graminearum и H. sativum, не были достоверными.
Полученные нами данные о том, что синтез WAMP-1a при нормальных условиях вегетации осуществляется в 5-дневных проростках, свидетельствует о его роли на ранних стадиях развития растения, а накопление этого пептида при действии патогенов и в условиях солевого стресса дает основание предполагать его участие в устойчивости к биотическому и абиотическому стрессу.
6.11. 4-Cys пептиды
Помимо дефензинов из семян T. kiharae нами было выделено 2 новых АМП, названных Tk-AMP-X1 и Tk-AMP-Х2, содержащих по 4 остатка цистеина в молекуле. С помощью масс-спектрометрии были установлены точные молекулярные массы этих пептидов: 3517 и 3857 Да, соответственно и определены их полные аминокислотные последовательности.
Tk-AMP-X1 1TDDRCERMCQHYHDRREKKQCMKGCRYGESD31
Tk-AMP-X2 1ADDRCERMCQRYHDRREKKQCMKRCRYG28
Было установлено, что остатки цистеина в этих пептидах соединены следующим образом: Сys5-Cys25 и Cys9-Cys21.
Сравнение аминокислотных последовательностей пептидов Tk-AMP-X1 и Tk-AMP-X2 показало, что выделенные пептиды обладают высокой гомологией. Установлено, что они ингибируют прорастание спор некоторых фитопатогенных грибов (F. graminearum, F. verticillioides, C. graminicola и D. maydis) in vitro показало, действуя в микромолярных концентрациях (табл. 9).
Таблица 9.
Антифунгальные свойства пептидов Tk-AMP-X1 и Tk-AMP-Х2*
| Пептид | Tk-AMP-X1 | Tk-AMP-Х2 |
| Fusarium graminearum | 7,5 | 7,5 |
| Fusarium verticillioides | 15 | 10 |
| Diplodia maydis | 30 | 17 |
*Приведены значения IC50, мкМ
Подводя итог исследованию молекулярного полиморфизма антимикробных пептидов пшеницы, следует особо отметить, что в этом виде присутствуют практически все известные у растений семейства АМП.: тионины, дефензины, ноттиноподобные пептиды и липид-переносящие белки. Кроме ранее известных семейств, нами были обнаружены новые семейства, ранее не описанные у растений. Так, было открыто новое семейство глицин-богатых пептидов, семейство 4-Cys пептидов, новое подсемейство D дефензинов и новый структурный тип гевеиноподобных пептидов. Полученные нами результаты значительно расширяют наши представления о чрезвычайном структурном многообразии молекулярных компонентов защитной системы злаков.
Представляло интерес выяснить, является ли высокий молекулярный полиморфизм характерным лишь для злаковых, либо он присущ также другим видам растений. Для решения этого вопроса были исследованы антимикробные полипептиды у представителя двудольных растений семейства Asteraceae – одуванчика лекарственного Taraxacum officinale.
Исследование полипептидов семян Taraxacum officinale, обладающих антифунгальной активностью, показало, что в отличие от T. kiharae, основными детерминантами антифунгальной активности полипептидной природы в семенах T. officinale являются не АМП, а запасные 2S альбумины, представленные несколькими изоформами, каждая из которых состоит из двух субъединиц, соединенных двумя дисульфидными связями. Установлено, что эти белки способны подавлять рост ряда фитопатогенных грибов и оомицета Ph. infestans в микромолярных концентрациях, причем изоформы различаются по активности. Эти результаты свидетельствуют о многообразии и видоспецифичности защитного арсенала растений.
ВЫВОДЫ
1. Разработана методология разделения и очистки запасных и защитных белков растений, позволяющая проводить структурно-функциональный анализ мономерных и полимерных проламинов (сем. Poaceae), 2S альбуминов (сем. Asteraceae), глобулинов (сем. Cucurbitaceae), PR-белков (сем. Solanaceae) и антимикробных пептидов (сем. Poaceae).
2. Впервые у гексаплоидной пшеницы Triticum aestivum проведено определение и сравнение N-концевых аминокислотных последовательностей глиадинов, кодируемых одним кластером генов (1D1 и 1B1), а также кластерами гомеологичных хромосом (D и B). Установлено, что компоненты одного кластера генов, локализованного на коротком плече хромосомы 1D (блок 1D1 у пшеницы Безостая 1) и хромосомы 1В (блок 1В1), существенно различаются по N-концевым аминокислотным последовательностям. У ω-глиадинов выявлено существование трех типов N-концевых аминокислотных последовательностей и показано, что N-концевые аминокислотные последовательности ω-глиадинов, кодируемых геномом D, значительно дивергировали от последовательностей ω-глиадинов генома В. В то же время выявлен значительный консерватизм аминокислотных последовательностей γ-глиадинов, кодируемых хромосомами А, В и D.
3. В ω- и γ-глиадинах обнаружены короткие повторяющиеся олигопептидные мотивы, что дает основание полагать, что гены ω- и γ- глиадинов произошли в результате многократных дупликаций коротких нуклеотидных последовательностей ДНК.
4. Исследование глиадинов двух видов эгилопсов Ae. squarrosa и Ae. longissima – предполагаемых доноров геномов B и D пшеницы, выявило высокую внутривидовую изменчивость этих видов и показало, что у эгилопсов встречаются те же типы N-концевых аминокислотных последовательностей глиадинов, как у гексаплоидных пшениц. Таким образом, хотя гены проламинов и претерпевали изменения в ходе эволюции за счет мутаций и хромосомных перестроек, определенная часть их последовательностей оставалась консервативной в течение длительного эволюционного периода.
5. Дисульфидное картирование двух представителей суперсемейства проламинов - глиадина γ-46 пшеницы и 2S альбумина подсолнечника выявило высокий консерватизм расположения дисульфидных связей как между глиадинами и 2S альбуминами, так и в пределах каждой из этих групп белков. В то же время у других членов суперсемейства проламинов, ингибиторов α-амилаз и трипсина злаков, только две дисульфидные связи с участием двух соседних остатков цистеина сохраняют свое положение. Эти различия, по всей видимости, связаны с необходимостью поддерживать структуру активных центров при взаимодействии с ферментами-мишенями.
6. При исследовании субъединиц глютенина гексаплоидной пшеницы T. aestivum. обнаружена новая D субъединица глютенина и показана ее роль в терминации роста полимерной цепи глютенинов, определяющей качество клейковины пшеницы. Показан консерватизм аминокислотных последовательностей в окружении остатков цистеина у высокомолекулярных субъединиц глютенина HMWx7 и HMWy18, и аллельной пары HMWx7 и HMWy9, обусловливающих хорошие вязко-эластичные свойства клейковины, что свидетельствует о ключевой роли дисульфидных связей в формировании надмолекулярной структуры клейковинного комплекса..
7. При использовании разработанного метода разделения полимеров глютенина в агарозном геле с измерением светорассеяния установлено, что молекулярные массы полимеров глютенина варьирует от 500000 до 5 млн. дальтон. Предложенный метод может быть использован для фракционирования глютенинов сортов пшениц, различающихся по технологическим характеристикам, а также для разделения других высокомолекулярных полимерных белков.
8. Разработаны электрофоретический и хроматографический методы исследования полиморфизма запасных белков тыквенных культур. Показана возможность идентификации генетически отдаленных форм огурца по электрофоретическим спектрам альбуминов, разделенных методом электрофореза в денатурирующих и кислых условиях, а также районированных сортов тыкв по электрофоретическим спектрам кукурбитина и альбуминов, а также по профилю элюции кислоторастворимых альбуминов при ОФ-ВЭЖХ.
9. Впервые изучено наследование субъединиц кукурбитина – основного глобулина семян тыквенных культур. Показано, что: компоненты электрофоретического спектра кукурбитина наследуются кодоминантно при отсутствии эффекта дозы генов. Аллели представлены отдельными полипептидами в электрофоретическом спектре, различающимися по относительной подвижности, а синтез вариабельных компонентов кукурбитина контролируется 4 независимыми локусами.
10. Предложена генетическая классификация кукурбитина и способ записи электрофореграмм этого белка в виде генетических формул, составленных для 38 сортов тыкв отечественной селекции. Показано, что большинство районированных сортов тыкв гетерогенны по кукурбитину. Эффективность предложенного экспресс-метода определения сортовой чистоты семян тыкв по электрофоретическим спектрам кукурбитина превышает традиционно используемый грунтконтроль.
11. Изучены PR-белки, относящиеся к защитной системе растений, у томатов с разными генами устойчивости (Tm-1, Tm-2 и Tm-22) к ВТМ инфекции. Обнаружены специфические белки, характерные для каждого генотипа, которые могут быть использованы в качестве маркеров соответствующих генов в селекционном процессе.
12. Впервые исследованы трансгенные растения табака, экспрессирующие двунитевую РНК, не имеющую гомологии с геномами вируса и табака, и показано повышение устойчивости трансгенных растений к тобамовирусам. В инфицированных трансгенных растениях выявлен индуцируемый инфицированием синтез двух новых полипептидов, относящихся к PR-белкам -семейству β-эндоглюканаз растений. Высказано предположение об участии этих белков в системе антивирусной защиты растений.
13. Среди PR-белков при вирусной инфекции в растениях томатов различного генотипа обнаружены интерфероноподобные белки, что свидетельствует о том, что у растений и животных действует сходный механизм защиты от патогенов, связанный с образованием антивирусных интерфероноподобных веществ.
14. Впервые при систематическом анализе антимикробных пептидов, важнейших компонентов врожденной защитной системы растений, у пшеницы Triticum kiharae Dorof. et Migusch. выделены 24 новых пептида, относящихся к 6 семействам, и установлена первичная структура 12 из них. Обнаружено новое семейство глицин-богатых пептидов растений и новый структурный тип гевеиноподобных пептидов. Выявлена высокая ингибирующая активность этих пептидов в системе in vitro в отношении ряда фитопатогенов.
15. Впервые показано наличие в зерновках пшеницы практически всех известных семейств антимикробных пептидов растений. Высокий молекулярный полиморфизм защитных пептидов у рода Triticum, вероятно, обеспечивает адаптацию к неблагоприятным факторам окружающей среды и является одной их причин, которые обусловливают распространение злаков по всем континентам и климатическим зонам Земного шара.
16. Выявлен молекулярный полиморфизм в пределах семейств антимикробных пептидов Triticum kiharae, который реализуется как на генном уровне (дефензины), так и на посттранскрипционном уровне (4-Cys и 6-Cys пептиды). Показано, что семейство дефензинов Triticum kiharae структурно неоднородно, и состоит, по крайней мере, из трех подсемейств, имеющих, по всей вероятности, различное эволюционное происхождение. Установлено, что детерминанты антифунгальной активности дефензинов семейства Poaceae локализованы в С-концевой области молекулы.
17. Впервые изучены дефензины Triticum monococcum, Triticum urartu, Triticum boeoticum, Aegilops squarrosa и Aegilops speltoides, предполагаемых доноров геномов полиплоидных пшениц. Сравнение дефензинов полиплоидных пшениц с дефензинами диплоидных видов свидетельствует о том, что структура дефензинов высоко консервативна в эволюции. Установлена геномная локализация генов дефензинов. Показано, что дефензины D1, D1.1 и D1.2 кодируются геномом А, D3.1 и D6.1 - геномом В, D3 и D6 - геномом D.
18. Среди антимикробных пептидов Triticum kiharae идентифицирован новый пептид WAMP-1, относящийся к ранее неизвестному структурному типу гевеиноподобных пептидов растений, обладающий высокой антифунгальной и антибактериальной активностями. Определена нуклеотидная последовательность кДНК, кодирующая предшественник пептида WAMP-1а и показано, что в пшенице экспрессируются еще две высокогомологичные мРНК. Установлена структура предшественников пептида WAMP-1а и его гомологов и показано, что они состоят из сигнального пептида, зрелого пептида и С-концевого продомена. Исследование регуляции экспрессии гена пептида WAMP-1 выявило высокий уровень экспрессии гена в проростках, а также повышение экспрессии в ответ на солевой стресс и заражение некоторыми фитопатогенными грибами, что свидетельствует об участии этого пептида в защите растений пшеницы от патогенов и солевого стресса. Разработана система гетерологичной экспрессии в прокариотической системе нового гевеиноподобного пептида Triticum kiharae WAMP-1а. позволившая получить рекомбинантный пептид, идентичный нативному, с высоким выходом. Предложенная бактериальная система экспрессии может быть использована при разработке технологии получения антимикробных пептидов.
19. Сравнение антимикробных полипептидов двух видов растений – однодольных Triticum kiharae и двудольных Taraxacum officinale, свидетельствует о видоспецифичности «защитного арсенала» растений. В отличие от пшеницы, в семенах Taraxacum officinale основными детерминантами антифунгальной активности являются не антимикробные пептиды, а запасные 2S альбумины.
20. Исследования запасных и защитных белков гексаплоидных пшениц и предполагаемых доноров их геномов свидетельствуют о том, что генетический состав ныне существующих диплоидных видов не идентичен составу тех видов, которые участвовали в создании тетра- и гексаплоидных пшениц, а также, что геном В культурных пшениц является сложным геномом, который модифицировался в ходе эволюции за счет интрогрессивной гибридизации с различными диплоидными видами родов Triticum и Aegilops.
СПИСОК ОСНОВНЫХ ПУБЛИКАЦИЙ ПО МАТЕРИАЛАМ ДИССЕРТАЦИИ
Статьи:
- Чугунова Е.Ю., Одинцова Т.И., Егоров Ц.А., Созинов А.А. Электрофоретическое исследование гетерогенности солерастворимой фракции белков семян хлопчатника. Биохимия, 1985, т.50, №6, с. 1030-1038.
- Егоров Ц.А., Одинцова Т.И., Созинов А.А. Селективное выделение омега-глиадинов. Биоорг. химия, 1986, т. 12, №5, с. 599-605.
- Одинцова Т.И., Егоров Ц.А., Созинов А.А. Выделение омега-глиадинов с применением хроматографии на тиопропил-сефарозе. Биохимия, 1986, т. 51, №7, с. 1124-1131.
- Дегтяренко Л.В., Одинцова Т.И., Кононков П.Ф. Исследование гетерогенности белков семян огурца методом электрофореза. Вестник с.-х. науки, 1986, №2, 132-134.
- Батманова Л.С., Одинцова Т.И., Егоров Ц.А., Кононков П.Ф., Созинов А.А. Исследование белков семян капусты методом электрофореза для использования в семенном сортовом контроле, Докл. ВАСХНИЛ, 1987, №7, с. 16-19.
- Одинцова Т.И., Овчинников А.А., Егоров Ц.А., Созинов А.А. N-концевые последовательности глиадинов различных глиадин-кодирующих локусов. Докл. АН СССР, 1987, т. 296, №5, с. 156-164.
- Egorov Ts.A., Odintsova T.I. Microsequence analysis of prolamins with gas-phase protein sequencer, In: Gluten Proteins (Lastity R., Bekes F., eds.), 1987, World Scientific Publishing Co., Singapore, p.434-440.
- Дегтяренко Л.В., Одинцова Т.И., Кононков П.Ф. Электрофорез кукурбитина как метод сортового семенного контроля тыквенных. Доклады ВАСХНИЛ, 1989, №1, с. 17-19.
- Одинцова Т.И., Дегтяренко Л.В., Егоров Ц.А., Кононков П.Ф. Высокоэффективная жидкостная хроматография белков семян тыквенных культур. Доклады ВАСХНИЛ, 1989, №7, с. 26-28.
- Одинцова Т.И., Егоров Ц.А. Выделение и характеристика глиадинов из семян Aegilops squarrosa. Биохимия, 1989, т. 54, №3, с. 396-407.
- Кононков П.Ф., Одинцова Т.И., Дегтяренко Л.В., Батманова Л.С., Крашенинник Н.В., Бунин М.С., Старцев В.И., Осыко Е.В., Егоров Ц.А. Методические указания по анализу белков семян и изоферментов методом электрофореза для контроля сортовой чистоты и селекции капустных и тыквенных культур. ВНИИСиСОК, 1990, Москва, ВАСХНИЛ.
- Одинцова Т.И., Янаудите Р.Л., Егоров Ц.А., Извекова Л.И., Андреева Э.Н., Пухальский В.А. Белки томатов при инфекции ВТМ. Докл. АН СССР, 1990, т. 313, №3, с. 733-736.
- Одинцова Т.И., Егоров Ц.А. N-концевые аминокислотные последовательности омега-глиадинов Aegilops longissima. К вопросу о происхождении геномов полиплоидных пшениц. Биохимия, 1990, т. 55, № 3, с. 509-516.
- Одинцова Т.И., Янаудите Р.Л., Егоров Ц.А., Извекова Л.И., Андреева Э.Н., Пухальский В.А. Обнаружение интерфероноподобных белков в листьях томатов. Докл. АН СССРб 1990, т. 314, № 5, 1250-1252.
- Фетисов А.В., Одинцова Т.И., Извекова Л.И., Пухальский В.А. Белки листьев огурца при вирусной инфекции. Изв. Акад. Наук., сер. биол., 1991, №4, с. 619-624.
- Одинцова Т.И., Турищева М.С., Андреева Э.Н., Пухальский В.А. Структурно-функциональные изменения в хлоропластах томатов с разными генами устойчивости к вирусу табачной мозаики. Генетика, 1996, т. 32, №11, с. 1545-1552.
- Egorov T.A., Odintsova T.I., Musolyamov A.K., Fido R., Tatham A.S., Shewry P.R. Disulphide structure of a sunflower seed albumin: conserved and variant disulphide bonds in the cereal prolamin superfamily. FEBS Letters, 1996, v. 396, p. 285-288.
- Одинцова Т.И., Фетисов А.В., Егоров Ц.А., Извекова Л.И,, Андреева Э.Н., Пухальсий В.А. Структурные исследования белка оболочки вируса зеленой крапчатой мозаики огурца. Докл. РАСН, 1996, №6, с.26-28.
- Egorov T.A., Odintsova T.I., Shewry P.R., Tatham A.S. Characterisation of high Mr wheat glutenin polymers by agarose gel electrophoresis and dynamic light scattering. FEBS Letters, 1998, v. 434, p. 215-217.
- Egorov T.A., Odintsova T.I., Musolyamov A.K. Disulphide mapping of γ-46 gliadin. J. Protein Chem., 1998, v. 17, p. 531-532.
- Егоров Ц.А., Одинцова Т.И., Мусолямов А.Х., Барбашов С.Ф., Пустобаев В.Н., Андерсен Дж., Репсторф П., Попино И. Частичная аминокислотная последовательность глиадина гамма-46. Биохимия, 1998, т. 63, с. 1243-1250.
- Егоров Ц.А., Одинцова Т.И., Мусолямов А.Х. Определение дисульфидных связей в глиадине гамма-46. Биохимия, 1999, т. 64, с. 294-297.
- Oleskina Yu.P., Yurina N.P., Odintsova T.I., Egorov T.A., Otto A., Wittmann-Liebold B., Odintsova M.S. Nucleoid proteins of pea chloroplasts: detection of a protein homologous tо ribosomal protein.. Biochem. Mol. Biol. Int. 1999, v. 47, № 5, p. 757-763.
- Egorov T.A., Odintsova T.I., Musolyamov A., Tatham A, Shewry P., Hojrop P., and Roepstorff P. Biochemical analysis of alcohol soluble polymeric glutenins, D-subunits and omega-gliadins from wheat cv. Chinese Spring. In: Wheat gluten (Shewry P.R., Tatham, A.S., eds.), 2000, The Royal Society of Chemistry, MPG Books Ltd, Botmin, Cornwall, UK, p. 166-170.
- Odintsova T.I., Egorov T.A., Musolyamov A.K., Tatham A.S., Shewry P.R., Hojrup P., Roepstorff P. Isolation and characterization of the high-molecular-weight glutenin subunits 17 and 18 from wheat isogenic line L-88-31. In: Wheat gluten (Shewry, P.R., Tatham, A.S., eds.), The Royal Society of Chemistry, MPG Books Ltd, Botmin, Cornwall, UK, 2000, p. 171-174.
- Одинцова Т.И., Андреева Э.Н., Пухальский В.А., Мусолямов А.Х., Егоров Ц.А. Структурный анализ белка оболочки вируса зеленой крапчатости огурца. Биохимия, 2000, т. 65, №5, с. 672-679.
- Рожнова Н.А., Геращенков Г.А., Одинцова Т.И., Мусин С.М., Пухальский В.А. Синтез новых белков картофеля in vitro при защитном действии арахидоновой кислоты во время вирусной инфекции. Физиол. Раст., 2001, т. 48, №6, с. 897-905.
- Родионова Н.А., Дубовая Н.В., Одинцова Т.И., Грачева Т.И., Безбородов А.М. Выделение эндо-1,4-бета-ксиланазы из Geotrichum candidum 3C с разной сорбционной способностью на нерастворимом субстрате. Прикл. биохим. микробиол. 2002, т. 38, №5, с. 490-494.
- Odintsova T.I., Muller E.-C., Ivanov A.V., Egorov T.A., Bienert R., Vladimirov S.N., Kostka S., Otto A., Wittmann-Liebold B., Karpova G.G. Characterization and analysis of post-translational modifications of the human large cytoplasmic ribosomal subunit proteins by mass spectrometry and Edman sequencing. J. Protein Chem., 2003, v. 22, № 3, р. 249-258.
- Odintsova T.I., Pukhalsky V.A., Egorov T.A., and Musolyamov A.K. Isolation and characterization of antimicrobial peptides from Triticum kiharae. Annual wheat newsletter, 2003, v. 49, p. 124-125.
- Коростылева Т.В., Одинцова Т.И., Козловская Г.В., Пухальский В.А. Исследование белков листьев в трансгенных растениях табака, экспрессирующих двунитевую РНК, при вирусной инфекции. Генетика, 2004, т. 40, №4, с. 531-537.
- Odintsova T.I., Pukhalsky V.A., Musolyamov A.K., Egorov T.A. Analysis of antimicrobial peptides from seeds of Triticum kiharae Dorof. et Migush. Annual wheat newsletter, 2004, v. 50, p. 152-153.
- Egorov T.A., Odintsova T.I., Pukhalsky V.A., Grishin E.V. Diversity of wheat antimicrobial peptides. Peptides, 2005, v. 26, p. 2064-2073.
- Rozhnova N., Odintsova T.I., Musin S., Gerashchenkov G. Protein patterns in potato plants in vitro during systemic antiviral resistance induced by alpha-tocopherol acetatae. Proc. Latv. Acad. Sci., 2005, v. 59, p. 67-72.
- Odintsova T.I., Egorov T.A., Musolyamov A.K., Pukhalsky V.A. Gly-rich antimicrobial peptides from seeds of Triticum kiharae Dorof. et Migusch. Annual wheat newsletter, 2005, v. 51, p. 137-138.
- Odintsova T.I., Pukhalsky V.A., Musolyamov A.K., Egorov T.A. Defensins of Triticum kiharae and other wheat species. Annual wheat newsletter, 2006, v. 52, p. 119-121.
- Odintsova T.I., Egorov T.A., Musolyamov A.K., Odintsova M.S., Pukhalsky V.A., Grishin E.V. Seed defensins from T. kiharae and related species: Genome localization of defensin-encoding genes. Biochimie, 2007, v. 89, p. 605-612.
- Рожнова Н.А., Одинцова Т.И., Геращенков Г.А. Белковый состав листьев табака при индукции антивирусной устойчивости активаторами защитных реакций и ВТМ-инфекцией. Физиол. раст., 2007, т.54, № 6, с. 870-875.
- Odintsova T.I., Egorov T.A., Musolyamov A.K.,Korostyleva T.V. Kozlovskaya G.V. Pukhalsky V.A. Defensins of Triticum kiharae and diploid Triticum and Aegilops species, Annual wheat newsletter, 2007, v. 53, p. 78-79.
- Одинцова Т.И., Егоров Ц.А., Коростылева Т.В., Козловская Г.В., Пухальский В.А. Исследование генома культурных растений и их сородичей: исследование спектров дефензинов у устойчивых и восприимчивых к фитопатогенам видов пшеницы. «Динамика генофондов», ФИАН, 2007, с. 139-141, ISBN978-5-902622-13-0.
- Пухальский В.А., Одинцова Т.И., Извекова Л.И., Андреева Э. Н. Коростылева Т.В., Истомина Е.А., Славохотова А.А., Шиян А.Н.. Козловская Г.В., Оболенкова Л.А., Бадаева Е.Д., Билинская Е.Н. Проблемы естественного и приобретенного иммунитета растений. К развитию идей Н.И. Вавилова, Вестник ВОГиС, 2007, т.11, №3/4, с. 631-650.
- Славохотова А.А., Андреева Э.Н., Шиян А.Н., Одинцова Т.И., Пухальский В.А. Особенности генов белка оболочки российских штаммов вируса зеленой крапчатой мозаики огурца, Генетика, 2007, т.43, № 11, 1535-1540.
- Odintsova T.I., Rogozhin E.A., Baranov Yu, Musolyamov A.Kh., Yalpani N., Egorov Ts.A., Grishin E.V. Seed defensins of barnyard grass Echinochloa crusgalli (L.) Beauv. Biochimie, 2008, v. 90, p. 1667-1673.
- Odintsova T.I., Korostyleva T.V., Odintsova M.S., Pukhalsky A.V., Grishin E.V., Egorov Ts.A. Analysis of Triticum boeoticum and Triticum urartu seed defensins: To the problem of the origin of polyploid wheat genomes. Biochimie, 2008, v. 90, p. 939-946.
- Рогожин Е.А., Одинцова Т.И., Мусолямов А.Х., Смирнов А.Н., Бабаков А.В., Егоров Ц.А., Гришин Е.В. Выделение и характеристика нового липид-переносящего белка из зерновок ежовника обыкновенного (Echinochloa сrusgalli). Прикладная биохимия и микробиология, 2009, т. 45, №4, с. 403-409.
- Ощепкова Ю. И., Вешкурова О. Н., Рогожин Е. А., Мусолямов А. Х., Смирнов А. Н., Одинцова Т.. И., Егоров Ц. А., Гришин Е. В., Салихов Ш. И. Выделение липидпереносящего белка Ns-LTP1 из семян чернушки посевной (Nigella sativa). Биоорг. химия, 2009, т. 35, №3; с. 344-349.
- Odintsova T.I., Vassilevski A.A., Slavokhotova A.A., Musolyamov A.K., Finkina E.I., Khadeeva N.V., Rogozhin E.A., Korostyleva T.V., Pukhalsky V.A., Grishin E.V., Egorov T.A. A novel antifungal hevein-type peptide from Triticum kiharae seeds with a unique 10-cysteine motif. FEBS J. 2009, v. 276, № 15, p. 4266-4275.
- Odintsova T.I., Rogozhin E.A., Sklyar I.V., Musolyamov A.K., Kudryavtsev A.M., Pukhalsky V.A., Smirnov A.N., Grishin E.V., Egorov T.A. Antifungal activity of storage 2S albumins from seeds of the invasive weed dandelion Taraxacum officinale Wigg. Protein Pept Lett. 2009, v. 17.
- Utkina L.L., Slavokhotova A.A., Odintsova T.I., Korostyleva T.V., Pukhalskiy V.A., Musolyamov A.K., Egorov T. A. Novel antimicrobial peptides from seeds of Triticum kiharae and Leymus arenarius. Annual Wheat Newsletter, 2009, v. 55, p. 181-183.
Глава в монографии:
Odintsova T.I. and Egorov Ts.A. Wheat antimicrobial peptides. In: Peptidomics: Methods and Applications (Soloviev M., Andren P., Shaw C., eds.), 2008, p. 99-117. John Wiley & Sons, Hoboken, New Jersey.
Авторские свидетельства:
1. Авторское свидетельство №1498410 на изобретение «Способ определения сортовой чистоты семян тыквенных культур», Одинцова Т.И., Дегтяренко Л.В., Кононков П.Ф., зарегистр. в Гос. реестре изобретений СССР 27.05.1987.
2. Авторское свидетельство № 1630703 на изобретение «Способ контроля сортовой чистоты семян крестоцветных культур», Одинцова Т.И., Батманова Л.С., Глазкова- Степаненко И.С., Кононков П.Ф., зарегистр. в Гос. реестре изобретений СССР 01.11.1990.
Патент:
Патент на изобретение №2380374 (РФ) «Пептид, обладающий антимикробной активностью», Одинцова Т.И., Егоров Ц.А., Гришин Е.В., Василевский А.А., Мусолямов А.Х., Рогожин Е.А., Славохотова А.А., Пухальский В.А., Шиян А.Н., зарегистр. в Гос. реестре изобретений РФ 27.01.2010.
Материалы конференций:
1. Одинцова Т.И., Егоров Ц.А. S-бедные проламины. Выделение и характеристика. Всесоюзн. симп. «Молекулярные механизмы генетических процессов», 1987, Москва,, c.133.
- Kononkov P.F., Degtyarenko L.V., Odintsova T.I. Identification of Cucurbitaceae species and varieties by electrophoresis of cucurbitin. III International symposium ISTA, 1988, Leningrad,
- Egorov T.A., Odintsova T.I., Pukhalsky V.A., Grishin E.V. Wheat peptidomics. 2nd International Conference “Genomics, Proteomics and Bioinformatics for Medicine”, 2004, Moscow.
- Егоров Ц.А., Одинцова Т.И., Бабаков Е.В, Пухальский А.В., Гришин Е.В. Разнообразие антимикробных пептидов растений. III сьезд общества биотехнологов России, 2005, Москва, с.111.
- Одинцова Т.И., Егоров Ц.А., Коростылева Т.В., Козловская Г.В., Пухальский В.А. Исследование дефензинов пшеницы Triticum kiharae Dorof. et Migusch. III международный симпозиум «Проблемы биохимии, радиационной и космической биологии», 2007, Дубна, Россия, с. 178-181.
- Одинцова Т.И., Егоров Ц.А., Коростылева Т.В., Козловская Г.В., Пухальский В.А. Уникальный антимикробный пептид семян пшеницы Triticum kiharae Dorof. et Migusch. Международная конференция «Современная физиология растений: от молекул до экосистем», 2007, Сыктывкар, Россия, с. 301-302.
- Одинцова Т.И., Рогожин Е.А., Жабон Е.О., Пухальский В.А., Егоров В.А. Антимикробные пептиды растений. Международная конференция «Научное наследие Н.И. Вавилова - фундамент развития отечественного и мирового сельского хозяйства», 2007, Москва РГАУ-МСХА им. К.А. Тимирязева, с. 178-179.
- Одинцова Т.И. Дефензины семян пшеницы. III Российский симпозиум «Белки и пептиды», 2007, Пущино-на-Оке, Россия, с.18.
- Уткина Л.Л., Славохотова А.А., Рогожин Е.А., Одинцова Т.И., Пухальский В.А., Кудрявцев А.М. Экспрессия гена нового антимикробного пептида колосняка песчанного Leumus arenarius в прокариотической системе. V сьезд генетиков и селекционеров, 2009, Москва, с.385.
- Odintsova T.I., Shcherbakova L, Flavel D., Egorov T.A., Suprunova T. Identification of a novel protein, a putative elicitor from a biocontrol Fusarium oxysporum, inducing tomato resistance to Fusarium wilt pathogen. XIV International Congress on Molecular Plant-Microbe Interactions, 2009, Quebec City, Canada, p.22.
- Уткина Л.Л., Славохотова А.А., Коростылева Т.В., Андреев Я.А., Василевский А.А., Рогожин Е.А., Одинцова Т.И., Егоров Ц.А., Гришин Е.В. Гетерологическая экспрессия генов, кодирующих новые защитные пептиды растений зерновок пшеницы Кихара (Triticum kiharae) и колосняка песчаного (Lemus arenarius) в клетках Escherichia coli. V Московский международный конгресс «Биотехнология: состояние и перспективы развития», 2009, Москва.
- Коростылева Т.В., Рогожин Е.В., Андреев Я.А., Одинцова Т.И. Изучение влияния биотических и абиотических факторов на экспрессию гена антимикробного пептида WAMP-1а из пшеницы Triticum kiharae Dorof et Migush, Всероссийская научная конференция «Устойчивость организмов к неблагоприятным факторам внешней среды», 2009, Иркутск, Россия, с. 247-250.