
Сущность жизни
| Вид материала | Документы |
- В. П. Крючков Рассказы и пьесы, 957.62kb.
- Тематическое планирование курса «Обществознание», 10-11 классы, 470.24kb.
- Мировоззрение, его сущность, роль в жизни людей, 1231.6kb.
- Экзаменационные вопросы по философии, 16.84kb.
- Г. А. Василевич, доктор юридических наук, профессор сущность конституции, 68.32kb.
- Мы представляем фрагменты книги ш. А. Амонашвили «школа жизни» (М.: Издательский дом, 882.19kb.
- Прогнозирование и планирование: основные понятия, сущность и сфера применения. Изучив, 994.42kb.
- Программа для поступающих на направление подготовки магистратратуры 080200 «менеджмент», 256.74kb.
- Размножение и индивидуальное развитие организмов, 44.8kb.
- Закономерности возникновения жизни в космосе., 3156.97kb.
Современное состояние теори гена ( свойства гена )
- Выступает как кодирующая система
- Обладает способностью к ауторепродукции ( репликации )
- Обладает способностью к мутациям ( элементарная единица мутации гена - мутон )
- Обладает способностью к рекомбинации ( элементарная единица рекомбинации гена - рекон )
- Обладает дискретностью действия
- Существуют структурные , функциональные , регуляторные и модуляторные гены
- Занимает определённый участок хромосомы – локус
Строение гена
- Генетический материал внутри гена сложно организован и имеет линейный порядок
- Ген состоит из многих мутационных мест (сайтов) , разделяемых при рекомбинации
Цистрон - наименьший сегмент ДНК ( 800 -1200 пар оснований ) , мутация которого сопровождается возникновением мутантного фенотипа - элементарная функциональная единица гена ( определяет синтез одного полипептида )
- Ген у эукариот состоит из нескольких обязательных элементов :
- регуляторная зона - регулирует активность гена в той или иной ткани на определённой стадии онтогенеза
- промотор - последовательность ДНК до 80 -100 пар нуклеотидов , ответственная за связывание РНК-полимеразы , осуществляющей транскрипцию данного гена
- структурная зона - часть гена , содержащая информацию о первичной структуре соответствующего белка-фермента ( существенно короче регуляторной зоны , но несколько тысяч пар нуклеотидов )
- терминатор - последовательноть нуклеотидов в конце гена , прекращающая транскрипцию
- структурная часть гена состоит из нуклеотидных последовательностей двух типов :
- Экзоны - участки ДНК , несущие информацию о строении белка ( входят в состав зрелой и-РНК )
- Интроны - участки ДНК не кодирующие структуру белка ( транскрибируются , но в состав зрелой и-РНК не входят , т. к. «вырезаются » в процессе сплайсинга )
Сплайсинг - ферментативный процесс вырезания интронов из молекулы РНК и сращивания экзонов при образовании зрелой и-РНК
Классификация генов
- Структурные гены - гены , кодирующие развитие конкретных признаков ( продуктом первичной активности гена является либо и-РНК и далее полипептид , либо р-РНК и т-РНК )
- Гены - модуляторы - гены , смещающие развитие признака в ту или иную сторону ( например , частоту мутирования структурных генов ) ; могут быть ингибиторами или супрессорами , подавляющими активность или интенсификаторами - повышающими активность генов
- Гены - регуляторы - гены, регулирующие активность структурных генов ( время включения различных локусов в онтогенезе )
Генотип - совокупность всех аллелей (генов) организма, полученных от родителей ( вся совокупность наследственной информации организма ) ; совокупность генов диплоидного набора хромосом клетки
- генотип будучи дискретным ( состоящим из отдельных генов ) функционирует как единое целое
Геном - совокупность генов , содержащихся в гаплоидном наборе хромосом клетки
Фенотип - совокупность всех внутренних и внешних признаков и свойств особи , сформировавшаяся на основе генотипа в процессе её онтогенеза, т. е. реализованная часть генотипа
- развивается при взаимодействии генотипа со средой обитания может относительно сильно варьировать у одной особи ) и
Понятие аллели
- Большинство генов существует в популяции в виде двух или большего числа альтернативных вариантов - аллелей
Аллель - различные формы одного и того же гена( признака ) , расположенные в одинаковых участках (локусах) гомологичных хромосом ( определяют альтернативные варианты развития одного и того же признака
- Все аллели данного признака ( гена ) локализуются в одной и той же хромосоме в определённом её участке - локусе ( в соответствующем локусе хромосомы может находиться лишь один из всех возможных аллелей конкретного гена )
Локус - сегмент ( участок ) хромосомы , в котором локализован ген
- Новые аллели возникают путём мутаций в одном и том же локусе хромосомы ( создаётся т. н. серия множественных аллелей , рассеяных в популяции данного вида - множественный аллелизм
Множественный аллелизм - явление существования в популяции более двух альтернативных аллельных генов , имеющих различное проявление в фенотипе ( например , признак цвета глаз у человека имеет в популяции множество аллельных генов , локализованных в одном локусе определённой хромосомы )
- Аллели отличаются друг от друга содержанием наследственой информации о признаке, развитие которого контролирует ген
- Каждый признак организма представлен в его кариотипе парой аллелей в силу наличия гомологичных хромосом ( одна из них всегда отцовская , другая - материнская ) ; в одной гамете может находиться только один аллель
Гомозигота ( по данному признаку ) - организм , содержащий одинаковые гены данной аллельной пары , образующий один сорт гамет по данному признаку и не расщепляющийся по фенотипу при скрещивании с себе подобными - АА или аа
Гетерозигота ( по данному признаку ) - организм , содержащий разные гены данной аллельной пары , образующий несколько сортов гамет , отличающихся аллелями и расщепляющийся на разные фенотипы при дальнейшем размножении - Аа
Доминантный аллель( ген, признак ) – аллель ( ген, признак ) « сильный», подавляющий, всегда проявляющийся в фенотипе
- Его проявление не зависит от наличия в организме другого аллеля данной серии ( всегда реализуется фенотипически т. к. кодирует более устойчивую форму фермента ) ; обозначается заглавной буквой алфавита - А
Рецессивный аллель( ген, признак ) – аллель ( ген, признак ) « слабый », подавляемый, обеспечивающий развитие признака лишь в отсутствии других аллелей данного гена
- обозначается прописной буквой алфавита - а ( проявляет своё действие только в гомозоготном состоянии - аа и не проявляющийся у гетерозигот - Аа )
Кодоминантные аллели - аллели в одинаковой мере функционально активные в случае их совместного присутствия в генотипе
Взаимодействие аллелей
- Полное доминирование - явление подавления фенотипического проявления признака доминантным аллелем (геном) действия альтернативного аллеля в гетерозиготе
- Неполное доминирование - взаимодействие двух аллелей , дающее в гетерозиготе промежуточный фенотип
- Кодоминантность - независимое друг от друга проявление аллелей в гетерозиготе
Моногибридное скрещивание
Моногибридное скрещивание - скрещивание родителей , отличающихся по проявлению одного признака ( одной альтернативной парой аллелей )
- Мендель проводил опыты с горохом ( очень удачный объект для генетического исследования т. к. горох имеет множество сортов , отличающихся только одним , двумя или несколькими признаками способен к само- и перекрёстному опылению , просто разводятся , имеют короткий период развития )
- Для скрещивания использовались экземпляры , относящиеся к чистым линиям , т. е. растениям , при самоопылении которых в ряду поколений не наблюдалось расщепления по изучаемому признаку
- Для записи проведённого скрещивания и его анализа Мендель ввёл буквенную символику
Буквенная символика по Г. Менделю
Р - перента (лат.) - родительские организмы , взятые для скрещивания
F - филие (лат.) - дети , гибридное потомство
х - скрещивание
А - доминантный признак ; а - рецессивный признак ( для обозначения аллелей других признаков используются любые другие буквы - В и в , С и с и т. д. )
АА - доминантная гомозигота ; аа - рецессивная гомозигота
Аа - гетерозигота при моногибридном скрещивании
АаВв - дигетерозигота при дигибридном скрещивании
- генотипическую формулу материнской особи записывают первой , а отцовскую - второй
- в первой строке записывают генотипические формулы родителей , во второй - типы их гамет (G) , в третьей - генотипы первого поколения и т. д.
- Для скрещивания были взяты два сорта , отличающиеся окраской семян - она м. б. жёлтой и зелёной
А - жёлтая окраска семени ( доминантный признак )
а - зелёная окраска семени ( рецессивный признак )
тогда , Р ♀ АА х ♂ аа
жёлтые зелёные
Гаметы (G) А а
F2 Аа 100 % жёлтые
I закон Менделя ( закон единообразия гибридов первого поколения или закон доминирования) - при скрещивании гомозиготных особей , отличающихся по одной паре альтернативных признаков , всё потомство в первом поколении единообразно как по фенотипу , так и по генотипу
- Мендель ставил опыты таким образом , что в одном эксперименте материнские растения имели жёлтые семена , а отцовские - зелёные , а в другом - наоборот ( такая система из двух скрещиваний носит название реципрокного скрещивания ; при этом одно из скрещиваний (любое) называется прямым , а другое - обратным ) ; в данном случае результаты прямого и обратного скрещивания были одинаковыми
Доминирование - явление преобладания признака одного из родителей у всех гибридных потомков
Доминантный признак - сильный , подавляющий , всегда фенотипически проявляющийся
Рецессивный признак - « слабый » , подавляемый , временно исчезающий , т. е. не проявляющийся у гетерозигот ( фенотипически проявляется только у рецессивных гомозигот - аа )
Наследование некоторых признаков человека
-
Признаки
Типы наследования
доминантный
Рецессивный
Размер глаз Цвет глаз Тип глаз Острота зрения Размер носа Форма носы .. Ширина ноздрей
Ширина ушей Длина ушей Подбородк Строение волос .. Цвет волос Поседение волос Облысение Овал лица Толщина кожи Цвет кожи Наличие веснушек Рост Преобладание руки Тип голоса у мужчин Тип голоса у женщин Щель между резцами Кисть руки Музыкальный слух Характер нижней губы Брови Ямочки на щеках Цвет глаз Зубы при рождении Группа крови Наследственная глухота Мочка уха Резус-фактор Свёртываемость крови Наличие пигмента Переносица Форма черепа Строение ногтей
Большие Карие Монголоидный Близорукость Крупный Острый , узкий , выпуклый с горбинкой , прямой Широкие
.. Широкие Длинные Длинный , прямой , широкий С мелкими завитками , вьющиеся , волнистые ,жёсткие прямые , «ёжик» Рыжие , каштановые До 25 лет У мужчин Круглое Толстая Смуглая Имеются Нормальный Праворукость Бас Сопрано Имеется С 6 , 7 пальцами ( полидактилия ) Имеется Толстая отвисающая , полные губы Мохнатые Имеются Карие Имеются А ( II ) , В ( III ) Отсутствует Свободная Положительный Нормальная Имеется Высокая и узкая Короткий Тонкие плоские
Маленькие Серые , голубые Косой Нормальная Средний или маленький Нормальный , широкий , вогнутый .. Узкие
Узкие Короткие Короткий , узкий , скошенный назад Мягкие прямые Светло-русые После 40 лет У женщин Продолговатое Тонкая Белая Отсутствуют Пропорциональная карликовость Леворукость Тенор Альт Отсутствует С 5 пальцами Отсутствует Нормальная , тонкие губы Нормальные Отсутствуют Голубые Отсутсвуют О ( I ) Имеется Приросшая Отрицательный Гемофилия Альбинизм Широкая и низкая Длинный Нормальные
Расщепление ( сегрегация ) генов ( признаков )
- Продолжая эксперименты , Мендель допустил самоопыление гибридов первого поколения
- Во втором гибридном поколении ( F2 ) появились особи и с доминантным и с рецессивным признаком т. е. произошло расщепление признака
Расщепление - явление распределения доминантных и рецессивных признаков среди потомства в определённом числовом соотношении
- Во всех подобных случаях соотношение числа потомков с доминантным и рецессивным признаком было близким к 3 : 1 ( три части потомков имели жёлтый цвет семян , и одна часть - зелёный )
Р ♀ Аа х ♂ Аа
жёлтые жёлтые
Гаметы (G ) А , а А , а
F2 АА ; Аа ; Аа ; аа
3 жёлтые : 1 зелёный - расщепление по фенотипу
1 АА : 2Аа : 1 аа - расщепление по генотипу
Второй закон Менделя ( закон расщепления ) - при скрещивании двух потомков первого поколения ( двух гетерозиготных особей ) во втором поколении наблюдается расщепление по фенотипу в отношении 3 : 1( 3 части потомков имеют доминантный , а 1 часть - рецессивный фенотип ) и по генотипу 1 : 2 : 1 ( 1 АА : 2Аа : 1аа )
- Данная закономерность носит статистический характер , т. е. справедлива лишь для большого числа исследуемых особей ( закон больших чисел )
- При дальнейших скрещиваниях гибридов F2 ( т. е. их многократном самоопылении ) и получении гибридов F3 , F4 и т. д. оказалось , что особи с генотипом АА и аа не расщеплялись , а особи Аа - расщеплялись в отношении 3 : 1
- Выводы :
- Отдельные признаки организмов при скрещивании не исчезают , а только временно подавляются и сохраняются в потомстве
- Каждая гамета получает лишь один ген из данной аллельной пары , причём количество гамет , несущих разные аллели соответствующего гена , одинаково
- Мужские и женские гаметы , несущие разные аллели одного гена , при оплодотворении комбинируются случайным образом
- Менделевские законы доминирования и расщепления являются универсальными , им подчиняются все живые организмы независимо от сложности их организации
Закон ( правило ) чистоты гамет ( Г. Мендель , 1865 г. )
- Рецессивный ген (а) в гетерозиготе (Аа) не проявляется , но не утрачивается , не изменяется и не смешивается с доминантным геном
- Аллельные гены , находясь в гетерозиготном состоянии , не сливаются , не разбавляются , не изменяют друг друга , поэтому в F2 прявляются в полном объёме , в « чистом » виде
- Гибриды F1 - Аа образуют не гибридные гаметы , а два сорта гамет , каждый из которых несёт либо полный ген А , либо - а , т.е. несут только один полный ген из аллельной пары
Правило чистоты гамет - при образовании половых клеток аллели не смешиваются , в каждую из гамет попадает только один ген из аллельной пары
Следствие - каждая гамета имеет по одному полному аллелю всех признаков организма и не содержит ( «чиста» ) другого аллеля данного гена
Цитологическое обоснование правила чистоты гамет
- В соматических клетках и зиготе находится диплоидный набор хромосом
- В одинаковых локусах гомологичных хромосом находятся аллельные гены ( у гетерозигот в одной гомологичной хромосоме находится доминантный аллель , в другой - рецессивный )
- При гаметогенезе в мейозе гомологичные хромосомы расходятся по разным клеткам (гаметам) , в результате образуются два сорта гамет по данной аллельной паре(одни несут аллель А , другие - а)
- При оплодотворении гаметы , несущие одинаковые или разные аллели , случайно сливаются друг с другом , принося в зиготу по одной хромосоме с одной аллелью (восстановление в зиготе парности гомологичных хромосом ) ; в силу статистической вероятности при достаточно большом числе гамет в потомстве будут 25% - АА , 50% - Аа , 25% - аа , т. е. соотношение 1АА : 2Аа : 1аа
- Для расчёта сочетаний разных типов гамет английский генетик Р. Пеннет предложил испоьзовать решётку , по горизонтали которой записываются отцовские гаметы , а по вертикали - материнские а в образующихся квадратах - сочетание гамет (зиготы)
- Для цитологического обоснования закона расщепления решётка Пеннета будет выглядеть так :
♀ А а

 ♂
♂ А АА Аа
 жёлтый жёлтый
жёлтый жёлтый жёлтый зелёный
а Аа аа
- Расщепление в F2 по генотипу составляет 1АА : 2Аа : 1аа , а по фенотипу - 3 жёлтых : 1 зелёный
Неполное доминирование
- В гетерозоготном состоянии доминантный ген не всегда подавляет проявление рецессивного гена
- В ряде случаев гетерозиготный гибрид F1 - Аа не воспризводит полностью ни одного из родительских признаков и его фенотип носит промежуточный характер ( причины пока не ясны )
Неполное доминирование ( промежуточный характер наследования ) - явление проявления у гетерозигот промежуточного фенотипа
- При срещивании ночной красавицы с красной окраской цветкоа (АА) с растением , имеющим белые цветки (аа) , в F1 у гибридов Аа образуется промежуточная розовая окраска цветка
- При скрещивани растений с розовыми цветками между собой в F2 происходит расщепление по фенотипу 1 красный : 2 розовых : 1 белый и по генотипу 1АА : 2Аа : 1аа , т. е. расщепление по фенотипу и генотипу одинаковое
 P ♀ АА х ♂ аа Р ♀ Аа х ♂ Аа
P ♀ АА х ♂ аа Р ♀ Аа х ♂ Аакрасный белый розовый розовый
G А а G А , а А , а
F1 Àà - 100 % F1 1 АА : 2Аа : 1аа - расщепление по генотипу
розовый 1 красн. : 2 розов. : 1 бел. - расщепление по фенотипу
- Неполное доминирование - широко распространённое явление ( обнаружено при наследовании окраски цветка у львиного зева , строения перьев у птиц , окраски шерсти у крупного рогатого скота , наследственных аномалий человека , например , серповидноклеточной анемии
Анализирующее скрещивание
- Организм , имеющий рецессивный фенотип , обязательно должен быть рецессивной гомозиготой (аа) т.е . генотип организма , имеющего рецессивный признак , определяется по его фенотипу
- Организмы , имеющие доминантный фенотип , могут обладать доминантным гомозиготным ( АА) или гетерозиготным ( Аа) генотипом , т. е. имеют неотличимый фенотип
- Для установления генотипа особи , обладающей доминантным фенотипом ( т. е. её гомо- или гетерозиготности ) , её скрещивают с особью , гомозиготной по рецессивным аллелям - (аа) и устанавливают неизвестный генотип по потомству
- Если от такого скрещивания всё потомство окажется однородным , т. е. расщепления не произойдёт , значит анализируемая особь гомозиготна - АА , если же произойдёт расщепление , то она гетерозиготна ( см. схему 1. и 2. )
1. Р ♀ АА х ♂ аа 2. Р ♀ Аа х ♂ аа
G А а G А , а а
F1 Аа F1 Аа ; аа
- Для гетерозиготной особи характерно расщепление в соотношении 1 : 1
- Определение генотипов имеет большое значение в селекционной работе , медицинской генетике ( например : у обоих родителей полидактилия ( многопалость ) , а ребёнок имеет нормальное строение кисти , следовательно родители гетерозиготны по этому признаку )
Анализирующее скрещивание - скрещивание особи с неизвестным генотипом и доминантным фентипом с особью рецессивной гомозиготой по интересующим признакам с целью установления генотипа особи по гибридному потомству
Дигибридное и полигибридное скрещиние . Третий закон Менделя
Полигибридное скрещивание - скрещивание родительских организмов , отличающихся и анализируемых по нескольким признакам ( по двум признакам или двум парам аллелей при дигибридном скрещивании , трём - тригибридном и т. д. )
- Законы доминирования и расщепления Менделя, наблюдавшиеся при моногибридном скрещивании сохраняются
- Для дигибридного скрещивания Мендель взял гомозоготные растения гороха , отличающиеся по двум генам ( двум парам аллелей ) - окраски семян ( жёлтая и зелёная ) и формы семян ( гладкая и морщинистая )
- Доминантные признаки - жёлтая окраска (А) и гладкая форма (В) семян , соответственно , зелёная окраска (а) и морщинистая форма (b) семени - рецессивны
- Первое поколение гибридов в этом случае будет единообразным по генотипу и фенотипу , т. к. каждое растение образует только один сорт гамет по изучаемым аллелям ( проявляются только доминантные признаки родителей )
Схема записи дигибридного скрещивания
1 . Р ♀ ААВВ х ♂ ааbb или 2 . Р ♀ ааВВ х ♂ ААbb
G АВ аb G аВ Аb
F1 АаВb - жёлт. , морщ. - 100 % F1 АаВb - жёлт. , морщ. - 100 %
- П
 ри скрещивании гибридов первого поколения (F1) во втором поколении (F2) происходит расщепление в строгом соответствии с законом Менделя и образуется четыре фенотипа - жёлтые гладкие , жёлтые морщинистые , зелёные гладкие и зелёные морщинистые
ри скрещивании гибридов первого поколения (F1) во втором поколении (F2) происходит расщепление в строгом соответствии с законом Менделя и образуется четыре фенотипа - жёлтые гладкие , жёлтые морщинистые , зелёные гладкие и зелёные морщинистые
- При образовании гамет у гибридов F1 из каждой аллельной пары генов в гамету попадает только один ( в силу статистической закономерности у дигетерозиготного гибрида (АаВb) возможно образование четырёх сортов гамет в одинаковом количестве - по 25% : АВ , Аb, аВ , аb )
- Число разных типов гамет у гетерозигот при полигибридном скрещивании определяется формулой 2 n , где n - количество пар альтернативных аллелей у родителей ( при дигибридном скрещивании - 22 - четыре сорта гамет , при тригибридном 23 - восемь сортов и т. д.)
- Во время оплодотворения каждая из четырёх типов гамет случайно встречается с любой из гамет другого организма , т. е. возможны 16 комбинаций ( все возможные сочетания мужских и женских гамет можно легко установить с помощью решётки Пеннета
 F1 (Р) ♀ АаВb х ♂ АаВb
F1 (Р) ♀ АаВb х ♂ АаВb


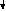


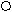

 Образуется 4 фенотипических класса :
Образуется 4 фенотипических класса : 


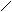
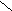 G АВ Аb aB ab 9 жёлтых гладких
G АВ Аb aB ab 9 жёлтых гладких АВ ААВВ ААВb АаВВ АаВb 3 жёлтых морщинистых
ЖГ ЖГ ЖГ ЖГ 3 зелёных гладких Аb ААВb Ааbb АаBb Aabb 1 зелёный морщинистый
F2 ЖГ ЖМ ЖГ ЖМ aB AaBB AaBb aaBB aaBb
ЖГ ЖГ ЗГ ЗГab АаВb Aabb aaBb aabb
 ЖГ ЖМ ЗГ ЗМ
ЖГ ЖМ ЗГ ЗМ- По фенотипу потомство (F2) расщепится на четыре группы в следующем соотношении : 9 : 3 : 3 : 1 ( при неполном доминировании число фенотипических классов будет возрастать )
- Если учитывать результаты расщепления по каждой паре аллелей в отдельности , то получится , что отношение числа жёлтых семян к числу зелёных и отношение числа гладких к морщинистым для каждой пары равно 3 : 1 ( каждая пара признаков при расщеплении в потомстве ведёт себя так же , как в моногибридном скрещивании , т. е. независимо от другой пары признаков ) дигибридное скрещивание есть два независимо идущих моногибридных скрещивания , результаты которых как бы накладываются друг на друга
- Математическим выражением расщепления по фенотипу при дигибридном скрещивании служит формула ( 3 + 1 )2 = 9 + 3 + 9 + 1 , что соответствует числу и относительной частоте каждого фенотипического класса
- В результате случайного сочетания 4 типов гамет при оплодотворении образуется 9 разных генотипов ( генотипических классов ) , проявляющиеся в виде 4 фенотипических классов :
ж
 ёлтые гладкие : ААВВ , ААВв , АаВВ , АаВв - 4 жёлтые морщинистые : ААвв , Аавв - 2 зелёные гладкие : ааВВ , ааВв - 2 т. е. расщепление по генотипу 4 : 2 : 2 :1 зелёные морщинистые : аавв - 2
ёлтые гладкие : ААВВ , ААВв , АаВВ , АаВв - 4 жёлтые морщинистые : ААвв , Аавв - 2 зелёные гладкие : ааВВ , ааВв - 2 т. е. расщепление по генотипу 4 : 2 : 2 :1 зелёные морщинистые : аавв - 2- Число генотипических классов в F2 определяется формулой - 3n Число фенотипических классов в F2 определяется формулой - 2n ( при полном доминировании )
- При тригибридном скрещивании тригетерозогиты (АаВbCc) образуют 8 типов гамет , дающих 64 сочетания ( число фенотипических классов и их численные соотношения устанавливаются по формуле ( 3 + 1 )3 = 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1 ( число фенотипических классов определяется количеством членов после раскрытия скобок - 8 )
- В общей форме , при любых скрещиваниях расщепление по фенотипу происходит по формуле ( 3 + 1 )n , где n - число анализируемых признаков у родителей
III закон Менделя ( закон независимого комбинирования признаков ) - при полигибридном скрещивании гомозиготных особей гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются со всех возможных сочетаниях
- Следствия - в F2 появляются гибридные формы , не свойственные родительским (рекомбинаниные)
- III закон Менделя выполним только при условии , что гены изучаеиых признаков расположены в разных парах гомологичных хромосом (в отличие от I и II законов , которые справедливы всегда)
Генетика пола
Пол - это совокупность признаков и свойств организма , определяющих его участие в размножении
Первичные половые признаки - морфофизиологические особенности организма , обеспечивающие образование гамет , их сближение и соединение при оплодотворении - наружные и внутренние органы размножения ( половые железы и выводящие протоки , добавочные железы , органы внутриутробного развития , наружные половые органы и т. д.)
Вторичные половые признаки - совокупность внешних признаков и особенностей , обеспечивающих обнаружение и привлечение партнёра ( их развитие контролируется гормрнами , синтезируемыми первичными половыми органами - половыми железами )
- Подавляющее большинство животных предствлено особями двух полов - мужского и женского
- Соотношение полов в популяциях раздельнополых организмов в среднем 1 : 1 ( у людей в среднем на каждые 100 девочек рождается 106 мальчиков ) ; такое соотношение полов обеспечивает максимальную вероятность встречи самцов и самок и поддержание оптимальной численности популяций ; в дальнейшем эти соотношения могут сильно изменяться в силу неодинаковой выживаемости особей разного пола ( у человека к 50 годам соотношение мужчин и женщин составляет 85 : 100 , а к 85 годам - 50 : 100 )
- Развитие признаков пола генетически контролируется , т. к. закономерно воспроизводтся в ряду поколений и наследуется как менделирующий признак
- Самцы и самки различаются по набору хромосом
Аутосомы - хромосомы одинаковые в клетках мужских и женских особей ( образуют гомологичные пары )
Половые хромосомы (гетеросомы) - пара хромосом , отличающиеся у разных полов по морфологии и заключённой в них генетической информации
- Большую из половых хромосом принято называть X (икс) - хромосомой , меньшую Y (игрек) -хромосомой ( у некоторых животных Y- хромосома может отсутствовать )
- Зигота человека и других организмов потенциально бисексуальна . Главным фактором , сдвигающим фенотип в мужскую сторону , является Y-хромосома . Выбор направления происходит на 6 -10 неделе эмбриогенеза
- В Y- хромосоме человека находится ген дифференцировки семенников , которые вырабатывают гормоны , обеспечивающие развитие мужских вторичных половых признаков( при отсутсвии Y-хромосомы зачаточные репродуктивные органы дифференцируются в яичники и у зародыша развиваются женские половые признаки )
- Пол будущего организма определяется сочетанием половых хромосом в зиготе в момент оплодотворения
- В зависимости от сочетания половых хромосом в зиготе различают 5 типов определения пола :
- XX , XY- у всех млекопитающих ( в том числе у человека ) , дрозофилы
- XY , XX - у части насекомых ( бабочек , ручейников ) , птиц , рептилий , некоторых амфибий и рыб
- XX , X0 ( 0 обозначает отсутствие Y- хромосомы ) - некоторые насекомые : клопы рода Protenor прямокрылые ( кузнечики)
- X0 , XX - у тли
- гаплоидно - диплоидный ( 2n , n ) встречается , например , у пчёл : самцы развиваются из неоплодотворённых гаплоидных яиц , самки - из оплодотворённых диплоидных ( эти организмы не имеют половых хромосом )
Гомогаметный пол - пол с одинаковыми половыми хромосомами , образующий один вид гамет по половым хромосомам ( XX ) ; все гаметы этого пола несут гаплоидный набор аутосом и одну X - хромосому ( А + X ) , где А - гаплоидный набор аутосом
Гетерогаметный пол - пол с различными половыми хромосомами , образующий два вида гамет , отличающиеся гетеросомами ( X и Y, X и 0) ; количество гамет , содержащих X- или Y- гетеросому приблизительно одинаково и одинаково способны к оплодотворению , что обеспечивает равное количество полов в популяциях большинства организмов
- У двудомных растений , имеющих мужские и женские растения , также изучены половые хромосомы ; гетерогаметными у большинства таких растений является мужской пол
- Пол особи может определяться :
- до оплодотворения яйцеклетки сперматозоидом ( прогамное определение пола )
- в момент оплодотворения ( сингамное определение пола ) ; наиболее часто встречающийся тип
родители 2А + XX х 2А + XY
гаметы А + X А + X , А +Y ( соотношение гамет с X- и Y-хромосомой - 1 : 1 )
зиготы 2А + XX , 2А + XY ( соотношение и зигот - 1 : 1 )
- после оплодотворения ( эпигамное определение пола ) - у морского кольчатого червя бонеллия , если личинка садится на дно , из неё развивается самка , а если прикрепляется к хоботку взрослой самки , то самец
- У дрозофилы Y - хромосома по размеру близка к к X- хромосоме , однако она генетически инертна , т. к. состоит в основном из гетерохроматина и играет незначительную роль в определении пола (особи с кариотипом X0 внешне типичные самцы , но стерильные , а особи с кариотипом XXY - плодовитые самки )
- У многих организмов пол определяется не столько сочетанием в зиготе X- иY-хромосом , сколько соотношением числа X-хромосом и наборов аутосом - половой индекс ( у нормальных самок половой индекс равен 1 ( 2X : 2 А ) , у нормальных самцов - 0,5 ( XY: 2А ) ; при половом индексе более 1 ( 3X : 2А развиваются сверхсамки , при величине ниже 0,5 - самцы , при значении более 0,5 , но менее 1 ( 2X : 3А развиваются интерсексы
Интерсексы - особи , занимающие по половым признакам промежуточное положение между самцами и самками ( не путать с гермафродитами )
- При утрате X- хромосомы одной из клеток на стадии первого деления зиготы развивается организм , половина клеток которого имеет нормальный кариотип (2АXX ) , несёт признаки самки, а другая половина, клетки которой лишены одной X- хромосомы ( 2АXО ) , имеет признаки самца - явление гинандроморфизма
Гинандроморфы - организм , одна часть которы , включая половые желез , женског, а другая - мужского типа
- У человека присутствие Y-хромосомы в кариотипе независимо от количества X -хромосом (2АXXY , 2АXXXY ) обеспечивает развитие мужского пола ; особи с катиотипом 2АXО являются женщинами
- У человека развитие организма по мужскому типу обеспечиваеися не только геном , расположенным в Y - хромосоме и определяющим синтез мужского полового гормона - тестостерона , но и геном , сцеплённым с X - хромосомой , контолирующим синтез белка - рецептора этого гормона
- В развитии признаков пола принимают участие также гены , локализованые в аутосомах
- Пол организмов развивается на основе полученной от родителей наследственной информации и контролируется взаимодействующими генами в половых хромосомах и аутосомах
Сцеплённое с полом наследование
- В половых хромосомах , кроме генов , определяющих развитие пола , локализуются « обычные » фенотипические гены , образующие группы сцепления гетеросом
Аутосомное наследование - это наследование признаков , гены которых локализованы в аутосомах
Сцеплённое с полом наследование - это наследование признаков , гены которых локализованы в половых хромосомах ( открыто Т. Х.Морганом )
- признаки , сцеплённые с полом , наследуются не в соответствии с законами Менделя
- у человека признаки , наследуемые через Y- хромосому , могутбыть только у лиц мужского пола , а наследуемые через X- хромосому - у лиц обоих полов
- признаки , наследуемые через Y-хромосому , называются голандрические ( голандрическое наследование )
Y- сцеплённое ( голандрическое наследование ) - наследование признаков , гены которых локализованы только в Y- хромосоме и передающихся от отца ко всем его сыновьям ( фенотипически проявляются в каждом поколении )
- у человека таких генов совсем немного : гипертрихоз ( развитие волос по краю ушной раковины , перепонки между пальцами , ген дифференцировки семенников
- по генам , локализованным в X- хромосоме женщины могут быть как гомо-,так и гетерозиготными а рецессивные аллели генов проявляются у них только в гомозиготном сотоянии -XаXа ; у мужчин все гены X-хромосомы , даже рецессивные , сразу же проявляются в фенотипе ( такой организм называют гемизиготным
Гемизиготные признаки - признаки , гены которых локализованы только в одной ( X или Y ) половой хромосоме и не имеющие аллельных генов в другой половой хромосоме
- по большинству генов X-хромосомы мужской организм гемизиготен
- Гены , локализованные в X- и Y- хромосомах можно разделить на две группы :
- гены негомологичных участков ( локусов ) X- и Y- хромосом ( гемизиготные , имеющиеся только в X- или Y- хромосоме ) ; наследование в этом случае называется сцеплённым с полом ; различают X-сцеплённое и Y-сцеплённое наследование
X - сцеплённое наследование -
- У человека выявлена локализация в X-хромосоме 95 признаков ; подавляющее их число гемизиготны ( т. е. не имеют гомологичных аллелей в Y-хромосоме ) - это дальтонизм , гемофилия , атрофия зрительного нерва , несахарный диабет , витаминоустойчивый рахит , тёмная эмаль зубов
- рецессивные для женщин гены этих локусов являются доминантными для мужчин ( в силу их гемизиготности ) ; возможен промежуточный характер проявления признака у гетерозигот , например , окраска шерсти у кошек
- гены общих гомологичных участков ( локусов ) , имеющиеся и в X- и в Y-хромосоме ( ген злокачественного новообразования - пигментной ксеродермы , диатеза , судорожных растройств , полной цветовой слепоты ( ахроматии ) и ряда других генетических патологий
- образуют синапсис при коньюгации , возможен кроссинговер
- гены гомологичных участков наследуются одинаково у мужчин и женщи
- признаки гомологичных участков называются неполно ( частично ) сцеплённымис полом
Признаки , ограниченные полом - признаки , гены которых локализованы в аутосомах , но проявляющиеся в зависимости от пола ( у одного пола признак проявится , у другого - нет )
- Проявление этих признаков зависит от соотношения половых гормонов
- Примерами таких признаков является наличие рогов у оленей ( самцы рогаты , а самки безроги ) или яйценоскость птиц , облысение у человека
Наследование

 Моногенное Полигенное
Моногенное Полигенное Аутосомное Сцепленное с половыми хромосомами
Аутосомное Сцепленное с половыми хромосомами
 Рецессивное Доминантное X-сцепленное Y-сцепленное
Рецессивное Доминантное X-сцепленное Y-сцепленноеРецессивное Доминантное
Н

 аследование
аследованиеНезависимое Частично сцепленное Полностью сцепленное
Наследственность - совокупность генетических механизмовь , обеспечивающих структурно - функциональную преемственность в ряду поколений ( обуславливают закономерности наследования )
Наследование - процесс воспроизведения в ряду поколений структурно - функциональной организации конкретного биологического вида
С
 цепленное наследование
цепленное наследование- Известно много случаев отклонения от III закона Менделя , т. е. принцип независимого комбинирования признаков в потомстве распространяется не на все гены .
Менделирующие признаки - признаки , гены которых локализованы в разных парах гомологичных хромосом и поэтому наследующиеся в соответствии с законом независимого распределения и комбинирования признаков в потомстве ( III законом Менделя )
- Количество признаков и генов , обуславливающих их развитие , у любого организма очень велико ( у человека , например , от 100 000 до 1000 000 структурных генов ) , а число хромосом невелико
- В каждой хромосоме оказывается сотни и тысячи неаллельных генов
- Закономерности наследования генов , локализованных в одной хромосоме , установлены выдающимся американским генетиком Т. Х. Морганом на классическом объекте генетических исследований - плодовой мушке дрозофиле ( кариотип дрозофилы составляет 4 пары хромосом крупного размера , она легка в содержении , быстро размножается , имеет множество ярких мутаций )
- При скрещивании гомозиготных линий мух с чёрным цветом тела и укороченными крыльями (аавв) с мухами , имеющими серый цвет тела и длинные крылья (ААВВ) , все гибриды F1 имеют серое тело и длинные крылья ( следовательно , признаки - серое тело и длинные крылья - доминируют ) соблюдается закон единообразия гибридов первого поколения
Р ♀ аавв х ♂ ААВВ . чёрное тело серое тело . укороченные крылья длинные крылья . G ав АВ . F1 АаВв
серое тело, длинные крылья .
- Далее сцепление изучали посредством анализирующего скрещивания , т. е. скрещивания полученных гибридов F1 с линией мух , гомозиготных по рецессивным генам ( при анализирующем скрещивании фенотип потомства прямо отражает типы гамет гетерозиготных родителей ; если гены не сцеплены , то у гетерозиготного организма образуется четыре типа гамет по 25% каждого сорта и , следовательно , четыре вида потомков в равном количестве , как это показано при независимом комбинировании признаков )
- В опытах Моргана при проведении анализирующего скрещивании гибридных самцов из F1 с самками , обладающими чёрным телом и укороченными крыльями , в F2 образуется всего два фенотипических класса , т. е. расщепление по фенотипу 1 : 1 ( 50% мух с серым телом и длинными крыльями - АаВв и 50% - с чёрным телом и укороченными крыльями – аавв) вместо ожидаемого 1 : 1 : 1 : 1 по Менделю :
Р ♀ аавв х ♂ АаВв
G ав АВ , ав
F2 АаВв аавв . . 50% 50%
- Такое отклонение от независимого распределения означает , что изучаемые гены локализованы в одной хромосоме , сцепляются и наследуются совместно ( сцеплённое наследование )
Сцеплённое наследование – явление совместного наследования генв , локализованных в одной хромосоме
Сцепление генов - локализация генов в одной хромосоме
Закон сцеплённого наследования Т. Моргана - гены , локализованные в одной хромосоме , сцепляются и передаются по наследству преимущественно совместно , составляя группу сцепления
Группа сцепления – гены , локализованные в одной хромосоме и наследующиеся преимущественно совместно
- Число групп сцепления равно числу пар хромосом , т. е. гаплоидному числу хромосом ( т. к. в гомологичных хромосомах находятся одинаковые гены ) - у гетерогаметного пола (XY) число групп сцепления на одну больше за счёт Y-хромосомы ; у человека у женщин 23 группы сцепления , у мужчин – 24 ; у вирусов , бактерий и сине-зелёных водорослей , имеющих одну хромосому , все гены принадлежат к одной группе сцепления
- Причиной образования двух фенотипических классов , вместо ожидаемых четырёх , является образование у дигетерозиготы ( АаВв ) только двух типов гамет ( АВ и ав ) в равном количестве , вследствие сцепления генов локализованных в одной хромосоме ( при независимом комбинировании дигетерозигота (АаВв) образует четыре сорта гамет в равном количестве – АВ , Ав , аВ и ав )
- Если в анализирующем скрещивании поменять местами родительские формы , т.е. скрестить дигетерозиготную самку с дигомозиготным рецессивным самцом , то в F2 образуется четыре фенотипических класса в неравном количестве : 41,5% серых длиннокрылых , 41,5% чёрных короткокрылых , 8,5% серых короткокрылых и 8,5% чёрных длиннокрылых :
Р ♀ АаВв х ♂ аавв
G АВ , Ав , аВ , ав ав
F2 АаВв ; Аавв ; ааВв ; аавв
41,5% 8,5% 8,5% 41,5%
- В F2 образуется 17% особей с перекомбинированными родительскими признаками - рекомбинантные или кроссоверные особи
- Такое отклонение от ожидаемого при независимом расщеплении свидетельствует о наличии сцепления
- Сцепление группы генов , локализованных в одной хромосоме , может быть полным и неполным
- Причиной нарушения сцепления служит кроссинговер – перекрёст гомологичных хромосом при их коньюгации в профазе I мейотического деления и возможный обмен гомологичными локусами , вследствие разрыва хромосом в точках контакта . В результате возникают качественно новые хромосомы ( кроссоверные ) , содержащие участки ( гены ) как материнских , так и отцовских хромосом
Кроссинговер – обмен сегментами между гомологичными хромосомами , сопровождающийся рекомбинацией сцеплённых генов ( Т. Морган )
Генетическая рекомбинация – процесс обмена участками между гомологичными хромосомами в результате кроссинговера , приводящий к образованию гамет с новым сочетанием аллелей
- биологическое значение кроссинговера заключается в создании новых , ранее не существовавших комбинаций генов ( комбинативная изменчивость ) и обеспечение повышения выживаемости организмов в процессе эволюции
В результате кроссинговера дигетерозиготная особь со сцеплённой парой признаков образует не два а четыре сорта гамет , причём в неравном количестве ( АВ , ав – родительские гаметы ; в сумме их всегда более 50% и аВ , Ав – кроссоверных или рекомбинантных гамет , в сумме всегда менее 50 %)
- появление рекомбинантных особей ( по 8,5% каждого типа ) обусловленно нарушением сцепления генов при кроссинговере – неполное сцепление
- Чем дальше друг от друга расположены гены в хромосоме , тем выше вероятность перекрёста и кроссинговера и тем больше процент кроссоверных гамет и больший процент рекомбинантных особей , отличных от родителейЧастота рекомбинаций ( кроссинговера ) вычисляется по формуле
Где x – частота рекомбинаций ( расстояние между генами в морганидах ) ; а – количество особей в первой кроссоверной группе ; с – количество особей во второй кроссоверной группе ; n – общее число потомков в данном опыте
- Анализ частоты рекомбинаций при неполном сцеплении позволяет установить :
- Относительное расстояние между генами , расположенными в одной хромосоме ; оно определяется по проценту кроссинговера
- процент кроссинговера между двумя генами равен сумме процентов рекомбинантных особей , отличных от родителей ; в нашем примере было 17% потомков с новыми комбинациями признаков
- за единицу расстояния между генами , находящимися в одной хромосоме принята морганида ( 1 морганида равняется 1% кроссинговера , т. е. эквивалентна 1% кроссоверных гамет ) ; в нашем примере расстояние между генами , определяющими окраску тела и развитие крыльев , равно 17% кроссинговера , или 17 морганидам ( другие сочетания генов дрозофилы отличаются иной частотой кроссинговера )
- процент кроссинговера для разных пар генов колеблется от долей единицы до пятидесяти , не превышая , однако , последнюю цифру ( практически всегда меньше 50) ; при постоянных условиях для каждой пары генов одной хромосомы это значение постоянно
- при расстояниив 50 морганид и более признаки наследуются независимо , несмотря на то , что гены локализуются в одной хромосоме
Правило Моргана – частота ( процент ) кроссинговера между двумя неаллельными генами , расположенными в одной хромосоме , прапорциональна расстоянию между ними т. е. сила сцепления генов обратно прапорциональна расстоянию между ними в хромосоме (чем ближе друг к другу расположены гены , тем реже между ними происходит кроссинговер и наоборот )
- Локализацию генов в одной или разных парах хромосом
- частота рекомбинаций при анализирующем скрещивании менее 50% свидетельствует о сцеплённом наследовании и нахождении пары генов в одной хромосоме ( при значительном расстоянии между генами частота рекомбинаций приблизится к 50% , т.е. значению, соответствующему независимому расщеплению )
- Линейный порядок и взаиморасположение генов в хромосоме ; составление генетических и цитоло- гических карт хромосом ( карт групп сцепления )
Генетическая карта хромосомы – графическое изображение расстояний между генами в группах сцепления , выраженное в процентах кроссинговера ( схема взаимного расположения генов , находящихся в одной группе сцепления )
Цитологическая карта хромосомы – набор и локализация определённых генов в физических райлнах хромосом
- составление карт хромосом основано на учёте частот рекомбинаций ( процента кроссинговера ) между генами , относящимися к одной группе сцепления и постулате о линейном расположении генов
- Рассмотрим три гена А , В и С , которые наследуются сцеплённо , т. е. находятся в одной группе сцепления . В дигибридном скрещивании установлены частоты кроссинговера между ними : А – В – 5% , А – С –12% , В – С – 7% . Порядок расположения генов в хромосоме возможен только следующий 5% 7%
 А В С
А В С12%
- Наиболее полно изучена генетика и построены генетические карты хромосом у дрозофилы , имеющей 500 генов в 4 группах сцепления , кукурузы 400 генов в 10 группах сцепления , мыши , дрожжей , гороха пшеницы , томата , у человека из 23 пар хромосом выявлено всего 10 групп сцепления с небольшим числом генов в каждой группе
- У самцов дрозофилы кроссинговер практически не происходит , поэтому у дигетерозиготных самцов гены , расположенныне в одной хромосоме , обнаруживают полное сцепление , т. е. наследуются совместно ( такие самцы образуют только два родительских сорта гамет в равном количестве – АВ, ав и не приводят к образованию рекомбинантных потомков )
Основные положения хромосомной теории наследственности
- Сформулированы Т. Морганом и его школой в 1911 – 1920 годах
- Признаки организмов формируются под воздействием генов , находящихся в хромосомах ; различные хромосомы содержат неодинаковое число генов ; набор генов каждой из негомологичных хромосом уникален
- Каждый ген занимает в хромосоме определённое место ( локус ) ; аллельные гены занимают идентичные локусы гомологичных хромосом
- Гены расположены в хромосоме линейно по всей её длинне в определённой последовательности
- Гены одной хромосомы образуют группу сцепления и наследуются преимущественно совместно, сила сцепления находится в обратной зависимости от расстояния между генами ; число групп сцепления равняется числу хромосом в гаплоидном наборе
- Между гомологичными хромосомами может происходить обмен аллельными генами ( кроссинговер ) , частота которого прапорциональна расстоянию между генами
- Каждый биологический вид обладает спецефическим набором ( количество , форма , размер и расположение ) хромосом – кариотипом
Генетика и эволюция. Эволюционно-генетическая характеристика популяции . Популяционная генетика
- основоположники популяционной генетики : А.С. Серебровский , С.С. Четвериков , Н.П. Дубинин , К. Пирсон , Д. Харди , В. Вайнберг )
- Популяционная генетика - основа современной синтетической эволюционной теории ( С Э Т )
Популяционная генетика - раздел генетики , изучающий генетический состав ( генетическую структуру ) популяций
- Популяционная генетика изучает действие генетических законов на уровне популяций .