
Биологические и эпидемиологические эффекты облучения в малых дозах
| Вид материала | Документы |
- Эффекты облучения в твердых телах, 247.82kb.
- Алкоголь… Практически все мы употребляем алкоголь, кто-то очень редко, исключительно, 78.97kb.
- Статья Общие положения > Настоящие Требования к выдаче Свидетельства о допуске к работам, 138.38kb.
- Трии, служит тезис о принципиальной допустимости, приемлемости, а порой даже благотворности,, 411.15kb.
- Курсовая работа тема: «Алкалоиды и история их открытия», 101.03kb.
- Некоторые нерешенные вопросы в исследовании сенсорной системы у человека и ее обусловленности, 144.69kb.
- Тверской Государственный Университет Химико-Биолого-Географический факультет Кафедра, 103.37kb.
- Травы Гекаты в роли целителей, 2705.54kb.
- Дозы излучения и единицы измерения, 180.6kb.
- Болезнь, характеризующаяся непреодолимым влечением к наркотикам (напр., морфину), вызывающим, 19.62kb.
4. ЗАВИСИМОСТЬ ОТ АДАПТИРУЮЩЕЙ ДОЗЫ
Экспрессия адаптивного ответа особенно зависит от величины адаптирующей дозы при соответствующей [фиксированной] мощности (уровня) дозы. Обнаружено, что оптимальная доза для клеток кожи мышей (m5S) — 1–5 сГр (Sas95), для фибробластов человека — 10–20 сГр (HE22) (Cor94) или 3–5 сГр (HE19, HE57) (Ish96), а для лимфоцитов человека — 1–2 сГр (Cai90, Cor94, Kel91, Sha87, Sha89, Vij89, Wol88). Соответствующая зависимость для лимфоцитов человека приведена на рис. 3.
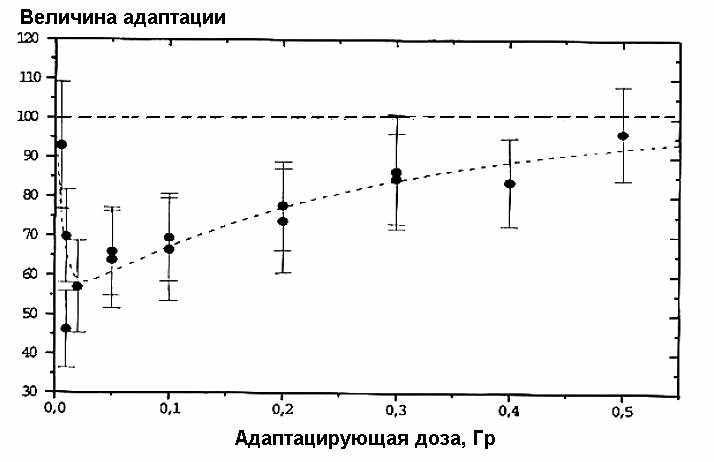
Рис. 3. Зависимость выраженности радиоадаптивного ответа в лимфоцитах человека от дозы адаптирующего облучения редкоионизирующей радиацией. По оси абсцисс — адаптирующая доза, Гр; по оси ординат — величина адаптации (снижение уровня аберраций хромосом). Объединенные данные (Sha87, Bai93).
5. ЗАВИСИМОСТЬ ОТ МОЩНОСТИ (УРОВНЯ) АДАПТИРУЮЩЕЙ ДОЗЫ
Соответствующие данные представлены на рис. 4 и 5
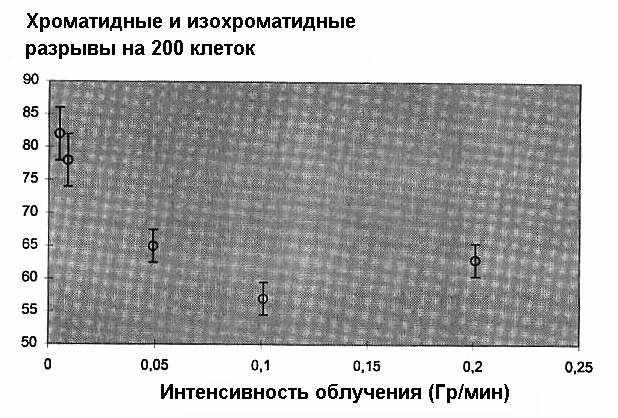
Рис. 4. Способность адаптирующей дозы в 0,01 Гр Х-излучения индуцировать радиационную адаптацию в зависимости от мощности дозы.
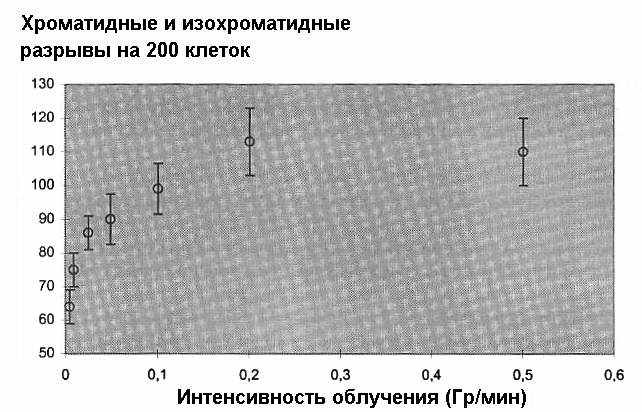
Рис. 5. Способность адаптирующей дозы в 0,5 Гр Х-излучения индуцировать радиационную адаптацию в зависимости от мощности дозы.
К удивлению, зависимости от мощности адаптирующей дозы для значений 0,01 Гр и 0,5 Гр оказались различными. В то время как в случае меньшей дозы количество повреждений ДНК повышается с увеличением мощности дозы, то для большей дозы характерна обратная картина (Sha89).
6. ЗАВИСИМОСТЬ ОТ ЛПЭ
Известно, что острое воздействие в малой дозе (0,005–0,2 Гр) редкоионизирующей радиации (Х- и -излучение) ведет к значительному снижению числа аберраций, индуцированных повреждающей дозой (0,4–4 Гр) (Sha94). К сожалению, плотноионизирующую радиацию очень редко использовали в качестве адаптирующей дозы. Исследования с отрицательными -мезонами показывают защитный эффект (Mar96), а протоны — нет (Kha91). Это может объясняться высоким повреждающим эффектом плотноионизирующей радиации, так как в данном случае имеется высокая локальная концентрация радикалов.
7. ЗАВИСИМОСТЬ ОТ ВЫБРАННОГО КРИТЕРИЯ ОЦЕНКИ РАДИОРЕЗИСТЕНТНОСТИ
Радиоадаптивный ответ определяли по различным конечным показателям (endpoints), среди которых — аберрации хромосом (Bor94, Cai90, Far93, Iku87, Iku89, Liu92, Mak90, San89, Sas95, Sha87, Sha89, Vij89, Vij95, Woj90, Woj92, Wol88), микроядра (Azz94, Cor94, Far93, Iku87, Iku89, Vij95, Woj97), ДР ДНК (Iku96), мутации (Cro90, Kel91, Rig93, San86, Sas95) и апоптоз (Ish96, Mar95, Mar96, Mar97, Mey94, Mus92, Sas95, Sin94, Ska94). Хотя степень экспрессии может отличаться для различных параметров, в случае аберраций хромосом и микроядер результаты для разных доноров оказались весьма сходными (рис. 6).
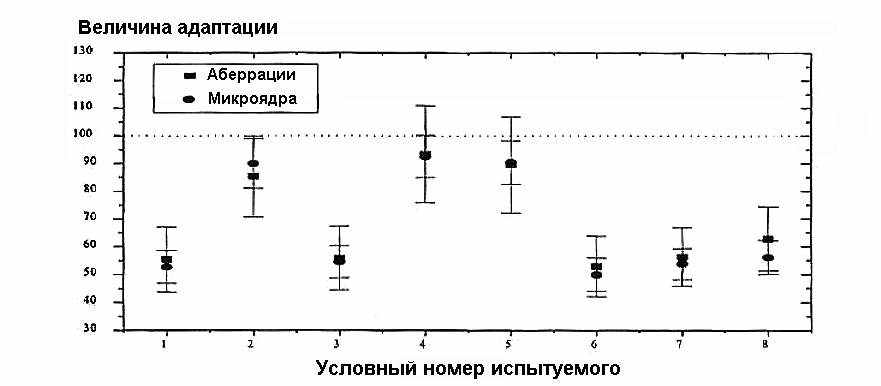
Рис. 6. Экспрессия радиоадаптивного ответа в лимфоцитах человека в зависимости от выбранного критерия оценки (аберрации, микроядра).
8. ПЕРВЫЙ МАТЕМАТИЧЕСКИЙ ПОДХОД
Как мы можем преуспеть в моделирования данных по адаптивному ответу, если рассматриваем очевидно разнородные экспериментальные результаты? Выше указано, что надо отличать причины такой вариабельности. Физиологические или генетические источники едва ли доступны для математических подходов, хотя ясные утверждения относительно возникновения радиоадаптации могут быть сделаны при рассмотрении целого массива данных. Намного более обещающей для математического анализа оказывается зависимость индукции процессов адаптации от физических факторов, типа дозы и уровня дозы, или же от природы излучения.
Но остается все еще открытым вопрос о том, реагируют ли клетки на радиационное воздействие более или менее уникальной программой генной экспрессии (в ответ на определенный сигнал), или же имеется широкий спектр возможного ответа согласно частоте и типу повреждений в пределах некоторых периодов времени. Другими словами, возникает вопрос, может ли отчетливый триггерный сигнал быть идентифицирован, или же нет. Что кажется ясным, так это то, что при радиоадаптации происходит синтез множества белков de novo. Но если величина вариаций в профиле генной экспрессии после облучения достаточно мала для выявления некоторого общего механизма радиоадаптации (он может быть и комплексным) в ответ на отчетливый триггерный сигнал, то вопрос все еще остается открытым.
С другой стороны, математические исследования могли бы также иметь некоторое значение с точки зрения внесения соответствующего вклада. Мы начали свое моделирование, основываясь на следующих рабочих гипотезах:
1) Действительно, имеется отчетливый триггерный сигнал при адаптации к ионизирующей радиации. Физические параметры облучения типа дозы, мощности дозы или природы излучения определяют вероятность появления триггерного сигнала. Как только сигнал прошел, клеточный статус трансформируется в преходящее «адаптированное» состояние с помощью более или менее инвариантных программ.
2) Вторая гипотеза относится к природе триггерного сигнала. Цель, на которую нужно воздействовать для того, чтобы радиоадаптация индуцировалась — это ДНК (на любом из ее уровней организации).
На основе этих предположений триггерный сигнал радиоадаптации является фокусом интересов при попытке найти возможные математические подходы, но не механизмом адаптации непосредственно. Характер триггерного сигнала может быть объяснен при помощи математических моделей.
Стратегия состоит в том, чтобы провести детальное микродозиметрическое исследование в соответствии с упрощенными микробиологическими предпосылками. Таким образом, «макроскопические» количества типа дозы, уровня дозы и характера излучения должны быть переведены в «микроскопические» категории типа количества энергии, поглощенной в определенных объемах в пределах указанных периодов времени или в категории частота / вероятности событий в указанных объемах. Тогда, после выяснения природы триггерного сигнала, вопрос переводится в задачу объяснить, каковы размеры мишеней, чтобы последовательность [ДНК] могла быть поражена, и какое минимальное количество энергии должно быть депонировано.
Уже можно сделать интересные предположения. Рассмотрим в качестве иллюстраций рис. 4 и 5. Удивительно, но мы видим отчетливый эффект уровня дозы для столь низких доз, как 0,01 Гр. Таким образом, объем мишени должен быть достаточно большим для того, чтобы многократные попадания были возможны (и, поэтому, осуществлялись их взаимодействия); при 0,01 Гр в качестве мишени должно выступать цельное клеточное ядро. Однако, если мощность дозы достаточно низка, радиоадаптация не происходит (рис. 4). Это позволяет сделать заключение об участии временной шкалы. «Порог» для установки триггерного сигнала составляет что-то около 10 сГр в мин (рис. 4).
Основываясь на суммарном диаметре мишени (среднее ядро клетки млекопитающих), можно сделать вывод о том, что необходимый интервал времени составляет приблизительно несколько сотен миллисекунд между двумя последующими событиями (Х-излучение). Это дольше, чем типичная временная диффузия через ядро клетки, но короче, чем время жизни вторичных радикалов, подобных H2O2 (см. Ada80).
Если интервал времени становится длиннее (уменьшающаяся мощность дозы), то триггерный сигнал исчезает. Оба наблюдения (размер мишени и временная шкала) привели нас к предположению, что триггерный сигнал генерируется через воздействие вторичных радикалов. Они, однако, недостаточно реактивны для причинения значительного повреждения ДНК. Таким образом, можно судить о возможных механизмах, вновь возвратившись к высоко реактивным радикалам, подобным радикалу ОH (гидроксил). Такие механизмы были описаны и, вероятно, они могут наблюдаться в пределах ядра клетки (Kie89).
В то же время, рис. 5 (снижение радиоадаптации с увеличением мощности дозы) показывает, что существуют дополнительные процессы, превалирующие при высоких дозах над механизмами, индуцирующими адаптивный ответ при сравнительно низких дозах. Подтверждение зависимости от мощности (уровня) дозы при дозах столь высоких, как 0,5 Гр, дало возможность заключить, что эти дополнительные механизмы играют роль для мишеней много меньшего размера. Техника нелинейной регрессии применительно к соответствующим хорошо разработанным моделям позволяет сделать вывод о диаметре мишени приблизительно в 0,8 мкм105.
Таким образом, мы видим два противодействующих процесса, которые должны быть сбалансированы для возникновения триггерного сигнала, причем каждый из них связан с различными мишенями.
9. ИНТЕРПРЕТАЦИЯ МАТЕМАТИЧЕСКИХ РАСЧЕТОВ
Несмотря на начало наших математических исследований, хотелось бы поспекулировать относительно интерпретации результатов. Принимая во внимание, что предположение о вторичных радикалах, являющихся посредниками триггерного сигнала, кажется верным для низких доз, они не обязательно участвуют в опосредовании дополнительных процессов (counteracting process), поскольку размер мишени и интервал времени между двумя последующими попаданиями слишком малы, чтобы объяснить эффект мощности дозы при более высоких дозах (рис. 5). Вместо этого, по-видимому, главную роль в дополнительных процессах при высоких дозах играют первичные радикалы. Однако остается открытым вопрос, на какие домены ДНК они воздействуют, исходя из нашей второй рабочей гипотезы. В соответствии с размером мишени приблизительно в 0,8 мкм (полученным в наших расчетах), мы заключаем, что ответ может быть связан с кластерами репликонов (организационная единица репликации ДНК в эукариотических клетках), которые имеют сопоставимый размер (Cle83).
Если некоторый типы повреждений ДНК внутри кластера репликонов инициируют репликацию цельного кластера, она приостанавливается, пока повреждения не репарируются. Таким образом, отсрочка локальной инициации репликации может подавлять триггерный сигнал к радиоадаптации. Это определяет центральную роль репликации для возникновения адаптивного ответа.
Действительно, существуют экспериментальные доказательства критической роли облучения в S-фазе, так как установлено, что клетки должны пройти S-фазу для проявления адаптивного ответа (Rya98). В работе (Rya98) выдвигается гипотеза, согласно которой начальные повреждения, вызванные некоторой низкой дозой, должны быть «восприняты» и, «в пути», «обработаны» клеткой. Эта «обработка» не может быть осуществлена когда-либо на протяжении клеточного цикла, но только в течение некоторых событий, происходящих в S-фазе.
Интересно, что мы можем видеть определенные параллели с SOS ответом у E. coli, где пострепликативные промежутки (то есть, основные повреждения, «процессируемые» репликацией) инициируют механизмы защиты, хотя регулирование процессов в клетках млекопитающих, конечно, является намного более сложным.
ЛИТЕРАТУРА
1. [Ada80] G.E. Adams, D.G. Jameson, Time effects in molecular radiation biology, Radiation Environmental Biophysics, 17, 95-114, 1980.
2. [Azz94] E.I. Azzam, G.P. Raaphorst and R.E.J. Mitchel, Radiation-induced adaptive response for protection against micronucleus formation and neoplastic transformation in C3H 10T1/2 mouse embryo cells, Radiation Research, 138, S28-S31, 1994.
3. [Bor94] P.M. Borodin, M. Inouye, S.-I. Oda, T. Ikushima, Y. Takagishi and H. Yamamura, Radioadaptive response in primary mouse spermatocytes revealed by analysis of synaptonemal complexes, Mutation Research, 310, 151-156, 1994.
4. [Bos89] A. Bosi and G. Olivieri, Variability of the adaptive response to ionizing radiations in humans, Mutation Research, 211, 13-17, 1989.
5. [Cai90] L. Cai and S.-Z. Liu, Induction of Cytogenetic Adaptive Response of Somatic and Germ Cells in vivo and in vitro by Low-Dose X-Irradiation, International Journal of Radiation Biology, 58, 187-194, 1990.
6. [Cor94] F. Cortes, I. Dominguez, M.J. Flores, J. Pinero, J.C. Mateos, Differences in the Adaptive Response to Radiation Damage in Go Human Lymphocytes Conditioned with Hydrogen Peroxide or Low-Dose X-Rays, Mutation Research, 311, 157-163, 1994.
7. [Cro90] N.E.A. Crompton, B. Barth and J. Kiefer, Inverse dose-rate effect for the induction of 6-thioguanine-resistant mutants in Chinese hamster V79-S cells by 60Co -rays, Radiation Research, 124, 300-308, 1990.
8. [Far93] Z. Farooqi and P.C. Kesavan, Low-Dose Radiation-Induced Adaptive Response in Bone Marrow Cells of Mice, Mutation Research, 302, 83-89, 1993.
9. [Gad98] P.K. Gadhia, Possible age-dependent adaptive response to a low dose of X-rays in human lymphocytes, Mutagenesis, 13, 151-152, 1998.
10. [Iku87] T. Ikushima, Chromosomal responses to ionizing radiation reminiscent of an adaptive response in cultured Chinese hamster cells, Mutation Research, 180, 215-221, 1987.
11. [Iku89] T. Ikushima, Radio-Adaptive Response: Characterization of a Cytogenic Repair Induced by Low-Level Ionizing Radiation in Cultured Chinese Hamster Cells, Mutation Research, 227, 241-246, 1989.
12. [Iku96] T. Ikushima, H. Aritomi and J. Morisita, Radioadaptive response: efficient repair of radiation-induced DNA damage in adapted cells. Mutation Research, 358, 193-198, 1996.
13. [Ish96] K. Ishii and M. Watanabe, Participation of gap-junctional cell communication on the adaptive response in human cells induced by low dose of X-rays, International Journal of Radiation Biology, 69, 291-299, 1996.
14. [Kal94] I. Kalina, H. Konecna and N. Racekova, Adaptive response to ionizing radiation in normal and aneuploid human lymphocytes, Folia Biologica, 40, 110-123, 1994.
15. [Kal97] I. Kalina, G. Nemethova, Variability of the Adaptive Response to low Dose Radiation in Peripheral Blood Lymphocytes of Twins and Unrelated Donors, Folia Biologica, 43/2, 91-95, 1997.
16. [Kel91] K.T. Kelsey, A. Memisoglu, D. Frenkel and H.L. Liber, Human lymphocytes exposed to low doses of X-rays are less susceptible to radiation-induced mutagenesis, Mutation Research, 263, 197-201, 1991.
17. [Kha91] E.K. Khandogina, G.R. Mutovin, S.V. Zvereva, A.V. Antipov, D.O. Zverev and A.P. Akifyev, Adaptive response in irradiated human lymphocytes: radiobiological and genetical aspects, Mutation Research, 251, 181-186, 1991.
18. [Kie89] J. Kiefer, Biologische Strahlenwirkung, Birkhauser Verlag, Basel, 1989.
19. [Liu92] S.Z. Liu, L. Cai and S.Q. Sun, Induction of a cytogenetic adaptive response by exposure of rabbits to very low dose-rate -radiation, International Journal of Radiation Biology, 62, 187-190, 1992.
20. [Mak90] T. Makinodan and S.J. James, T cell potentiation by low dose ionizing radiation: possible mechanisms, Health Physics, 59/1, 29-34, 1990.
21. [Mar95] B. Marples and M.C. Joiner, The elimination of low-dose hypersensitivity in Chinese hamster V79-379A cells by pretreatment with X-rays of hydrogen peroxide. Radiation Research, 141,160-169, 1995.
22. [Mar96] B. Marples and K.A. Skov, Small doses of high-linear energy transfer radiation increase the radioresistance of Chinese hamster V79 cells to subsequent X-irradiation, Radiation Research, 146,382-387, 1996.
23. [Mar97] B. Marples, P. Lambin, K.A. Skov and M.C. Joiner, Low dose hyper-radiosensitivity and increased radioresistance in mammalian cells, International Journal of Radiation Biology, 71, 721-735, 1997.
24. [Mey94] M. Meyers, J. McCaffrey, E. Odegaard and D.A. Boothman, X-ray-inducible transcripts and altered cell cycle regulation are involved in adaptive survival responses in human cells, Radiation Research, 141, 119-120, 1994.
25. [Mus92] R. J. Muschel, H.B. Zhang, G. Iliakis, and W.G. McKenna, Effects of ionizing radiation on cyclin expression in HeLa cells, Radiation Research, 132, 153-157, 1992.
26. [01184] G. Olivieri, J. Bodycote and S. Wolff, Adaptive response of human lymphocytes to low concentrations of radioactive thymidine, Science, 223, 594-597, 1984.
27. [Rig93] O. Rigaud, D. Papadopoulo and E. Moustacchi, Decreased deletion mutation in radioadapted human lymphocytes. Radiation Research, 133, 94-101, 1993.
28. [Rya98] N.I. Ryabchenko, M.M. Antoshchina, E.V. Fesenko, T.I. Ivanova, T.V. Kondrashova and V.A. Nasonova, Cytogenetic adaptive response in cultured human lymphocytes: dependence on the time of exposure to adapting and challenging doses of gamma-rays, Mutat. Res., 418, 7-19, 1998.
29. [San86] B.J.S. Sanderson and A.A. Morley, Exposure of human lymphocytes to ionizing radiation reduces mutagenesis by subsequent ionizing radiation, Mutation Research, 164, 347-351, 1986.
30. [San89] K. Sankaranarayanan, A. von Duyn, M.J. Loos and A.T. Natarajan, Adaptive response of human lymphocytes to low-level radiation from radioisotopes of X-rays, Mutation Research, 211, 7-12, 1989.
31. [Sas95] M.S. Sasaki, On the reaction kinetics of the radioadaptive response in cultured mouse cells, International Journal of Radiation Biology, 68, 281-291, 1995.
32. [Sas95] M.S. Sasaki, On the reaction kinetics of the radioadaptive response in cultured mouse cells. International Journal of Radiation Biology, 68, 281-291, 1995.
33. [Sha87] J.D. Shadley, V. Afzal and S. Wolff, Characterization of the adaptive response to ionizing radiation induced by low doses of X-rays to human lymphocytes, Radiation Research, 111,511-517, 1987.
34. [Sha89] J.D. Shadley and J.K. Wiencke, Induction of the adaptive response by X-rays is dependent on radiation intensity. International Journal of Radiation Biology, 56, 107-118, 1989.
35. [Sin94] B. Singh, J.E. Arrand and M.C. Joiner, Hypersensitive response of normal human lung epithelial cells at low radiation doses, International Journal of Radiation Biology, 65, 457-464, 1994.
36. [Ska94] L.D. Skarsgard, M.W. Skwarchuk and B.G. Wouters, The survival of asynchronous V79 cells at low radiation doses: modeling the responses of mixed cell populations, Radiation Research, 138, S72-S75, 1994.
37. [Sko94] K.A. Skov, H. Zhou and B. Marples, The effect of two topoisomerase inhibitors on low-dose hypersensitivity and increased radioresistance in Chinese hamster V79 cells, Radiation Research, 138, S117-120, 1994.
38. [UNS94] United Nations Scientific Committee on the Effects of Atomic Radiation, Sources and effects of ionizing radiation, New York, United Nations, 1994.
39. [Vij89] Vijayalaxmi and W. Burkart, Resistance and cross-resistance to chromosome damage in human blood lymphocytes adapted to bleomycin, Mutation Research, 211, 1-5, 1989.
40. [Vij95] Vijayalaxmi, B.Z. Leal, T.S. Deahl and M.L. Meltz, Variability in adaptive response to low dose radiation in human blood lymphocytes: consistent results from chromosome aberrations and micronuclei, Mutation Research, 348, 45-50, 1995.
41. [Wie86] J.K. Wiencke, V. Afzal, G. Olivieri and S. Wolff, Evidence that the [3H] - thymidine-induced adaptive response of human lymphocytes to subsequent doses of X-rays involves the induction of a chromosomal repair mechanism, Mutagenesis, 1, 375-380, 1986.
42. [Woj90] A. Wojcik and H. Tuschi, Indications of an Adaptive Response in C57BL Mice Pre-Exposed in vivo to Low Doses of Ionizing Radiation, Mutation Research, 243, 67-73, 1990.
43. [Woj92] A. Wojcik, K. Bonk, W.-U. Muller, C. Streffer, U. Weissenborn and G. Obe, Absence of adaptive response to low doses of X-rays in preimplanation embryos and spleen lymphocytes of an inbred mouse strain as compared to human peripheral lymphocytes: a cytogenetic study, International Journal of Radiation Biology, 62, 177-186, 1992.
44. [Woj97] M. Wojewodzka, M. Kruszewski and I. Szumiel, Effect of signal transduction inhibition in adapted lymphocytes: micronuclei frequency and DNA repair. International Journal of Radiation Biology, 71, 245-252, 1997.
45. [Wol88] S. Wolff, V. Afzal, J.K. Wiencke, G. Olivieri and A. Michaeli, Human lymphocytes exposed to low doses of ionizing radiations become refractory to high doses of radiation as well as to chemical mutagens that induce double-strand breaks in DNA, International Journal of Radiation Biology, 53, 39-48, 1988.
46. [Yon96] M. Yonezawa, J. Misonoh and Y. Hosokawa, Two types of X-ray-induced radioresistance in mice: presence of 4 dose ranges with distinct biological effects. Mutation Research, 358, 237-243, 1996.
47. [You89] J.H. Youngblom, J.K. Wiencke and S. Wolff, Inhibition of the adaptive response of human lymphocytes to very low doses of ionizing radiation by the protein synthesis inhibitor cycloheximide, Mutation Research, 227, 257-261, 1989.
48. [Zha95] L. Zhang, Cytogenetic adaptive response induced by pre-exposure in human lymphocytes and marrow cells of mice, Mutation Research, 334, 33-37, 1995.
Ответ лимфоцитов популяции человека и антиоксидантная защита их от воздействия радиации в низких дозах {58}
G.J. Koteles*, I. Bojtor*, G. Bognar* and M. Otos**
* Национальный исследовательский институт радиобиологии и радиационной гигиены им. Фредерика Жолио-Кюри, Будапешт
** Атомное объединение, Paks, Венгрия
Koteles G.J., Bojtor I., Bognar G., Otos M. Lymphocyte response in human population and its antioxidant protection against low doses of ionizing radiation. In: “The Effects of Low and Very Low Doses of Ionizing Radiation on Human Health”, ed. by WONUC. 2000. Elsevier Science B.V. P. 295–304.
G.J. Koteles*, I. Bojtor*, G. Bognar* and M. Otos**
* Frederic Joliot-Curie National Research Institute for Radiobiology and Radiohygiene, Budapest, Hungary
** Atommed Ltd., Paks, Hungary