
О. А. Громова Допустить к защите
| Вид материала | Диплом |
- Юрия Иосифовича Громова. Фестиваль-конкурс, 83.33kb.
- Г. Г. Исаев (гл редактор), Е. Е. Завьялова, Т. Ю. Громова, 4887.01kb.
- Тематическое и поурочное планирование по физике к учебнику С. В. Громова, Н. А. Родиной, 1532.4kb.
- Архангельский Государственный Технический Университет Институт экономики, финансов, 947.48kb.
- Я. И. Кузьминову Код абитуриента : заявление прошу допустить меня к участию в конкурс, 31.08kb.
- Кафедра Информационных Систем и Технологий. Сдана на проверку Допустить к защите 2007, 625.08kb.
- И. И. Ползунова Факультет руководящих работников и специалистов «Допустить к защите», 782.6kb.
- И. И. Ползунова Факультет руководящих работников и специалистов «Допустить к защите», 785.97kb.
- Г. Р. Латфуллин, О. Н. Громова Организационное поведение Авторский коллектив:, 4898.59kb.
- А. М. Горького Институт управления и предпринимательства Кафедра управления Формирование, 530.85kb.
Содержание фосфора и магния в головном мозге белых крыс контрольной и экспериментальной групп
| показатели | x1 | x2 | d1 | d2 | t | Р | % |
| Фосфор мкг/кг | | ||||||
ЦИЛДК | 279450 | 517070 | 7969,99 | 4716,3 | 25,7 | Р< 0,001 | +85,03 |
ГТ | 421910 | 284370 | 9973,4 | 1669,9 | 13,6 | P< 0,001 | -32,5 |
ОЛ | 411390 | 317960 | 115092,2 | 5148,4 | 0,8 | P> 0,05 | -22,7 |
| Магний мкг/кг | | ||||||
ЦИЛДК | 39458 | 21960 | 345,183 | 14,4 | 32,1 | P 0,001 | -44,34 |
ГТ | 27430 | 29450 | 133,2 | 166,29 | 9,5 | P 0,001 | +7,36 |
ОЛ | 20402 | 25904 | 437,4 | 364,2 | 9,7 | P 0,001 | +27 |
Таблица 2
Содержание кобальта и цинка в головном мозге белых крыс контрольной и экспериментальной групп
Показатели | x1 | x2 | d1 | d2 | t | P | % |
| Кобальт мкг/кг | | ||||||
ЦИЛДК | 16,29 | 12,84 | 0,3 | 0,07 | 10,2 | 0,001 | -21 |
ГТ | 1,19 | 4,98 | 0,09 | 0,2 | 14,9 | 0,001 | 318,4 |
ОЛ | 4,44 | 2,07 | 0,17 | 0,39 | 13,2 | 0,001 | -53,3 |
Цинк мкг/кг | | ||||||
ЦИЛДК | 2881,4 | 4233,3 | 108,7 | 62,7 | 10,8 | 0,001 | 47 |
ГТ | 4191 | 2210,9 | 57,1 | 27,5 | 31,2 | 0,001 | -47,2 |
ОЛ | 3145,3 | 4033,1 | 11,9 | 42 | 0,00005 | 0,05 | 28,2 |
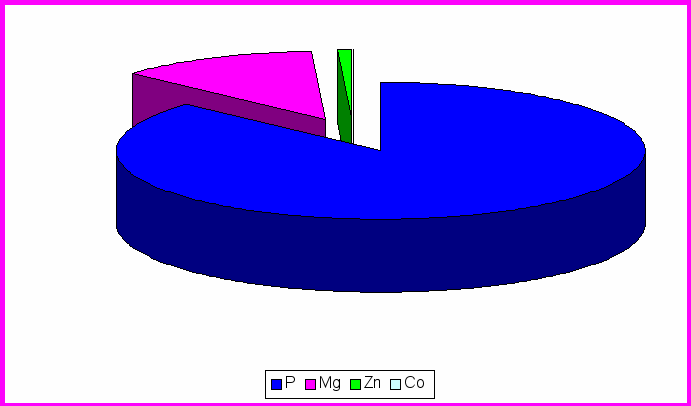
Рис.2. Содержание микроэлементов в центральной извилине лобной доли коры у контрольной группы крыс
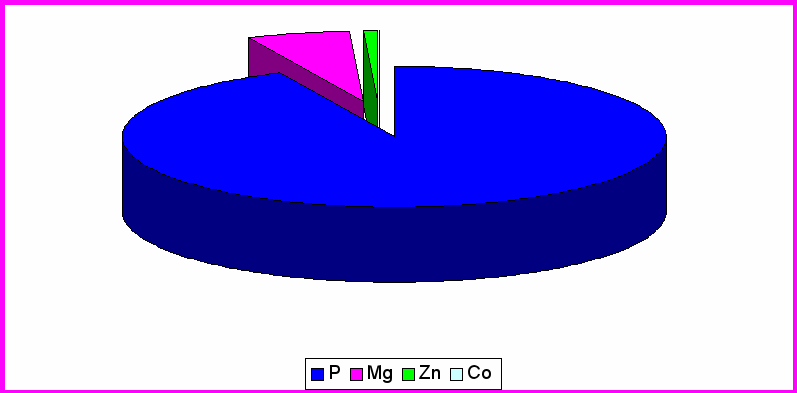
Рис. 2. Содержание микроэлементов в гипоталамусе контрольной группы крыс
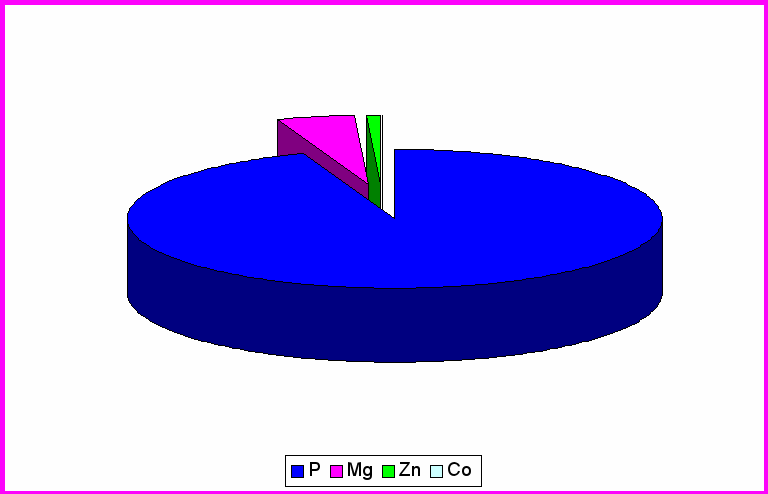
Рис.2. Содержание микроэлементов в обонятельных луковицах контрольной группы крыс
3.2. Распределение макро- и микроэлементов в различных отделах головного мозга при влиянии MgB6
Опытная группа крыс (6 особей) получала внутрижелудочно MgB6 в течение 10 дней в дозе 5 мл/кг массы тела.
Согласно таблице 1 видно, что среднее содержание фосфора в различных структурах головного мозга распределяются следующим образом: ЦИЛДК-517070мкг/кг; ГТ- 284370мкг/кг; ОЛ- 317960мкг/кг. Из рисунка 3 выяснили, что большее процентное содержание во всех трёх отделах приходится на фосфор (ЦИЛДК-95%; ГТ-90%; ОЛ-91%)
Содержание Mg составило в ЦИЛДК-21960мкг/кг; ГТ-29450мкг/кг; ОЛ- 25904мкг/кг. Из рисунка 3 видно, что процентное содержание Mg в ГТ (9%) увеличивается по сравнению с ЦИЛДК (4%) и ОЛ (7%).
Как выяснилось из таблицы 2, на которой представлены данные по содержанию Co и Zn в отделах головного мозга экспериментальной группы животных. В ЦИЛДК содержится 12,84мкг/кг Co и 4233,3мкг/кг Zn. В ГТ содержание Co составило 4,98мкг/кг, а Zn 2210,9мкг/кг. В ОЛ (табл.2) содержание Co составляет 2,07 ,а Zn 4033,1мкг/кг. Из рис.3 видно, что Co , Zn составляют меньшее количество % , во всех трёх отделах головного мозга, т.к. концентрация в организме микроэлементов определяется тысячными долями грамма, но без их участия не могут происходить сложные биологические процессы. С их помощью поддерживается работа ферментативных систем, регулируется температура тела. (Громова, Кудрин 2001).
Таким образом метод статистической обработки показал, что отличия в распределении МЭ в различных отделах головного мозга присутствует, следовательно данные нужно сравнить с контролем.
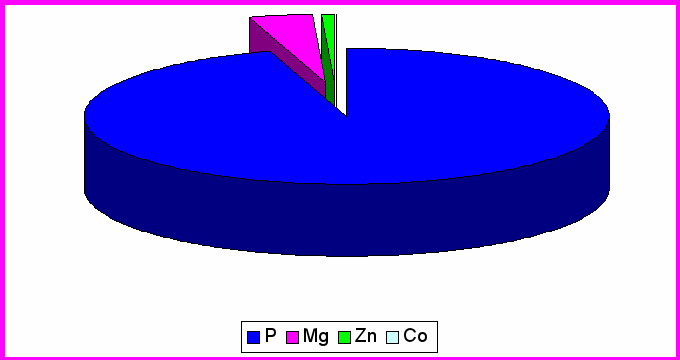
Рис.3. Влияние MgB6 на содержание микроэлементов в центральной извилине лобной доли коры
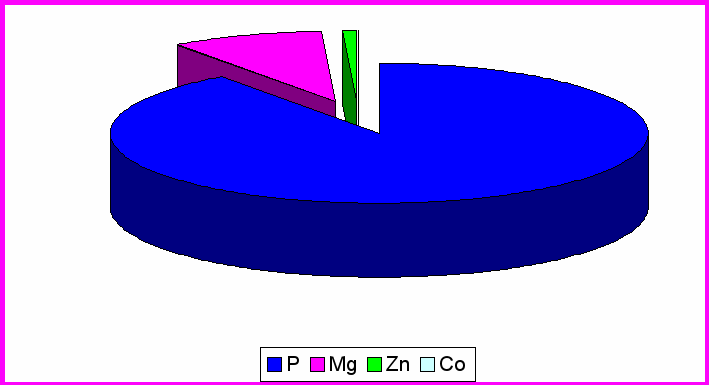
Рис. 3. Влияние MgB6 на содержание микроэлементов в гипоталамусе
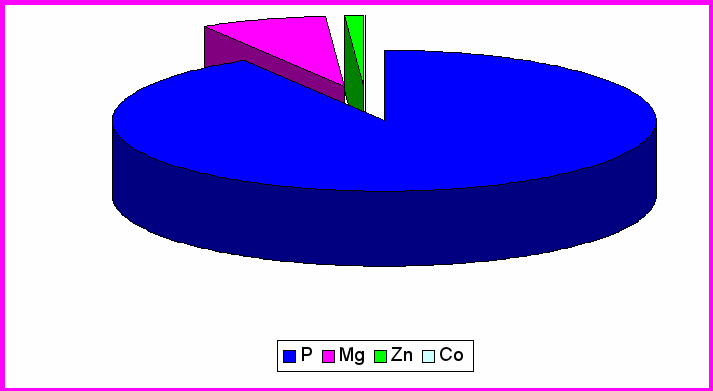
Рис.3. Влияние MgB6 на содержание микроэлементов в обонятельных луковицах
Сравнение содержания микро- и макроэлементов в головном мозге контрольной и экспериментальной группы животных
Чтобы выявить влияние пищевой добавки MgB6 на содержание ионов в различных отделах головного мозга, мы сравнили полученные данные 2 и 3 группы животных. Из таблицы 1 видно, в ЦИЛДК у животных получавших MgB6 содержание фосфора выросло на 85% (P < 0,001); в ГТ уменьшилось на 32,5 % (P < 0,001). Эти изменения в ОЛ оказались
недостоверными в ЦИЛДК у животных получавших MgB6.
Содержание магния уменьшилось на 44,3 % (P < 0,001), в ГТ возросло на 7,3% , ОЛ возросло на 27 % (P < 0,001).
Из таблицы 2 видно, что в ЦИЛДК у экспериментальных животных содержание Co уменьшается на 21% (P < 0,001), в ГТ выросло 318% (P < 0,001) , в ОЛ уменьшилось на 53 % (P < 0,001). В ЦИЛДК у экспериментальных животных содержание Zn возросло на 47% (P < 0,001) , в ГТ уменьшилось на 47 % (P < 0,001). Эти изменения в ОЛ оказались недостоверными.
Мы сравнили достоверность распределения МЭ в различных структурах головного мозга контрольной и экспериментальной групп животных (рис.4)видно, что 10-ти дневное последовательное введение препарата достоверно приводит к увеличению содержания фосфора в ЦИЛДК и уменьшению содержания магния в ЦИЛДК, содержание кобальта и цинка уменьшается.
В ГТ введение препарата достоверно приводит к уменьшению содержания фосфора (3 %), к увеличению содержания магния (3 %), кобальта (0,0012%). В ОЛ введение препарата достоверно приводит к уменьшению содержания фосфора (4%), кобальта (0,0005%), к увеличению содержания магния (2%), цинка (0,5%).
Таким образом из результатов работы видно, что при введении MgB6 концентрация магния увеличивается в ГТ и ОЛ, а в ЦИЛДК уменьшается.
Механизм распределения пищевой добавки не изучен, хотя приём этой пищевой добавки должен быть согласован с врачом, т.к. пищевая добавка значительно изменяет ионный состав ГТ, ЦИЛДК, ОЛ.
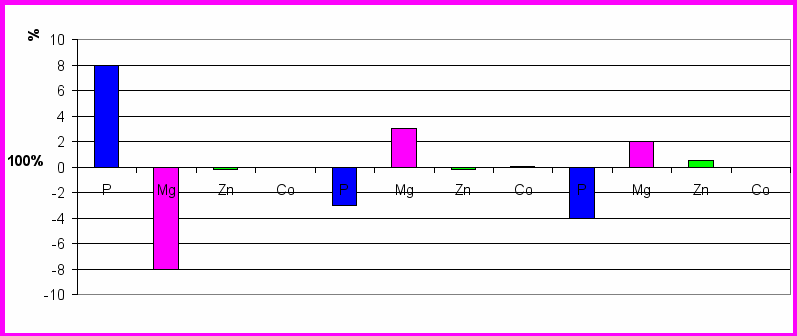
ЦИЛДК ГТ ОЛ
Рис.5 Сравнение содержания микроэлементов в головном мозге контрольной и экспериментальной групп крыс
ГЛАВА IV. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Для исследования были взяты 18 белых крыс, массой 180 2,6(г), которые содержались в изолированных клетках в условиях вивария на стандартном пищевом рационе при свободном доступе к воде. Все животные были разделены на 3 группы:
1 группа – интактные животные. Группа необходима для того, чтобы исключить влияние на состав МЭ в головном мозге крыс условий жизни и питания в виварии.
2 группа – контрольная группа животных, получившие внутрижелудочно диситиллированую воду. Эта группа необходима для исключения влияния механического раздражения рецепторов слизистой желудка на микроэлементный состав головного мозга.
3 группа – экспериментальные животные, которые в течение 10-ти дней внутрижелудочно получали MgB6 в дозе 5мл/кг.
На исследование брали следующие структуры головного мозга: ЦИЛДК, ГТ, ОЛ. Затем при помощи метода «мокрого озоления » эти структуры подготавливали к исследованию на масс- спектрометре с индуктивно - связанной аргоновой плазмой. При помощи этого метода были выделены микроэлементы , входящие в состав головного мозга, из которых для исследования были взяты P, Mg, Zn, Co. Содержание этих МаЭ и МЭ определялось у каждой группы. Для исследования были взяты именно эти МЭ т.к они играют неотъемлемую роль в поддержании минерального гомеостаза нейронов.
Нейрохимические эффекты кобальта связаны с тем, что кобальт может стабилизировать разнообразные рецепторы, в частности регулирует адрено и норадренолитические эффекты.(M.Antony,1995). Комплекс Co с АТФ оказывает антигипоксическое действие и нейропротекторное при диффузной хронической церебральной анемии Co при поступлении в цитоплазму нейронов активизирует HIF- 1. Он играет главную роль в антистрессорной защите мозга, предупреждение апоптоза и механизмах клеточного цикла и роста нейронов (О.А. Громова; А.В. Кудрин ,2001).
Содержание Zn в тканях серого вещества мозга от 150-200мк моль(Weiss J.H,. 1993)в терминальных отростках нейронов концентрация Zn в 2,5 – 3 раза выше (Frederickson 1983).Ингибитор плазмогена подавляет Zn зависимый апоптоз нейронов. Цинк стабилизирует гематоэнцефалический барьер, при интоксикации тяжёлых Ме: Pb, Hg, т.е. Zn может быть , как антагонистом тяжёлых Ме, так и профилактическим средством в группах риска. В неврологической практике дефицит Zn встречается чаще, чем избыток (О.А. Громова; А.В. Кудрин 2001).
Фосфор принимает участие в построении нервной и костной ткани. Фосфорные соли калия, натрия участвуют в создании буферности крови (Саноцкий И.В., 1979).
Магний черезвычайно важен для нормального функционирования нервной системы одним из важнейших эффектов магния это торможение процессов возбуждения в коре головного мозга и связанная с этим реализация наркотического, снотворного, анальгетического, противососудистого эффектов. Доказана роль магния в качестве модулятора эффектов возбуждающие аминокислоты в ЦНС. Магний своего рода изоляционный материал на пути проведения нервного импульса. (De Bloc 1999) на большом экспериментальном уровне показали зависимость выраженности элементарной готовности в коре головного мозга от уровня ионов Mg. Дефицит ионов магния повышает чувствительность организма к нейроинфекциям (Takeda A, 2001).
В ходе исследования были получены следующие результаты: изменений в содержании МЭ у интактной группы животных (внутрижелудочное введение диситиллированной воды) выявлено не было. Это свидетельствует о том ,что внутрижелудочное введение воды не вызывает изменений в содержании МЭ в головном мозге крыс. При 10-ти дневном последовательном введении MgB6 содержание фосфора в ЦИЛДК увеличилось на 8 %. Также при этом способе введения MgB6 наблюдалось снижение содержания магния в ЦИЛДК, но увеличение его в ГТ и ОЛ. Наблюдается снижение содержания Co в ОЛ и в ЦИЛДК. Это отрицательно сказывается на организме т.к. Co участвует в процессах кроветворения и регенерации (Авцын А.П. с соавт., 1991), стимулирует лейкопоэз (Соловьёв 1997).Также можно видеть снижение содержания цинка в ГТ и его увеличение в ОЛ. Дефицит Zn встречается значительно чаще, чем избыток. При кровоизлияниях происходят большие потери цинка, а введение этой добавки усиливает его выход. Неврологическими признаками дефицита цинка являются, по данным (Frederickson,1983)депрессии, эмоциональная лабильность (Кудрин П.В. с соавт.,2000).
В ходе исследования было выявлено неравномерное распределение МЭ в структурах головного мозга крыс.
Распределение МЭ в структурах ЦНС неодинаково (В.С. Райцес, 1981), это связанно с особенностями обменных процессов (A.Takeda, 2001). Ряд учёных считают, что содержание МЭ выше, в участках мозга с наиболее интенсивной деятельностью (Громов Л.А.,1992).
Головной мозг, особенно кора больших полушарий, лидирует по содержанию большинства МЭ (Москалёв Ю.И.,1985). Так в коре больше всего содержится меди,титана, но мало содержится кремния, алюминия, магния (R.A. Goyer et al, 1995).
Гипоталамус содержит меньше МЭ, чем подкорковые и стволовые структуры. Количество МЭ в передних отделах гипоталамуса выше, чем в задних (M.Antony, 1995).
В ходе исследования литературные данные были подтверждены. В эксперименте, как и в литературных данных, наибольшее содержание МЭ наблюдалось в ЦИЛДК, затем в ГТ, наименьшее содержание МЭ наблюдалось в ОЛ.
В связи с результатами можно сделать следующее заключение:
Принимать пищевую добавку MgB6 нужно в строго указанном количестве, т.к. высокие дозы пищевой добавки влияют на содержание МЭ в нейронах головного мозга, нарушая их функциональное состояние.