
Федеральная программа книгоиздания России Рецензенты: канд психол наук С. А. Исайчев, доктор биол наук И. И. Полетаева Равич-Щербо И. В. и др. Р12
| Вид материала | Программа |
- Вестник балтийской педагогической академии вып. 94. – 2010 г. Актуальные проблемы нравственного, 2431.92kb.
- Рецензенты: профессор, доктор психол наук Филонов Л. Б., вед науч сотрудник, канд психол, 2609.63kb.
- Научный выпуск вестник балтийской педагогической академии вып. 29. – 2000 г. Поиск, 1745.18kb.
- Общеобразовательная программа дошкольного образования Авторский коллектив, 5619.19kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Введенским Игорем Витальевичем Рецензенты доктор психол наук В. А. Лабунская канд психол, 375.9kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- Пояснительная записка, 12621.4kb.
- В организации совместных отношений, 1137.14kb.
179
Основное ограничение метода заключается в том, что генетически идентичные родители могут создавать в своих семьях сходную среду, в которой взаимодействия в диадах ребенок Х МЗ родитель и ребенок Х МЗ дядя (тетя) будут весьма похожими, а у ДЗ родителей со своими детьми и племянниками будет такое же средовое и генетическое сходство, как у родителей и детей в обычной семейной ситуации. Поэтому данный вариант близнецового метода может успешно решать некоторые частные вопросы, однако для разделения генетической и средовой дисперсии признака он имеет меньшую разрешающую способность, чем другие методы психогенетики [363].
Метод контрольного близнеца (взаимоконтроля близнецов, контроля по партнеру, близнеца-свидетеля) основан на том, что МЗ близнецы, — генетически одинаковые люди, имевшие общую эмбриональную и в обычных условиях семейную среду, — являются идеальным контролем друг к другу. Если группу МЗ близнецов разделить на две выборки так, чтобы в каждую вошли по одному близнецу из каждой пары, то исследователь получит уникальную ситуацию — две выборки, уравненные и по генетической конституции, и по основным средовым (общесемейным) параметрам. Далее можно, например, одну группу тренировать, а другую — нет или тренировать их в разном возрасте, как это было сделано в работе А. Гезелла, впервые применившего данный метод и прослеживавшего вместе с коллегами развитие одной женской пары МЗ близнецов с первых месяцев жизни до 14 лет [272, 273, 274, 275]. Они получили материал, свидетельствующий о зависимости эффективности обучения (двигательного и речевого) от возраста. Одновременно можно оценить и длительность удержания эффекта тренировки: поскольку в раннем детстве созревание существенно определяется генетической программой, тип и темп его у МЗ близнецов в основном одинаковы. В упомянутых работах А. Гезелла и его сотрудников показано, например, что разница в речевом развитии, стимулированном тренировкой в возрасте 1,5-2 лет, стерлась в течение трех месяцев.
Шведский исследователь А. Незлунд [цит. по: 434] оценил таким методом разные способы обучения чтению и показал, что одного, оптимального для всех способа нет, их эффективность зависит от уровня интеллектуального развития ребенка.
В упоминавшихся (см. Введение) работах Московского Медико-генетического института этот метод (5 пар МЗ близнецов 5-6 лет) тоже использован для анализа не только эффекта обучения, но и конструктивной деятельности. Показано, что обучение при помощи метода «моделей» эффективнее, чем простое копирование: не только улучшается сама конструктивная деятельность, но и перестраиваются другие психические функции ребенка. Позже, в 1956 г., была опубликована работа А.Р. Лурия и Ф.Н. Юдович о развитии речи, где МЗ близнецы также служили контролем друг к другу [ 102].
180
Р. Пломин с соавторами [363] отмечал, что таких работ очень мало — «менее дюжины», но и их, тем не менее, делят на три группы. В первую входят наиболее ранние работы, в которых исследуется взаимодействие между созреванием и тренировкой, при этом близнецов-партнеров обучают одному и тому же, но в разных возрастах. К данной группе относятся, например, упоминавшиеся работы А. Гезелла с сотрудниками. Во второй группе работ сравнивается эффективность разных способов обучения, в качестве примера может служить упомянутое шведское исследование обучения чтению. Авторы третьей, наиболее обширной группы работ изучают вопрос о том, насколько существенными могут быть различия, создаваемые средой у генетически идентичных индивидуумов. Примером могут служить экспериментальные методы изменения средовых воздействий, использованные в упоминавшихся исследованиях Московского Медико-генетического института.
Метод близнецовой пары. Еще в 30-х годах появились работы, в которых рассказывалось об особой психологической ситуации в парах близнецов, особом «эффекте пары», или «эффекте близнецовости». Они были выполнены и в психоаналитической традиции, и в психологии развития, и, специально, в русле психогенетических исследований. Основными характеристиками этой особой психологической ситуации являются две борющиеся тенденции: к идентификации со своим со-близнецом и, наоборот, к индивидуализации каждого члена пары. Первая приводит к ощущению себя сначала членом пары и лишь затем — отдельной личностью; вторая, в крайних случаях, может привести к выраженным конфликтным отношениям близнецов, своеобразному протесту против «второго — такого же». С возрастом первая тенденция обычно ослабевает, уступая место второй, причем у близнецов-мужчин данный процесс происходит легче, чем у женщин.
На этом фоне существуют более тонкие оттенки внутрипарных отношений. Например, еще в 1934 г. X. фон Браккен описал феномены гармонического и дисгармонического соперничества близнецов: «соревнование друг с другом» и «соревнование друг против друга». Сначала предполагалось, что первое характерно для МЗ близнецов, второе — для ДЗ, однако в дальнейшем это не подтвердилось; оба типа диадических взаимодействий встречаются и у тех, и у других. Он описал и некоторые специфические для пары социальные роли: «министра внешних сношений», осуществляющего общение с внешним миром, и «министра внутренних дел» («совесть пары»). Позже ролевые отношения, существующие в повседневной жизни близнецов, были описаны итальянским исследователем Л. Геддой и другими зарубежными авторами, а среди отечественных В.В. Семеновым [132, 137] и Н.В. Искольдским [65, 66]. Они же продемонстрировали, как распределение ролей влияет на оценку внутрипарного сходства и, соответственно, на величину коэффициента наследуемости, о чем уже го-
181
ворилось ранее. А в работах И.И. Канаева [69, 70] было показано, как роли «старшего» (родившегося первым) и «младшего», ведущего и ведомого, формируют различия в психологическом облике близнецов.
Однако наиболее детально близнецовую ситуацию исследовал Р. Заззо и описал в 1960 г. в книге «Близнецы: пара и личность» (на русский язык она, к сожалению, не переведена; краткий анализ см.: [152]). Он ввел и само понятие «близнецовая ситуация», показал ее проявления и обосновал необходимость ее изучения.
Сам Р. Заззо назвал исследование близнецовых пар «третьим методом» наряду с методами «контрастных групп» Гальтона (имеется в виду сопоставление групп МЗ и ДЗ близнецов) и «контрольного близнеца» Гезелла [459]. Согласно Р. Заззо, на фоне сходства близнецов, порождаемого и генетическим сходством и тем, которое формируется широкой средой, выделяется еще один фактор, создающий и специфическое сходство, и различия в глубине его, — близнецовая ситуация, своеобразный «микрокосм», результатом действия которого может стать и конвергенция, и дивергенция развития.
С помощью этого варианта метода решаются две задачи. Первая из них — общепсихологическая: поскольку индивидуум существует только внутри некоторых систем связей (например, в диаде) и «в этом смысле можно сказать, что мы все «близнецы»» [459, с. 25], постольку близнецовая пара, особенно генетически идентичные МЗ близнецы, может быть прекрасной моделью для изучения процессов индивидуализации, влияния конкретных средовых (внутрипарных и семейных) факторов на формирование тех или иных психологических черт и т.д. Вторая задача по существу есть проверка, во-первых, валидности самого близнецового метода, а также постулата о равенстве средовых влияний в парах МЗ и ДЗ близнецов и, во-вторых, вопроса о том, репрезентативна ли среда близнецов среде, в которой развиваются одиночнорожденные дети. Если условия развития и жизни близнецов высоко специфичны, имеют собственные, только им присущие закономерности, то близнецы перестают быть выборкой, репрезентативной общей популяции, и не могут быть использованы для изучения общих закономерностей.
К двум указанным задачам необходимо, на наш взгляд, добавить третью, связанную с жизнью самих близнецов. Чрезмерная, не компенсируемая идентификация себя со своим со-близнецом, или пары как целого, как особой «единицы», приводящая к отсутствию личной идентичности, может стать причиной формирования личностных черт, затрудняющих социальную адаптацию и в детстве, и во взрослом возрасте. Кроме того, отмеченные еще Р. Заззо и затем А. Р. Лурия особенности речевого развития близнецов, точнее формирование примитивной, синпрактической речи («криптофазия», по Заззо), может привести к отставанию и в общем интеллектуальном развитии [см.: 97]. Поэтому изучение специфических психологических характеристик
182
близнецовой пары необходимо и для проведения консультационной работы с родителями, имеющими детей-близнецов.
* * *
Методы, которыми располагает психогенетика, позволяют весьма надежно решать ее главную задачу: выяснение той роли, которую играют факторы наследственности и среды в формировании межиндивидуальной вариативности психологических и психофизиологических признаков, индивидуальных траекторий развития и т.д. Особенно ценен имеющийся в литературе анализ ограничений каждого метода, который позволяет либо контролировать помехи, либо компенсировать их объединением разных методов. Это — обязательное условие продуктивного развития любой точной науки.
Кроме того, методы психогенетики позволяют более надежно решать ряд негенетических задач, связанных с выделением актуальных для различных психологических черт средовых переменных, с более точной оценкой эффектов внешних воздействий и многих других.
Глава VIII
СТАТИСТИЧЕСКИЕ МЕТОДЫ ПСИХОГЕНЕТИКИ
Образно говоря, статистика является «правой рукой» психогенетики. Как уже отмечалось, психогенетика изучает вопросы наследования поведенческих признаков и психологических функций в популяциях, и по определению эта наука озабочена не отдельными индивидуальностями, а их разнообразием, т.е. популяционной изменчивостью (вариативностью, дисперсией) изучаемого признака. Иными словами, психогенетику интересуют вопросы, касающиеся характеристик распределений (среднего, дисперсии и других моментов распределения) индивидуальных значений по изучаемому признаку в популяции, а также вопросы о том, влиянием каких факторов — генетических или средовых — можно объяснить наблюдаемую изменчивость. Статистики, описывающие параметры популяции (выборки), приводятся в любом руководстве по статистике, поэтому здесь мы их касаться не будем, а перейдем сразу к статистическим решениям собственно психогенетических задач.
Как уже говорилось, психологические признаки принадлежат к классу количественных признаков, законы наследования которых существенно отличаются от менделевских. Особая здесь и статистика. Последовательно рассмотрим связанные с этим вопросы.
183
близнецовой пары необходимо и для проведения консультационной работы с родителями, имеющими детей-близнецов.
* * *
Методы, которыми располагает психогенетика, позволяют весьма надежно решать ее главную задачу: выяснение той роли, которую играют факторы наследственности и среды в формировании межиндивидуальной вариативности психологических и психофизиологических признаков, индивидуальных траекторий развития и т.д. Особенно ценен имеющийся в литературе анализ ограничений каждого метода, который позволяет либо контролировать помехи, либо компенсировать их объединением разных методов. Это — обязательное условие продуктивного развития любой точной науки.
Кроме того, методы психогенетики позволяют более надежно решать ряд негенетических задач, связанных с выделением актуальных для различных психологических черт средовых переменных, с более точной оценкой эффектов внешних воздействий и многих других.
Глава VIII
СТАТИСТИЧЕСКИЕ МЕТОДЫ ПСИХОГЕНЕТИКИ
Образно говоря, статистика является «правой рукой» психогенетики. Как уже отмечалось, психогенетика изучает вопросы наследования поведенческих признаков и психологических функций в популяциях, и по определению эта наука озабочена не отдельными индивидуальностями, а их разнообразием, т.е. популяционной изменчивостью (вариативностью, дисперсией) изучаемого признака. Иными словами, психогенетику интересуют вопросы, касающиеся характеристик распределений (среднего, дисперсии и других моментов распределения) индивидуальных значений по изучаемому признаку в популяции, а также вопросы о том, влиянием каких факторов — генетических или средовых — можно объяснить наблюдаемую изменчивость. Статистики, описывающие параметры популяции (выборки), приводятся в любом руководстве по статистике, поэтому здесь мы их касаться не будем, а перейдем сразу к статистическим решениям собственно психогенетических задач.
Как уже говорилось, психологические признаки принадлежат к классу количественных признаков, законы наследования которых существенно отличаются от менделевских. Особая здесь и статистика. Последовательно рассмотрим связанные с этим вопросы.
183
1. ГЕНЕТИКА КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ И ЕЕ ЗНАЧЕНИЕ ДЛЯ ПСИХОГЕНЕТИКИ
Генетика количественных признаков предоставляет психогенетике общую теорию, на базе которой строится методологический аппарат изучения природы индивидуальных психологических различий.
В самом общем виде генетика количественных признаков — применительно к психологическим задачам — исходит из того, что люди отличаются друг от друга по ряду сложных психологических признаков, и предлагает модель, в рамках которой межиндивидуальные различия по этим признакам могут быть описаны в терминах фенотипи-ческой дисперсии признака в популяции, а сама стенотипическая дисперсия может быть разложена на составляющие ее генетические и средовые компоненты.
МОДЕЛЬ ОДНОГО ГЕНА
Количественные генетические модели позволяют описать измеряемые эффекты различных генотипов, возможных в отдельно взятом локусе, и суммировать эффекты всех локусов, контролирующих тот или иной поведенческий признак. Причем количество локусов, контролирующих данный признак, обычно неизвестно, и чаще всего ученые делают допущение о том, что генетический контроль большинства поведенческих признаков осуществляется большим количеством генов, вклад которых в дисперсию изучаемого признака примерно одинаков. Одним из характерных признаков количественных генетических моделей является то, что они предполагают существование нормально распределенных фенотипических значений признаков, контролируемых множеством генов, эффекты которых, в свою очередь, опосредованы средовыми влияниями. Многолетние психологические исследования показали, что распределение большинства поведенческих признаков действительно соответствует нормальной кривой. Поэтому допущение о нормальности распределения признака, контролируемого большим количеством генов и значимых средовых влияний (т.е. являющегося мультифакторным), — психологически адекватная и статистически удобная модель для психогенетики. Важно заметить, что ожидаемая от полигенной системы нормальность распределения, будучи статистически удобной и эмпирически оправданной, не зависит от количества генов, контролирующих эту систему. Как статистические характеристики, так и теоретические положения, лежащие в основе моделей количественной генетики, одинаково правомерны для моделей, содержащих 1, 2, 25 или более генов. Именно поэтому мы начнем изложение основных признаков количественной генетики с рассмотрения модели одного гена и только потом перейдем к модели множественных генов (так называемой полигенной модели).
184
Аа аа__________________+------->___________АА
Рис. 8.1. Соотношение трех генотипов, возможных для двуаллельной системы.
Параметры d и h представляют собой разницы эффектов генов А-а. Аа может находиться на любой из сторон от т, что, соответственно, будет влиять на знак h. В примере, показанном па рисунке, значение h положительно [253, 342].
Генотипическое значение. Генотипическим значением называется некоторое количественное значение, приписываемое определенному генотипу. Так, в рамках простейшей двуаллельной системы (А и а) существуют два параметра, определяющие измеряемые эффекты трех возможных генотипов (АА, Аа и аа). Этими параметрами являются параметр d, представляющий собой удвоенную разницу между гомозиготами АА и аа, и параметр h, определяющий измеряемый эффект гетерозиготы Аа таким образом, что он не является точным усредненным эффектом двух гомозигот, Средняя точка между двумя гомозиготами, точка т, отражает среднее эффектов двух гомозиготных генотипов. Параметры d и h называются эффектами генотипов. Графически соотношение трех генотипов показано на рис. 8.1. Если в локусе отсутствует доминантность, то h будет равняться нулю, а значение генотипа Аа будет соответствовать значению в точке т. При полной доминантности значение Аа будет равняться значению АА. Если же доминантность А частичка, то Аа будет находиться ближе к точке АА (или аа, в зависимости от направления доминантности) и значение h будет положительным.
Приведем пример. Предположим, что известны гены, которые влияют на вес человека. Предположим также, что нормальный вес женщин среднего роста составляет 48-70 кг, т.е. разница между максимальным и минимальным значениями по весу равна 22 кг. Теперь предположим, что гены, контролирующие вариативность веса человека, расположены на каждой из 22 аутосомных хромосом (по одному на каждой), причем все гены вызывают примерно одинаковые эффекты. Тогда в рамках нашей гипотетической системы гомозиготы по каждому из изучаемых генов вкладывают примерно ± 1/2 кг (от средней точки), в зависимости оттого, являются они гомозиготами по аллелям, обозначаемым заглавной буквой (АА, ВВ, СС и т.д. - обладание этими генотипами повышает рост), или гомозиготами по аллелям, обозначаемым строчными буквами (аа, bb, ее и т.д. - обладание этими генотипами понижает рост). Рассмотренный пример, однако, невероятен по крайней мере по двум причинам: во-первых, генов, контролирующих вариативность веса человека, мы не знаем и, во-вторых, в реальной ситуации вклады генотипов, скорее всего, будут меньше или больше, чем 1/2 кг, затрудняя подсчет генотипического значения.
185
Модели генетики количественных признаков, во всяком случае в их классическом варианте, не являются ни средством идентификации конкретных генов, контролирующих вариативность признака, ни средством точного определения вклада каждого генотипа. Эти модели решают другую задачу, а именно задачу определения общего вклада генотипа в вариативность изучаемого признака в популяции.
Аддитивное генотипическое значение. «Аддитивное генотипическое значение» представляет собой фундаментальное понятие количественной генетики, поскольку оно отражает, насколько генотип «истинно наследуется». Аддитивный (суммарный) эффект генов представляет собой не что иное, как сумму эффектов отдельных аллелей. Более точно, аддитивное генотипическое значение есть генотипическое значение, обусловленное действием отдельных аллелей данного локуса. Генная доза генотипа подсчитывается на основе того, сколько аллелей определенного типа (например, аллелей А) присутствует в данном генотипе. Если наличие определенного аллеля в генотипе увеличивается на 1 (как это происходит, например, в случае перехода от генотипа аа к генотипу Aа), то аддитивное значение увеличивается на некоторую определенную величину. На рис. 8.2 дана графическая иллюстрация аддитивного генотипического значения при отсутствии доминантности. Эффект генотипа аа = -d, поэтому эффект аллеля (гена) а = 1/2(—d); эффект генотипа АА = d, поэтому эффект аллеля А = 1/2(d); соответственно, эффект генотипа Аа = 1/2(d) + 1/2(-d) = 0. Заметим, что аддитивные генные значения зависят от частоты встречаемости аллелей в популяции. При отсутствии доминирования аддитивный эффект полностью определяет генотипическое значение. Доминантность, однако, вносит самые разные отклонения от ожидаемых значений, — об этом пойдет речь ниже.
Теперь допустим, что каждый аллель генотипа имеет некоторый средний эффект. В этом смысле аддитивное генотипическое значение
представляет собой сумму средних эффектов каждого аллеля для всех аллелей, входящих в гено-' тип. Каждый аллель характеризу-
' ется определенным аддитивным
1 эффектом, соответственно, при
t унаследовании определенного ал-
\ леля от родителя ребенок насле-
j дует и аддитивный эффект этого
j_____ аллеля, т.е. вклад аллеля в генотип ребенка будет таким же, ка-
Рис. 8.2. При отсутствии доминант- ким был его (аллеля) вклад в ге-
ности (h = 0) аддитивное генотипи нотип родителя. И неважно,
ческое значение определяется генной сколько (много или мало) алле-
дозой. лей присутствует в данном локу-
186
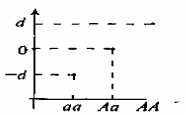
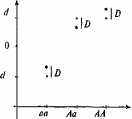
Рис. 8.3. Графическое изображение доминантных отклонений.
Наличие доминантности вносит разного рода изменения в аддитивное гепотипи-ческое значение.
Обозначения: • — аддитивные генотипические значения при условии полной доминантности; * — наблюдаемые аддитивные значения, определяемые на основе генных доз; D — доминантные отклонения, которые представляют собой разницу между ожидаемыми аддитивными значениями, соответствующими ситуации полной доминантности, и значениями, наблюдаемыми для определенных генных доз.
се или сколько локусов вовлечено в контроль вариативности по тому или другому признаку. Иными словами, аддитивное генотипическое значение представляет собой не что иное, как сумму вкладов каждого аллеля в генотип.
Доминантные отклонения. Доминантные отклонения есть мера того, насколько генотип отличается от своего ожидаемого аддитивного значения.
Доминантные отклонения (рис. 8.3) — это разница между ожидаемыми и наблюдаемыми значениями генотипов. Феномен доминантности допускает, что два аллеля одного локуса могут взаимодействовать друг с другом и тем самым менять генотипическое значение, которое наблюдалось бы в том случае, если бы они были независимы друг от друга и делали независимые вклады в генотипическое значение. Так, в результате взаимодействия аллелей Аа наблюдаемое генотипическое значение меньше того, которое ожидалось бы при условии полной доминантности. Напротив, значения АА и аа выше ожидаемых при допущении, что аллель А полностью доминантен по отношению к аллелю а.
Доминантность обязана своим возникновением уникальному сочетанию аллелей в данном локусе. Очевидно, что генотип потомка, наследующего только один аллель от каждого из родителей, в подавляющем большинстве случаев не может воспроизвести уникальность генотипа одного из них. Поэтому потомки будут отличаться от своих родителей в той мере, в какой аллели данного локуса не суммируются линейным образом при определении генотипического значения.
187
Рассмотрев типы генетических влияний, определим, как частоты встречаемости аллелей, определяющие эти типы, задают среднее значение генотипа в популяции. Допустим, что в популяции аллели А и а встречаются с частотами р и q, соответственно. Тогда первая колонка в табл. 8.1 показывает три возможных генотипа, вторая — частоты их встречаемости в популяции (при допущении, что особи в этой популяции образуют родительские пары случайным образом) и третья — значение генотипа. Популяционное среднее получается путем умножения значений генотипа на частоту встречаемости тех аллелей, которые этот генотип составляют, и последующего суммирования значений все трех генотипов.
Таблица 8.1