
1 Определение экосистемы. Свойства экосистемы. Структура экосистемы. Разнообразие экосистем биосферы Предметом
| Вид материала | Закон |
- Программа по «Экологии и природопользованию», 197.56kb.
- Б н. Ольга Викторовна Анисимова. Объем курса 36 часов. Форма отчет, 48.4kb.
- В. В. Снакин экосистемы концепция экосистемы, 239.27kb.
- Задачи на исследование изучить понятие экосистемы и исследовать их законы и закономерности, 22.52kb.
- М. Ю. Подходы к исследованию чувствительности модели экосистемы шельфа к вариациям, 593.73kb.
- Программа курса "Охрана окружающей среды", 611.72kb.
- " Организация экологического мониторинга и системы предотвращения загрязнения экосистемы, 143.97kb.
- Конспект лекций Москва 2006 г. Лекция № Основные источники техногенного загрязнения, 1559.24kb.
- Содержание: Введение, 770.55kb.
- Воздействие, 206.34kb.
12 Энергетическая структура экосистемы. Поступление и преобразование энергии в экосистеме.
Среди различных аспектов структуры экосистемы на первый план выступает энергетическая структура, т.к. поток энергии в значительной степени определяет характер потоков вещества и особой информации взаимодействии, а также пространственно динамической закономерности экосистем. Согласно закону максимилизации энергии (Г. и Ю. Одумы) выживает та экосистема, которая обеспечивает максимум, поступления и наиболее эффективное использование энергии. Поток энергии в экосистеме осуществляется через сложный механизм, включающий процессы ассимиляции, накопления, транспортиров и высвобождения энергии. Важнейший этап в том механизме - ассимиляция энергии представляет собой превращение кинетической энергии фотонов, в биологически доступную потенциальную энергию химических связей в органических соединениях. По способности к ассимиляции организмы делятся на два типа:
- автотрофы (продуценты) – организмы, самостоятельно синтезирующие органические вещества поглощая минимум вещества и энергию поступающую из вне.
- гетеротрофы (консументы, редуценты) организмы, получающие необходимые органические вещества, и энергию с пищей представляющую собой живую или мертвую биосферу.
К автотрофным относят зеленые растения и мех виды бактерий, гетеротрофы – относятся все животные, грибы, подавляющее большинство бактерий, водоросли, безхлорофильные растения. По своей функции в экологии гетеротрофы делятся на:
1) консументы (животные-фитофаги и зоофаги) ко второй группе относятся
2) редуценты деструкторы или микроконсументы, микроорганизмы (бактерии, грибы, разлагающее мертвое органическое вещество-детрит) до неорганических соединений.
Автотрофные организмы способные ассимилировать энергию двух источников: солнца и химическую энергию окислительно-востановительных реакций. Процесс ассимиляции солнечной энергии фотосинтез, а процесс ассимиляции химической энергии хемосинтез автотрофы для которых внешним источником энергии служит солнечный свет называются фотоавтотрофами или фотосинтетиками, автотрофы путем окисления получают энергию, разнообразных химических неорганических соединений называется хемоавтотрофами или хемосинтетиками
Транспорт энергии в экосистемах осуществляется через пищевые цепи. Пищевая цепь - это перенос энергии в форме пищи от ее источника (автотрофа) через ряд организмов, происходящих путем поедания одних органов другими
Пастбищная цепь, начинающаяся живыми растениями - продуцентами живой тканью, которого питаются растительные животные фитофаги.
Детритная цепь, начинающаяся мертвым органическим веществом, потребляемым мертвоедами, организмами – детритофагами. В сложных природных сообществах пищевые цепи тесно переплетаются друг, с другом образуя пищевые сети. Организмы, отстающие от начала пищевой цепи на одинаковое количество звеньев образуют трофический уровень. На первом трофическом уровне находятся только продуценты, ко 2 трофическому уровню относятся детритофаги, фитофаги (первичные консументы) на 3 м трофическом уровне находятся зоотрофы или вторичные консументы и т.д.
Границы между трофическими уровнями не четкие т.к. многие животные потребляют энергию более чем с одного трофического уровня, т.к. используя любую пищу, которая подходит им по размеру и другим признакам (до 80% общего потока энергии по пищевой цепи подменяется).
Согласно 2 закону термодинамики, процессы, связанные с превращением энергии, могут происходить самопроизвольно при условии, что часть этой энергии рассеивается в виде тепла. Зеленые растения ассимилируют от 1 до 5% получаемой солнечной энергии, в связи с этим в каждом звене пищевой цепи теряется значительная доля (80% - 95%) энергии заключенной в доступной пище. На каждый последующий трофический уровень поступает не более 80% синтезируемых углеводов, а растения получают от 1 до 5% солнечной энергии, то есть эффективность фотосинтеза составляет больше 1,5%. Фитофаги для образования своей биомассы, используют в среднем 1% энергии содержащейся в поедаемых ими растениях, на других трофических уровнях эффективность усвоения энергии доходит до 10%.
Из правила 10% вытекает следующие дополнения:
Чем ближе организм находится к началу пищевой цепи, тем большим количеством доступной энергии он располагает.
Пищевые цепи, включающие более 4-х 5 трофических уровней, встречается крайне редко.
Закономерности транспорта энергии в пищевых цепях удобно изображать графически с помощью диаграмм - экологических пирамид. Они представляют собой соотношения численности организмов их биомасс эквивалентным этим биомассам энергий или продуктивности организмов на различных трофических уровнях пищевой цепи.
Так как все экосистемы, включая биосферу, являются открытыми, то для своего функционирования, они должны получать и отдавать энергию, т.е. реальная функционирующая экосистема должна иметь вход и выход переработанной энергии. Энергия солнечного света поступает в экосистему, где фотоавтотрофными организмами превращается в химическую энергию, используемую для синтеза органических веществ из неорганических. В экосистеме поток энергии направлен в одну сторону: часть поступающей энергии солнца, преобразуется растениями и переходит на качественно более высокую ступень, превращаясь в органическое вещество, которое представляет собой более концентрированную форму энергии. Большая часть солнечной энергии приходит через экосистемы и покидает ее. В отличие от энергии, вода и элементы питания, необходимые для жизни, могут использоваться многократно (после отмирания органические вещества превращаются в неорганические). В состав экосистемы входит два компонента: сообщество живых организмом или биоценоз (биотический компонент) и физико-химическая среда или биотоп (абиотический компонент).
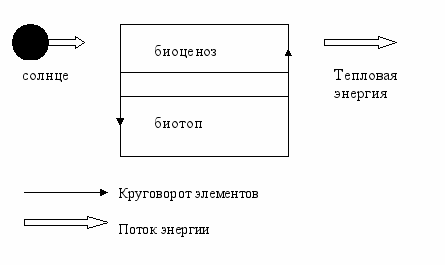
 Рис.1 - Функциональная схема экосистемы.
Рис.1 - Функциональная схема экосистемы.Таким образом, экосистему можно представить как единое целое, в котором биогенные вещества из абиотического компонента включаются в биотический и обратно, т.е. происходит постоянный круговорот веществ с участием биотического и абиотического компонентов.
Любая природная система может развиваться только за счет использования материально-энергетических и информационных возможностей окружающей ее среды (закон развития природной системы за счет окружающей среды).
13 Биологические ритмы. Сезонные и суточные биологические ритмы. Фотопериодизм
Биологические ритмы — это периодически повторяющиеся изменения интенсивности и характера биологических процессов и явлений. Они в той или иной форме присущи всем живым организмам и отмечаются на всех уровнях организации: от внутриклеточных процессов до биосферных. Биологические ритмы наследственно закреплены и являются следствием естественного отбора и адаптации организмов. Ритмы бывают внутрисуточные, суточные, сезонные, годичные, многолетние и многовековые.
Примерами биологических ритмов являются: ритмичность в делении клеток, синтезе ДНК и РНК, секреции гормонов, суточное движение листьев и лепестков в сторону Солнца, осенние листопады, сезонное одревеснение зимующих побегов, сезонные миграции птиц и млекопитающих и т.д. Биологические ритмы делят на экзогенные и эндогенные.
Экзогенные (внешние) ритмы возникают как реакция на периодические изменения среды (смену дня и ночи, сезонов, солнечной активности).
Эндогенные (внутренние) ритмы генерируются самим организмом. Ритмичность имеют процессы синтеза ДНК, РНК и белков, работа ферментов, деление клеток, биение сердца, дыхание и т.д. Внешние воздействия могут сдвигать фазы этих ритмов и менять их амплитуду. Среди эндогенных различают физиологические и экологические ритмы.
Физиологические ритмы (биение сердца, дыхание, работа желез внутренней секреции и др.) поддерживают непрерывную жизнедеятельность организмов.
Экологические ритмы (суточные, годичные, приливные, лунные и др.) возникли как приспособление живых существ к периодическим изменениям среды.
Физиологические ритмы существенно варьируют в зависимости от состояния организма, экологические — более стабильны и соответствуют внешним ритмам.
Экологические ритмы способны подстраиваться к изменениям цикличности внешних условий, но лишь в определенных пределах. Такая подстройка возможна благодаря тому, что в течение каждого периода имеются определенные интервалы времени (время потенциальной готовности), когда организм готов к восприятию сигнала извне, например яркого света или темноты. Если сигнал несколько запаздывает или приходит преждевременно, соответственно сдвигается фаза ритма. В экспериментальных условиях при постоянном освещении и температуре этот же механизм обеспечивает регулярный сдвиг фазы в течение каждого периода. Поэтому период ритма в этих условиях обычно не соответствует природному циклу и постепенно расходится по фазе с местным временем.
Эндогенный компонент ритма дает организму возможность ориентироваться во времени и заранее готовиться к предстоящим изменениям среды. Это так называемые биологические часы организма. Многим живым организмам свойственны циркадные и цирканные ритмы. Циркадные (околосуточные) ритмы — повторяющиеся изменения интенсивности и характера биологических процессов и явлений с периодом от 20 до 28 ч. Цирканные (окологодичные) ритмы — повторяющиеся изменения интенсивности и характера биологических процессов и явлений с периодом от 10 до 13 месяцев. Циркадные и цирканные ритмы регистрируются в экспериментальных условиях при постоянной температуре, освещенности и т.д.
Ритмический характер имеют физическое и психологическое состояния человека. Нарушение установившихся ритмов жизнедеятельности может снижать работоспособность, оказывать неблагоприятное воздействие на здоровье человека. Изучение биоритмов имеет большое значение при организации труда и отдыха человека, особенно в экстремальных условиях (в полярных условиях, в космосе, при быстром перемещении в др. часовые пояса и т.д.).
Несовпадение во времени между природными и антропогенными явлениями часто приводит к разрушению природных систем. Например, при проведении слишком частых рубок леса.
Современная систематика живых организмов строится на основе степени родства организмов. В основу экологических классификаций могут быть положены самые разнообразные критерии: способы питания, передвижения, отношения к температуре, влажности, свободному кислороду и т.п. Разнообразие адаптации к среде создает необходимость множественных классификаций.
На растения влияют не только интенсивность и яркость, но и продолжительное освещение с которым связан суточный и сезонный ритмы развития растений. Этот ритм называют фотопериодизмом. Ритмические изменения морфологических, биохимических и физических свойств и функций организмов под влиянием чередования и длительности освещения получили название фотопериодизма.
По реакции растений на длину светового дня выделяют растения длинного дня (фотопериод более 16-20 часов). К ним относится картофель, пшеница, шпинат. Растения короткого дня (фотопериод менее 8-12 часов) и нейтральные к длине обычного дня (горчица, одуванчик, томат). Освещенность и фотопериодизм имеет определенное значение для развития многих животных. Личиночные стадии многих беспозвоночных тормозятся интенсивным светом из-за чувствительности к ультрофиалетовым лучам. С освещенностью, связанны ночной и дневной образы жизни. Периодичность освещения, изменение фотопериода опережают изменение температуры и служат сигналом для запуска периодичных сезонных изменений жизнедеятельности, таких как цветение, размножение, линька, спячка, метаморфозы у насекомых
Каждое местообитание характеризуется определенным световым режимом, соотношением интенсивности (силы), количества и качества света.
14 Экологическая ниша. Фундаментальная и реализованная экологические ниши. Принцип Гаузе
Любая популяция (вид) занимает определенное местообитание и определенную экологическую нишу.
Местообитание — это территория или акватория, занимаемая популяцией (видом), с комплексом присущих ей экологических факторов. Местообитание вида является компонентом его экологической ниши. Применительно к наземным животным местообитание вида называется стация, местообитание сообщества — биотоп.
Экологическая ниша — совокупность всех факторов среды, в пределах которых возможно существование вида в природе. То есть экологическая ниша — это место вида в природе, включающее не только его положение в пространстве и отношение к абиотическим факторам, но и его функциональную роль в сообществе (прежде всего трофический статус). Местообитание — это как бы «адрес» организма, а экологическая ниша — это его «профессия».
Для характеристики экологической ниши обычно используют два важных показателя: ширина ниши и степень перекрывания ее с соседними. Экологические ниши разных видов могут быть разной ширины и перекрываться в различной степени.
Разделение экологических ниш между видами происходит за счет приуроченности разных видов к разным местообитаниям, разной пищи и разному времени использования одного и того же местообитания.
Принцип конкурентного исключения (принцип Гаузе) гласит: «Два вида не могут сосуществовать в одной и той же местности, если их экологические потребности идентичны. Такие виды обязательно должны быть разобщены в пространстве или во времени».
Группы видов в сообществе, обладающие сходными функциями и нишами одинакового размера, т.е. роль которых в сообществе одинакова или сравнима, называются гильдиями. Например, лианы тропического леса представлены многими видами растений. Между видами внутри гильдии наблюдается особенно острая конкуренция.
Виды, занимающие одинаковые ниши в разных географических областях, называются экологическими эквивалентами. Например, крупные кенгуру Австралии, бизоны Северной Америки, зебры и антилопы Африки и т.д. являются экологическими эквивалентами. В настоящее время они значительно замещены коровами и овцами.
Выделяют 4 типа расположения экологических ниш 2-х видов одного биотопа:
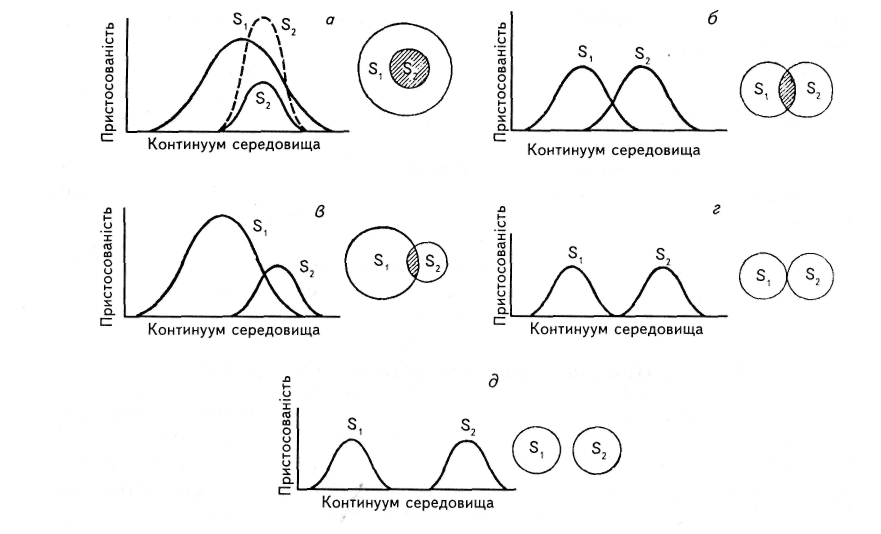
Рис. 1 Випадки можливих взаємних ніш, проілюстровані за допомогою поняття щільності пристосованості (зліва) та моделей теорії множин (справа):
а — ніша всередині ніші. Ніші виду 2 (S2) розташовані всередині ніші (St). Можливі два наслідки конкуренції: 1) якщо вид 2 має перевагу (переривчаста лінія), то він буде співіснувати при неповному використанні спільних ресурсів з видом і; 2) якщо переваги має вид 1 (суцільні лінії), то він буде використовувати весь градієнт ресурсів, а вид 2 буде витіснений;
6 — перекривання ніш однакової ширини. Конкуренція однакова в обох напрямках;
в — перекривання ніш неоднакової ширини. Конкуренція неоднакова у двох напрямах, оскільки частина простору ніш, яка входить до області перекривання, у виду 2 більша, ніж у виду 1;
г — прилягання ніш. Пряма конкуренція неможлива, але подібне може бути наслідком її уникання;
д — повний розділ ніш. Конкуренція неможлива і її важко навіть передбачити. Чому ж тут не спостерігається кормова конкуренція? А тому, що кожний із видів в процесі еволюції пристосувався до своєї екологічної ніші.
Екологічна ніша — фізичний простір з властивими йому екологічними умовами, що визначають існування будь-якого організму, місце виду в природі, що включає не лише становище його в просторі, а й функціональну роль у біоценозі та ставлення до абіотичних факторів середовища існування. Екологічна ніша характеризує ступінь біологічної спеціалізації даного виду.
Различают фундаментальную (потенциальную) нишу, которую организм мог бы занимать в отсутствие конкурентов, хищников и других врагов и в которой физические условия оптимальны; и реализованную – фактический диапазон условий существования организма, который или меньше фундаментальной ниши или равен ей.
Фундаментальную нишу называют иногда предконкурентной, а реализованную – постконкурентной.
Два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое то время обязательно будет вытеснен. Выделяют четыре направления конкуренции влияющие на:
- географическое расширение видов (интродукция)
- на размещение вида в различных биотопах одной и той же местности.
- влияние на морфологию и продуктивность.
- влияние на эволюцию видов биоценоз конкурентные отношения
15 Экологические стратегии популяций
Экологическая стратегия выживания — комплекс свойств популяции, направленных на повышение вероятности выживания и оставление потомства, называется. Это общая характеристика роста и размножения. Сюда входят темпы роста особей, время достижения половозрелости, плодовитость, периодичность размножения и т.д.
Так А.Г. Раменский (1938) различал три основные типа стратегий выживания среди растений: виоленты, патиенты и эксплеренты.
Виоленты (силовики) — подавляют всех конкурентов, например, деревья, образующие коренные леса.
Патиенты — виды, способные выжить в неблагоприятных условиях («тенелюбивые», «солелюбивые» и т.п.).
Эксплеренты (наполняющие) — виды, способные быстро появляться там, где нарушены коренные сообщества, — на вырубках и гарях (осины), на отмелях и т.д.
Экологические стратегии популяций отличаются большим разнообразием. Но при этом все их многообразие заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r-стратегия и К-стратегия.
г-стратеги (r-виды, г-популяции) — популяции из быстро размножающихся, но менее конкурентоспособных особей. Имеют j-образную кривую роста численности, не зависящую от плотности популяции. Такие популяции быстро расселяются, но они малоустойчивы. К ним относятся бактерии, тли, однолетние растения и др. (табл. 6).
К-стратеги (К-виды, К-популяции) — популяции из медленно размножающихся, но более конкурентоспособных особей. Имеют S-образную кривую роста численности, зависящую от плотности популяции. Такие популяции населяют стабильные местообитания. К ним относятся человек, кондор, деревья и др. (табл. 6).
Приспособление особей в популяции в конечном счете направлены на повышение выживания и оставления потомства. Экологическая стратегия популяций это общая характеристика ее роста и размножения сюда входят темпы роста особей, время достижения половой зрелости, плодовитости, периодичность размножения вида и занимающие нарушения места обитания и имеющие большое значение ч-видами или виды оппортуниста. Виды с относительно низким значением ч и для в которых скорость размножения чувствительность к плотности популяции называется К-видами.
Характерные особенности
| r | К |
| Различают быстро (высокая плодовитость, время генерации короткое) | Размножение медленно низкая плодовитость продолжительность времени генерации |
| V размен не зависит от плотности популяции | V размен зависит от плотности популяций если р падает V быстро увеличивается |
| Энергия и вещество распределены между многими потомками | Энергия и вещество концентрируются в немногих потомках, родители заботятся о потомстве |
| Размеры популяций некоторое время могут превышать поддерживающую емкость среды | Размеры популяций близки к равномерному уровню |
| Вид не всегда устойчив на данной территории | Вид устойчив на данной территории |
| Расселяются широко и в больших количествах | Расселяются медленно |
| Размножение идет с большой затратой энергии и вещества | Размножение идет с относительно малыми затратами, большая часть энергии расходуется на вегетативный рост |
| Малые размеры особей | Крупные размеры, у деревянных стеблей и большие корни |
| Малая продолжительность жизни, могут рассеяться на открытых местах, место обитания сохраняется не долго | Большая продолжительность жизни плохо приспособлены к росту на открытых местах , место обитания устойчивое и сохраняется долго |
| Слабые конкуренты, слабо развиты защитные приспособления, лучше приспособлены к изменению окружающей среды (бактерии, тля, мучные хрущаки, однолетние растения) | Сильные конкуренты хорошие защитные механизмы, менее устойчивы к изменениям условий среды (крупные тропические бабочки, кондор, альбатрос, человек, деревья) |
Виды с r- стратегий быстрее заменяют нарушенное местообитания (горная порода, лесные вырубки, выгоревшие участки леса) Они занимают данное место обитания в течение жизни одного или нескольких поколений, в дальнейшем они переселяются на новое место. Виды с К-стратегией более конкурентно способные и обычно вытесняют К-виды. Могут использовать одно и то же место обитания по разному, то они могут сосуществовать вместе.
Следует отметить, что одну и ту же среду обитания разные популяции могут использовать по-разному, поэтому в одном и том же местообитании могут сосуществовать виды с r- и К-стратегиями. Между этими крайними стратегиями существуют переходы. Ни один из видов не подвержен только r- или только К- отбору Гомеостаз популяции — поддержание определенной численности (плотности). Изменение численности зависит от целого ряда факторов среды — абиотических, биотических и антропогенных. Однако всегда можно выделить ключевой фактор, наиболее сильно влияющий на рождаемость, смертность, миграцию особей и т.д.