
Высшее профессиональное образование т. Я. Дубнищева концепции современного естествознания
| Вид материала | Документы |
СодержаниеИонный перенос 12.4. Процессы фотосинтеза и клеточного дыхания |
- Учебно-методический комплекс концепции современного естествознания высшее профессиональное, 2306.3kb.
- Учебно-методический комплекс концепции современного естествознания высшее профессиональное, 2307.28kb.
- С. Г. Хорошавина концепции современного естествознания курс лекций, 6750.33kb.
- С. Г. Хорошавина концепции современного естествознания курс лекций, 5892.74kb.
- В. М. Найдыш Концепции современного естествознания, 8133.34kb.
- Учебно-методический комплекс дисциплины концепции современного естествознания Специальность, 187.08kb.
- Концепции Современного Естествознания, 274.86kb.
- Программа курса «Концепции современного естествознания», 168.05kb.
- Программа дисциплины Концепции современного естествознания Специальность/направление, 456.85kb.
- Бюллетень новых поступлений в нб согу за период с 05. 2011 по 10. 2011гг, 975.89kb.
Клеткам присуще мембранное строение — это одно из положений клеточной теории. Среди мембранных органоидов — наружная цитоплазматическая мембрана (НЦМ), эндоплазматиче-ская сеть (ЭПС), аппарат Гольджи (АГ), лизосомы (Л), митохондрии (М), пластиды (П). В основе всех этих органелл лежит биологическая мембрана, все они имеют единый план строения. Мембранные структуры — арена важнейших жизненных процессов.
Биологическая мембрана (клеточная или плазматическая) — пленка, покрывающая клетку, и настолько тонкая, что ее удалось обнаружить лишь с помощью трансмиссионного электронного микроскопа. Все мембраны построены по одному плану, всегда слоистые. Поперечный разрез показывает, что по обе стороны внутренней, более светлой линии расположены более темные. Мембраны были открыты более века назад, но их роль в механизмах жизнедеятельности клеток до недавнего времени сводили в основном к барьерной функции. Опыты показали, что малые молекулы быстрее усваиваются живой клеткой, чем большие, и вещества, растворимые в воде и нерастворимые в жирах, проникают в клетку медленнее, чем растворимые в жирах. Значит, мембраны содержат жироподобные вещества — липиды и белки, способные связывать воду.
Липиды в мембранах содержат фосфорную кислоту, потому их называют фостатидами. Пример — лецитин. Капля такого ли-
477

 пида мгновенно растекается по водной поверхности, и пленка образует мономолекулярный слой. Они обладают водоотталкивающими, или гидрофобными (от греч. phobos — страх, боязнь), свойствами. Фосфорная кислота растворима в воде, или гидрофильна (от греч. philia — любовь). Она как бы притягивает воду, а водоотталкивающие остатки жирных кислот, расположенные на другом конце молекулы, как бы избегают ее. Так как гидрофобные концы липидных молекул не могут сближаться ни с клеточной оболочкой, ни с протопластом, они обращены друг к другу «головами». Так образуется бимолекулярный слой, у которого наружу выставлены гидрофильные части. Дополнительные гидрофильные белки в мембране повышают устойчивость описанного выше липидного бимолекулярного слоя.
пида мгновенно растекается по водной поверхности, и пленка образует мономолекулярный слой. Они обладают водоотталкивающими, или гидрофобными (от греч. phobos — страх, боязнь), свойствами. Фосфорная кислота растворима в воде, или гидрофильна (от греч. philia — любовь). Она как бы притягивает воду, а водоотталкивающие остатки жирных кислот, расположенные на другом конце молекулы, как бы избегают ее. Так как гидрофобные концы липидных молекул не могут сближаться ни с клеточной оболочкой, ни с протопластом, они обращены друг к другу «головами». Так образуется бимолекулярный слой, у которого наружу выставлены гидрофильные части. Дополнительные гидрофильные белки в мембране повышают устойчивость описанного выше липидного бимолекулярного слоя.Структуру мембран — внутри «масло» (двойная липидная пленка), снаружи «хлеб» (белковая оболочка) — называют сэндвич-структурой. Такую структуру химики могут получать искусственно.
Мембрана — двухмерно ориентированный раствор разных белковых молекул и белковых кластеров из нескольких сотен молекул в вязком слое. Белковые молекулы в большинстве своем свернуты в клубки (глобулы) и асимметричны. Их выступающие из мембраны части обладают электрическими зарядами, причем на внешней поверхности суммарный заряд оказывается отрицательным. Молекула фосфолипидов сильно асимметрична. Одна ее часть несет электрические заряды, образуя «полярную» головку, другая — электронейтральный углеводородный «хвост». В водной среде полярные головки выступают из воды, а углеводородные хвосты, из-за гидрофильных и гидрофобных взаимодействий погружаются. Так как все белки участвуют в диффузном движении, их распределение по мембране в каждый момент случайно. Коэффициент диффузии белковых молекул по мембране порядка 5 • 10-14 м2/с, а для гемоглобина в водном растворе — на три порядка больше. Значит, вязкость мембранной фазы на три порядка больше, чем у воды. Некоторые белки способны только к поступательному перемещению, другие могут вращаться в плоскости мембраны, есть и такие, которые перемещаются с одной стороны мембраны на другую. Последние участвуют в транспортировании веществ через мембрану.
Ионный перенос — проявление мембранной возбудимости. Через мембрану осуществляется обмен с внешней средой — питание и выделение отходов. Несмотря на хаотические движения, молекулы стремятся переместиться в сторону меньшего давления (перемещение по градиенту давления, или концентрации, называют диффузией). Мембрана обеспечивает стабильность химического содержимого клетки и, обладая избирательной способностью, регулирует обмен с окружающей средой. Вещества, растворимые в липидах, проходят через мембрану, не растворяясь в ней. Пере-
478
мещение ионов и органических мономеров типа аминокислот и глюкозы происходит много быстрее, чем этого можно было бы ожидать от полярных молекул. Имеет место и перемещение против градиента концентрации — так называемый активный транспорт, требующий затрат энергии. Наиболее изу
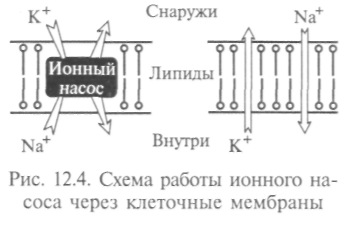 чен такой активный транспорт: процесс откачки ионов натрия из клетки и накачки в нее ионов калия, в котором «Na—К — насос» использует энергию АТФ (рис. 12.4). Этим путем откачиваются ионы натрия из клетки и накачиваются ионы калия против градиента концентрации.
чен такой активный транспорт: процесс откачки ионов натрия из клетки и накачки в нее ионов калия, в котором «Na—К — насос» использует энергию АТФ (рис. 12.4). Этим путем откачиваются ионы натрия из клетки и накачиваются ионы калия против градиента концентрации.Специальные белковые молекулы в мембране переносят различные вещества. Так, с их помощью клетки печени, эритроциты и мышечные клетки быстро поглощают глюкозу. В настоящее время выяснены пять молекулярных форм переносчика глюкозы, причем каждая из них приспособлена к нуждам той ткани, в которой она содержится. Посредством такой облегченной диффузии вещества могут выводиться из клеток. Через мембрану осуществляется и пассивный транспорт. Таков осмос — прохождение воды через полупроницаемую мембрану. Могут через нее путем диффузии проникать вещества, растворимые в липидах (жирные кислоты и эфи-ры), и некоторые ионы.
Клеточная мембрана, помимо барьерной функции, обеспечивает обмен между цитоплазмой и внешней средой, из которой в клетку поступают вода, ионы, различные молекулы, а выводятся продукты обмена веществ и синтезированные в клетке вещества. Транспортные функции не ограничиваются маленькими молекулами. Благодаря фагоцитозу, открытому и описанному И. И. Мечниковым (1882), в клетку могут проникать и крупные молекулы биополимеров. Твердая частица, оказавшаяся вблизи клетки, окружается выростами мембраны и затягивается внутрь. Процесс фагоцитоза свойствен простейшим, лейкоцитам, клеткам капилляров костного мозга, печени, надпочечников, селезенки. Существует и еще один вид активного транспорта — пиноцитоз. Таким путем происходит поглощение клеткой жидкости в виде мелких капель с растворенными в них высокомолекулярными веществами. Капли захватываются выростами мембраны, погружаются в цитоплазму и усваиваются. Это явление свойственно животным клеткам.
Регулируя обмен между клеткой и окружением, мембраны обладают рецепторами, воспринимающими внешние сигналы (свет, движение бактерий к источнику пищи, ответы на гормоны). Безусловно, важно, что на них происходит превращение энер-
479
 гии. Так, на внутренних мембранах хлоропластов происходит фотосинтез, а на внутренних мембранах митохондрий — окислительное фосфорилирование. Компоненты мембран движутся и перестраиваются, поскольку созданы из белков и липидов, что определяет одно из важнейших свойств живого — раздражимость.
гии. Так, на внутренних мембранах хлоропластов происходит фотосинтез, а на внутренних мембранах митохондрий — окислительное фосфорилирование. Компоненты мембран движутся и перестраиваются, поскольку созданы из белков и липидов, что определяет одно из важнейших свойств живого — раздражимость.Роль мембран стала вырисовываться иначе после того, как английские ученые химик Д. Кроуфут-Ходжкин и физиолог А. Хаксли сформулировали теорию проведения нервного импульса (1952), а Е.Сюзерленд открыл (1972) существование на возбудимой мембране переносчика информации внутрь клетки (молекулы цАМФ — циклического аденозинмонофосфата). Этот вопрос еще недостаточно изучен и является предметом особого интереса, так как через познание механизма функционирования возбудимых мембран лежит путь к диагностике и лечению многих болезней.
Возбудимость — реакция клетки на воздействие, происходящая с многократным усилением по энергии. Возбудимость — общее свойство клеток, не только сердечных, мышечных или нервных. Состояние мембраны отражает состояние клетки в целом. Возбудимыми называют мембраны, окружающие клетку и способные менять свою проницаемость для ионов при различных химических и физических воздействиях. Функционирование таких мембран обеспечивает не только внутриклеточную регуляцию, но и управление и синхронизацию работы соседних клеток и даже органов с помощью химических и электрических каналов связи, которые составляют основу гормональной и нервной регуляции. Основные компоненты этих систем находятся в мембранах.
12.4. Процессы фотосинтеза и клеточного дыхания
От фотосинтеза, который делает энергию и углерод доступными для живых организмов и обеспечивает выделение кислорода в атмосферу, зависит все живое на Земле. Менее 1 % солнечной энергии, падающей на Землю, поглощается растениями. Они связывают углекислый газ атмосферы (и воду) в количестве около 150 1012 кг сухого органического топлива в год, или порядка 1 кг сухого вещества с 1 м2 за год. Часть этого органического вещества поглощается травоядными животными, которыми, в свою очередь, питаются другие животные и человек. Растительные и животные остатки разлагаются бактериями и грибами до уровня исходных неорганических веществ. Затем этот круговорот замыкается: энергия солнечного излучения, поглощенная растениями, переходит в теплоту и излучается Землей в космическое пространство. И жизнь на Земле есть процесс поглощения солнечного света. Человечество зависит от фотосинтеза и потому, что оно использует ископаемое энергетическое топливо, образовавшееся за
480
миллионы лет. Годовая фиксация углерода в процессе фотосинтеза оценивается в 75 • 1012 кг. Из общего количества солнечной радиации, попадающей на Землю, до поверхности доходит примерно 50 %, а из нее только 25 % лучей имеют длины волн, подходящие для фотосинтеза, 1 % энергии доходит до растений, а 0,4 % используется ими для увеличения своей биомассы (рис. 12.5).
Автотрофные («самопитающиеся») организмы осуществляют фотосинтез, не питаясь другими организмами. Такие есть среди бактерий, источник энергии для них — химические реакции. Но запасы химической энергии на Земле ничтожны по сравнению с энергией, поступающей от Солнца.
В конце XVIII в. считали, что растения получают питательные вещества из воды, находящейся в почве. Голландский естествоиспытатель Ян ван Гельмонт первым проделал опыт по изучению процесса питания растений. Он посадил дерево ивы массой 2,3 кг в кадку с землей массой 90,8 кг, поливал его и лишь через пять лет обнаружил, что масса почвы почти не изменилась, а дерево выросло и стало весить 76,9 кг. Английский химик Дж. Пристли, открывший кислород (1774) и получивший хлористый водород и аммиак, обнаружил, что растения и животные по-разному меняют состав окружающего их воздуха. Помещая в закрытый сосуд горящую свечу и мышь, он отметил, что свеча гасла, а мышь из-
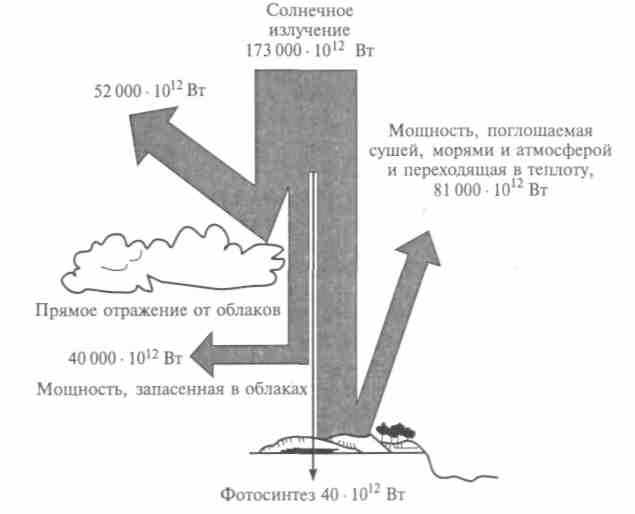
Рис. 12.5. Распределение мощности солнечного излучения, падающего
на Землю


дыхала. Но, когда он помещал в сосуд живое растение, свеча после этого какое-то время горела, хотя сосуд был по-прежнему закрыт. Так Пристли установил, что «в растениях присутствует что-то способное исправлять воздух, испорченный горением свечи». В 1782 г. Ж. Сенебье сумел показать, что растения поглощают С02 и одновременно выделяют 02.
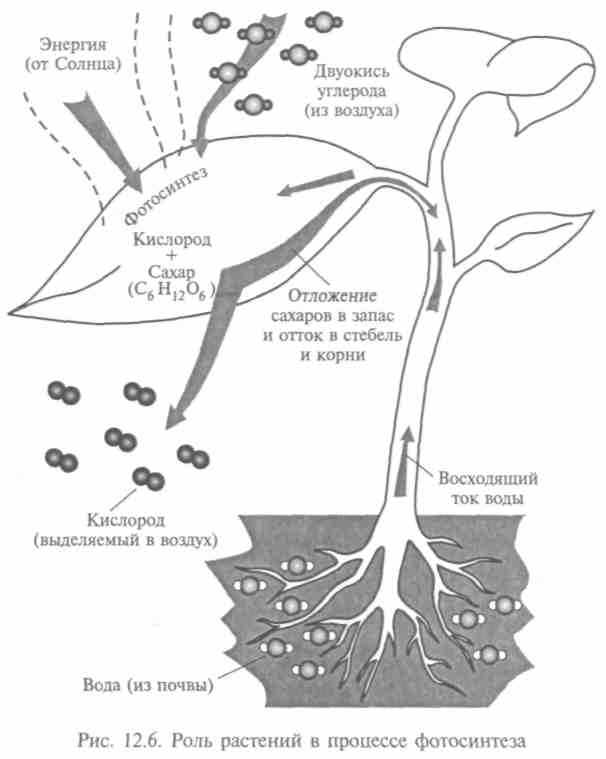
К началу XIX в. было выяснено, что растения могут выделять кислород только на свету, отсюда и название — фотосинтез (рис. 12.6). Австрийский врач Я.Ингенхауз написал первое уравнение процесса, не зная еще, какие растительные ткани образуются. При использовании микроскопа нашли, что крахмальные зерна при фотосинтезе растут. Отсюда предположение, что при фотосинтезе возникают углеводы, имеющие своим источником С02.
Исходные соединения для фотосинтеза — неорганические вещества: вода и двуокись углерода. Они энергетически бедны, но
482
из них строятся более сложные богатые энергией питательные вещества. В качестве побочного продукта фотосинтеза вьщеляется молекулярный кислород. Процесс фотосинтеза обычно представляют уравнением: 6С02 + 6Н20 = С6Н,206 + 602.
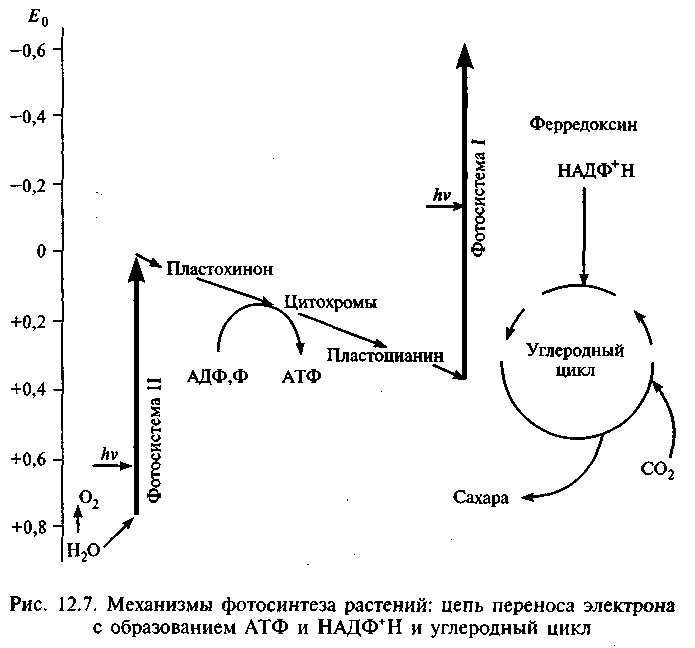
Реакция идет за счет энергии света и хлорофилла, и получаются молекулы кислорода и сахара (рис. 12.7). В 1941 г. с помощью масс-спектрометра методом изотопов было установлено, что источником кислорода служит вода. (Обычно кислород имеет массовое число 16, но есть и стабильный изотоп с массовым числом 18.) Оказалось, что на первой стадии фотосинтеза водород получается путем расщепления воды. Энергию для этого растения берут у света (реакция фотолиза), выделяя кислород как ненужный побочный продукт. Во второй стадии водород соединяется с двуокисью углерода и образуется углевод. Присоединение водорода — один из примеров реакции восстановления. Для первой стадии характерны световые реакции, для второй свет не нужен; хотя они тоже происходят на свету (рис. 12.8), их называют тем-новыми.
483

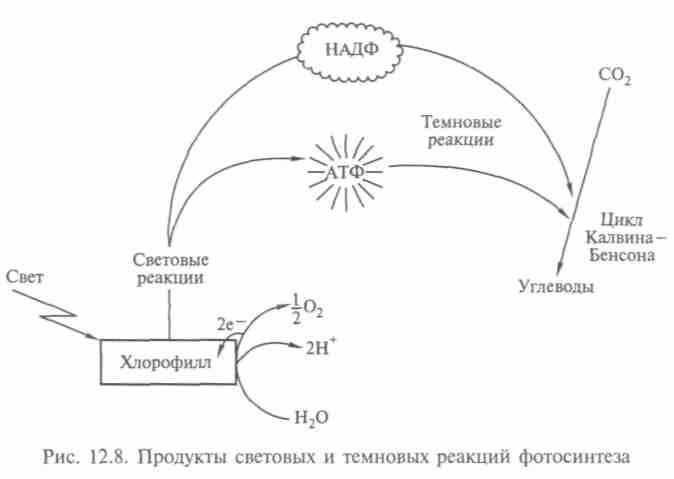
В конце 50-х гг. выяснили, что каждая из стадий реакций фотосинтеза происходит в разных частях листа: первая — в мембранах хлоропластов, вторая — в их строме. Арион показал (1958), что первая стадия во многом аналогична дыханию, при котором происходят фосфорилирование АДФ с затратами энергии, перенос электронов в мембранах и преобразование световой энергии в химическую. Дыхание у растений — процесс окисления углеводов с освобождением энергии, необходимой для жизнедеятельности, он происходит в митохондриях. У аэробных организмов поглощается 02 и выделяется С02. Выделенная энергия идет на синтез АТФ. Поэтому процессы дыхания у растений и фотосинтез — две стороны обмена веществ: диссимиляции и ассимиляции.
Немецкий ученый Т. Энгельман в конце XIX в. показал, что зеленый пигмент хлоропластов — хлорофилл — важен в процессе фотосинтеза. Экспериментируя с водорослью спирогирой, он определил области спектра света, в которых выделение кислорода идет наиболее эффективно. Ими оказались красные и синие области, поглощаемые именно хлорофиллом, обеспечивая ему зеленый цвет. Другие пигменты хлоропластов (они имеют желтую или бурую окраску, наблюдаемую у листьев осенью, когда хлорофилл разрушается и уже не маскирует цвет листа) играют вспомогательную роль в этом процессе, перенося энергию. Английский ученый Дж. Стоукс установил, что зеленый растительный пигмент состоит из смеси различных веществ (1864). Русский физиолог и биохимик М. С. Цвет исследовал их с помощью изобретенного им метода хроматографии. Немецкий химик и биохимик
484
Р. Вильштеттер разработал методы извлечения растительных экстрактов без повреждения молекул и детально исследовал хлорофилл, обратив внимание на аналогию с гемоглобином крови. Его работы по изучению структуры хлорофилла продолжил химик-органик X. Фишер.
Синтезировать молекулу хлорофилла удалось только в 1960 г. А. Калояну, Г. Колеру и Р. Вудворду. Все эти исследования с хлорофиллом важны для консервации зеленых кормов, в которых потери питательных веществ обычно составляют более 50 %. При определенной дозировке серной и соляной кислот, прекращающих процессы окисления в растительной массе, удалось обеспечить почти полное сохранение витаминов и белков. Выдающуюся роль в решении этих проблем сыграл финский биохимик А. Виртанен, создавший метод консервирования зеленых кормов.
Фотосинтетические пигменты и молекулы, встроенные в мембраны тиколаида хлоропласта (см. рис. 12.3), образуют цепь переноса электронов. Мембраны окружены стромой — основным веществом хлоропласта, которое содержит хлоропластную ДНК, рибосомы и ферменты, участвующие в фиксации углерода. Снаружи стромы окружены двойной мембраной, отделяющей хлоропласт от цитоплазмы клетки. Пигмент хлорофилл уникален: при поглощении солнечной энергии один из электронов отрывается от своей молекулы, а затем передается по электронтранс-портной цепи от одного переносчика к другому. Эти электроны замещаются путем разложения воды и разделения ее водородных атомов на ионы водорода и электроны. При этом для получения одной молекулы кислорода должны разложиться две молекулы воды, и останется четыре протона внутри тиколаида. Почти весь кислород в земной атмосфере возник именно таким образом.
Электроны, пройдя по электронтранспортной цепи, присоединяются к молекуле-переносчику, которой служит НДЦФ+, переходящий в восстановленную форму НАДФ+Н. Этот процесс происходит на наружной поверхности мембран. Ионы водорода, пройдя по каналам в мембране на наружную сторону, приобретают энергию за счет электрохимического потенциала для синтеза АТФ. Последние используются в серии реакций, которые «фиксируют» С02 в форме углеводов. Сначала С02 присоединяется к органической молекуле — пятиуглеродному сахару, переводя ее в нестабильную шестиуглеродную форму. Она очень быстро распадается на две трехуглеродные молекулы, которые принимают по фосфатной группе от АТФ; эти группы присоединяются высокоэнергичной связью, обогащая энергией всю молекулу. После передачи энергии молекуле связи рвутся, и каждая молекула присоединяет по одному атому водорода от НАДФ+Н. В то же время происходит перенос четырех протонов с наружной стороны мембраны во внутреннюю. Одновременно протекает несколько таких однотипных реакций, и они идут по циклам.
Знаменитые опыты с хлореллой в 1946 г. провел американский биохимик М. Калвин. Он помещал зеленые водоросли в специальный сосуд и освещал их, одновременно пропуская С02 через воду,
485


 меченую изотопом С-14. При освещении изотоп включался в цепь фотосинтеза. При изменении времени облучения (от 1 до 30 с) последовательно прослеживались разные этапы фотосинтеза. Калвин показал, что двуокись углерода фиксируется в форме фос-фоглицериновой кислоты. Это было открытие — углерод просто подключался к одному из известных звеньев цепи углеводного обмена. Так было доказано, что углерод входит в состав глюкозы и других сложных Сахаров. В последующем Калвин продолжал работы по применению открывшейся ему тайны фотосинтеза в повышении урожайности и развитию «зеленой энергетики».
меченую изотопом С-14. При освещении изотоп включался в цепь фотосинтеза. При изменении времени облучения (от 1 до 30 с) последовательно прослеживались разные этапы фотосинтеза. Калвин показал, что двуокись углерода фиксируется в форме фос-фоглицериновой кислоты. Это было открытие — углерод просто подключался к одному из известных звеньев цепи углеводного обмена. Так было доказано, что углерод входит в состав глюкозы и других сложных Сахаров. В последующем Калвин продолжал работы по применению открывшейся ему тайны фотосинтеза в повышении урожайности и развитию «зеленой энергетики».Водоросли, составляющие огромную группу растений, являются фо-тосинтезирующими организмами, выделяющими кислород. Они эволюционировали в водной среде и освоили ее. Считают, что из синезеленых водорослей произошли все хлоропласты растений. На долю океана приходится 50 % мировой первичной продукции в виде фиксированного углерода, и ее образуют водоросли, хотя фотосинтез происходит только в поверхностных слоях, куда проникает солнечный свет и где лимитирующим фактором является доступность биогенных элементов, особенно азота и фосфора. С водорослей начинаются почти все пищевые цепи (планктон, рыбы). Благодаря фотосинтезу поддерживается уровень кислорода в атмосфере, 50 % которого поставляют водоросли. Найдены ископаемые остатки синезеленых водорослей, живших 3 млрд лет назад, а первые организмы, освоившие сушу, возникли лишь 420 млн лет назад. Вероятно, при переходе на сушу главной проблемой было обезвоживание — нужно было выработать приспособления для добывания и запасания воды. Для фотосинтеза и дыхания нужно, чтобы обмен двуокиси углерода и кислорода происходил не с окружающим раствором, а с атмосферой. Проблематично было и размножение растений без воды и питания. На суше фотосинтез происходит над поверхностью земли на свету, а минеральные соли и вода находятся в земле, поэтому часть растения должна быть в темноте под землей, а часть — в воздухе. Кроме того, водная среда обеспечивает постоянство условий внешней среды, а воздух более подвержен изменениям таких параметров, как температура, интенсивность освещения, концентрация ионов в среде и кислотность рН.
В настоящее время выявлены молекулярные механизмы одного из типов фотосинтеза у бактерий. Спектроскописты определили последовательность и временные параметры световых реакций фотосинтеза и скорости взаимодействий. Пикосекундная абсорбционная спектроскопия позволила разрешить временные интервалы до триллионной доли секунды. Интенсивности двух лазерных лучей, проходящих через исследуемую кювету, были столь малы, что не нарушали процессов фотосинтеза, короткая вспышка только инициировала фотосинтез почти одновременно во всех частях исследуемой области. Световой луч контролировал изменение состава образца.
Удалось проследить путь электрона от одной мембраны до другой вследствие поглощения фотона. Специалисты по рентгено-
486
структурному анализу расшифровали пространственную структуру области, где происходят световые реакции, и выяснили взаимное расположение в ней различных молекул. Молекулярные генетики установили локализацию и организацию генов, кодирующих основные компоненты в этой области, так что теперь можно манипулировать этими генами. Д. Юван сумел так изменить их, что получил бактерии, отличающиеся от обычных. Это открывает новые возможности генной инженерии и позволяет досконально понять процессы. Особенностью фотосинтеза этих бактерий было отсутствие выделения кислорода в отличие от зеленых растений, но в фотосинетезе принимают участие те же молекулы хлорофилла. Интерес к этим бактериям связан с тем, что они получают необходимую энергию разными способами, а не только от света.
Хемосинтез — процесс синтеза органических веществ из неорганических за счет энергии химических реакций, протекающих при окислении неорганических веществ. Хемотрофы — бактерии тарификаторы, серобактеры, железобактеры и пр. — в качестве источника водорода используют не воду, а Н2 или H2S, поэтому они кислород не выделяют. И за счет только процесса хемосинтеза аэробные организмы жить не смогли бы.
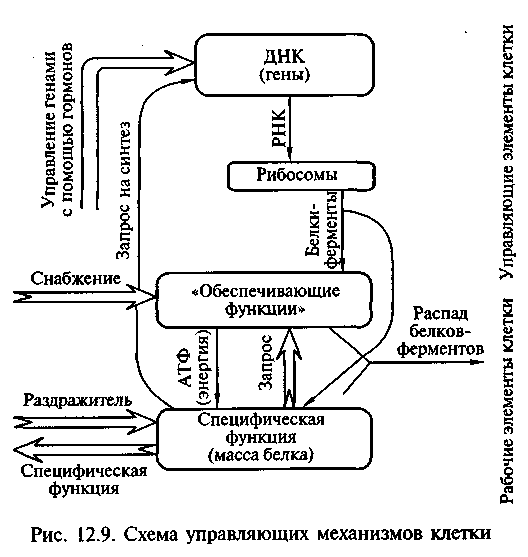
На клеточном уровне организации действуют управляющие ее работой механизмы (рис. 12.9).
487


 12.5. Формирование идей эволюции в биологии
12.5. Формирование идей эволюции в биологииРазвитие во времени — неотъемлемое и характерное свойство живой природы. Идеи единства и развития природы можно проследить с древнейших времен. Проблемы происхождения и эволюции жизни на Земле были в центре философских и религиозных систем. Так, Г.Лейбниц, развивая идеи Аристотеля, провозгласил принцип градации и предсказал существование переходных форм между растениями и животными. В России эти идеи активно поддерживал А. Н. Радищев. В дальнейшем этот принцип был развит в представлении о «лестнице существ» от минералов до человека и Бога, доказывая трансформизм живой природы. Так выявлялась не эволюция, а общность и сходство форм и усложнения организмов. Но сопоставлялись время существования Земли и время формирования форм живого на ней. Бюффон обосновывал в своей «Естественной истории» историю Земли и доказывал единство происхождения живого планом строения. Другой энциклопедист, Д.Дидро, считал, что разнообразие органического мира объясняют мелкие изменения живых существ и длительность существования Земли. У Мопертьюи были догадки о корпускулярной природе наследственности, эволюционной роли вымирания неприспособленных форм и роли изоляции в возникновении новых форм. Кант в «Космогонии» говорил о развитии живого мира в течение миллионов лет.
К. Линней считал, что близкие виды внутри рода могли развиться естественным образом без участия высших сил. Эволюционные представления были характерны для К.Вольфа, М.В.Ломоносова и А. Н. Радищева. Ломоносов считал изменения в неживой природе причиной изменений мира живого, по останкам вымерших форм судил об условиях существования их в далекие времена. Он писал, что время, необходимое для создания организмов, больше, чем определяется церковным исчислением. Но эти идеи формировали пока только представление о последовательности природных тел. Ограниченную трансформацию видов допускал Ж. Бюффон, считая, что разные типы животных имеют разное происхождение и возникли в разное время. Эта концепция обобщала многие наблюдения и факты, выделяла идею глубокой взаимосвязи между видами, подвидами, родами и другими таксонами, подготавливая почву для эволюционизма. Так, до конца XVIII в. господствовала мысль о «целесообразности порядков в природе» (сотворении кошек для пожирания мышей и т.п.). Постепенно возникал вопрос о возникновении такой целесообразности. Общество не могло еще воспринять идеи эволюции, так как обсуждались не доступные для проверки масштабы времен.
Идею эволюции живого перевел на уровень теории эволюции Ж. -Б. Ламарк. Он считал, что Бог сотворил материю и движение, а далее развитие происходило по естественным причинам. Опираясь на многочисленные факты изменяемости видов, Ламарк в книге «Философия зоологии» (1809) выдвинул гипотезу о меха-
488
низме эволюции, основанном на двух предпосылках: наследование приобретенных признаков и упражнение или неупражнение частей организма. Он представил эволюционное обоснование «лестницы существ», основанное на принципах градации (внутреннего стремления к совершенству) и изначальной целесообразности реакции организма на изменения внешней среды (признание возможности прямого приспособления). Далее Ламарк формулировал два закона: 1 — изменение привычек следует сразу за изменением условий и 2 — эти изменения передаются по наследству. Основа эволюции — врожденная способность к самосовершенствованию, фактор явно нематериальный: «творить может только Бог, тогда как природа может только производить», а изменения во внешней среде могут изменить формы поведения, поэтому органы или структуры способны приобрести новые функции, а эти новые функции органов и изменения в них могут быть переданы потомкам. Так вытянулась шея у жирафа, увеличились перепонки у водоплавающих, развивается мускулатура при занятиях спортом. Эта часть учения Ламарка отвергалась, как противоречащая появляющимся новым знаниям о механизме наследования, для нее еще не пришло время. Большую роль в возникновении новых видов Ламарк отводил переменам климата и гидрогеологического режима. Политические страсти внутри биологии скомпрометировали важность идей типа «наследуется все благоприобретенное» (она получила название «ламаркизм»). Как подчеркивал К.А.Тимирязев, Ламарк не сумел объяснить целесообразность организмов. Но «роль Ламарка в биологии колоссальна», — отметил современный генетик Л. Н. Серавин (1994).
Предшественником идей Дарвина в России был зоолог К. Ф. Ру-лье, развивавший идеи возникновения органического мира из неорганического. Он выделял наследственность и изменчивость в качестве основных свойств организмов, говорил и о постепенном изменении организмов под влиянием внешних условий.
Учение катастрофизма отражало идеи развития природы, сформировавшейся к началу XIX в. Французский зоолог Ж. Кювье выделял четыре типа животных — позвоночные, мягкотелые, членистые и лучистые. И с каждым из них он сопоставлял некий «план композиции», некую «творящую силу», которая после очередной катастрофы в геологической истории обеспечивает восхождение органических форм. Тем самым геологическую эволюцию планеты Кювье связывал с эволюцией живого, признавая роль катастроф и неравномерности темпов преобразований в природе, что не потеряло своего значения и поныне. Ему возражал У. Смит, один из основоположников биостратиграфии, отмечавший непрерывность распространения сходных видов в близких по возрасту слоях.
Концепцию униформизма сформировали противники теории катастроф. Дж. Геттон, Ч.Лайель, М. В.Ломоносов критиковали идеи
489



 Кювье за неопределенность причин катастроф, за укорачивание возраста Земли. Под впечатлением успехов классической механики они считали мир познаваемым и предлагали опираться на преемственность настоящего и прошлого, выделяя непрерывность действия законов и факторов в истории Земли и возможность обратимости явлений (см. гл. 9). При эволюции живого Лайель допускал возможность актов творения, демонстрируя прогрессивные изменения ископаемых останков. Униформисты считали, что вымирание несовместимо с естественным образованием новых видов, и потому предполагали участие творца. Фактически они свели историю планеты к цикличности и случайным изменениям.
Кювье за неопределенность причин катастроф, за укорачивание возраста Земли. Под впечатлением успехов классической механики они считали мир познаваемым и предлагали опираться на преемственность настоящего и прошлого, выделяя непрерывность действия законов и факторов в истории Земли и возможность обратимости явлений (см. гл. 9). При эволюции живого Лайель допускал возможность актов творения, демонстрируя прогрессивные изменения ископаемых останков. Униформисты считали, что вымирание несовместимо с естественным образованием новых видов, и потому предполагали участие творца. Фактически они свели историю планеты к цикличности и случайным изменениям.Униформизм, как и ламаркизм и катастрофизм, предварял теорию естественного отбора, частично конкретизировал идею эволюции. Английский экономист Т.Мальтус привлек внимание к репродуктивному потенциалу человека и указал на экспоненциальный рост численности населения. В 1788 г. он опубликовал «Трактат о народонаселении», в котором убедительно и ярко обрисовал, к чему может привести ничем не сдерживаемый рост населения. Благодаря Дарвину выражение Мальтуса «борьба за существование» приобрело широкую известность. Они оба считали ее результатом несоответствия между быстрым ростом популяций и ограниченностью пищевых ресурсов. Идеи Мальтуса и Лайеля оказали большое влияние на Ч.Дарвина. Он верил в познаваемость законов природы, в возможности объяснения наблюдений. Дарвин считал проблему происхождения человека связанной с эволюцией неорганического и органического мира. К середине XIX в. в разных областях биологии был накоплен огромный фактический материал, который нуждался в обобщении. Да и практика сельского хозяйства требовала теории, которая бы открыла пути селекции.
Понятие «эволюция» в биологию было введено швейцарским ученым Ш. Боннэ (1762). Он понимал под этим термином не только саму идею развития, но и отмечал изменчивость и некий отбор в становлении форм живого. Ламарк объяснял изменчивость влиянием наследственности и внешних факторов — питанием, климатом, упражнением органов. Дарвин создал в 1859 г. теорию эволюции, обобщив отдельные эволюционные идеи и разрешив накопившиеся противоречия. У него эволюция определяется триадой: наследственность, изменчивость и естественный отбор.
Остовом теории эволюции путем естественного отбора послужил огромный материал, собранный и до Дарвина, и им самим.
Ч.Дарвин, в юности собирающийся стать пастором, интересовался зоологией как любитель. Он предпринял пятилетнее морское путешествие на корабле «Бигль», во время которого занимался геологическими исследованиями, собирал ботанические, зоологические и палеонтологические коллекции. Кульминацией, с точки зрения формирования его эволюционных взглядов, явилось исследование флоры и фауны Галапагосских островов, где он увидел в действии процесс эволюции при сравнении близких видов вьюрков, ящериц, черепах, о чем и написал в своей первой книге (1839). По прибытии в Англию он проанализировал
490
историю селекции и выявил отличия между породами и сортами. Здесь он усмотрел творческое начало в деятельности селекционеров, позволяющее накопить изменения в результате отбора.
Дарвин собрал обширный материал об изменчивости организмов и видов и отметил почти всегда постоянную численность популяций. Способность к размножению, свойственная всему живому, обеспечивает сохранение вида. Численность популяций на Земле контролируется различными факторами среды (пространство, свет, пища, тепло). Исследуя и сопоставляя огромный материал и находясь под впечатлением идеи Мальтуса, Дарвин начал понимать, что при интенсивной конкуренции между членами популяции любые изменения, благоприятные для выживания в данных условиях, повышают способность особи к размножению и оставлению плодовитого потомства. Но каждый вид производит больше особей, чем выживает их до взрослого состояния, а среднее число взрослых особей почти постоянно. Ненужные формы при этом отбрасываются путем нового механизма — естественного отбора. Черновой вариант своей теории он сделал в 1842 г.
Понятиям изменчивость и наследственность, которые Ламарк связывал с приспособляемостью, передаваемой по наследству и являющейся основой видообразования, Дарвин придал принципиальное значение. Определенная изменчивость — это способность всех особей определенного вида одинаковым образом реагировать на изменения среды, при этом изменения в организмах не наследуются (сейчас это — адаптивная модификация). Неопределенная изменчивость приводит к существенным изменениям в организме, которые наследуются с усилением в следующих поколениях (мутация, по современной терминологии). Она тоже связана с условиями окружающей среды, но не непосредственно. Дарвин считал, что именно такая изменчивость играет ведущую роль в эволюции.
Естественный отбор — механизм эволюции, материал для него — наследственная изменчивость. В нем Дарвин соединил многие биологические знания, в том числе опыт практической селекции.
А. Уоллес, один из основоположников зоогеографии, много путешествовавший по Южной Америке и Юго-Восточной Азии, тоже читал Мальтуса и пришел к идеям, близким к теории Дарвина. Уоллес и Дарвин выступили с сообщениями о роли естественного отбора в эволюции на заседании Линнеевского общества. В 1859 г. Дарвин опубликовал свою книгу «Происхождение видов путем естественного отбора», она разошлась в первый же день, и, говорят, по своему воздействию на человеческое мышление уступала только Библии. Эти идеи вызвали бурные дискуссии в обществе и церкви. Уоллес отрицал приложимость отбора к
491


 «возникновению человеческих способностей», а Э. Геккель, страстный сторонник Дарвина, назвавший его «Ньютоном органического мира», прилагал идею естественного отбора к развитию общества. Ботаник А. Н. Бекетов в работе «Гармония в природе» (1858) привел обширные материалы об изменении растений в разных условиях и выделил их борьбу за существование. Русский князь, географ и геолог П. А. Кропоткин, известный как теоретик анархизма, много путешествующий по Сибири, наблюдал перемещения больших масс животных, спасающихся от стихийных бедствий. На основе этого он выделил в качестве факторов эволюции взаимопомощь и кооперацию.
«возникновению человеческих способностей», а Э. Геккель, страстный сторонник Дарвина, назвавший его «Ньютоном органического мира», прилагал идею естественного отбора к развитию общества. Ботаник А. Н. Бекетов в работе «Гармония в природе» (1858) привел обширные материалы об изменении растений в разных условиях и выделил их борьбу за существование. Русский князь, географ и геолог П. А. Кропоткин, известный как теоретик анархизма, много путешествующий по Сибири, наблюдал перемещения больших масс животных, спасающихся от стихийных бедствий. На основе этого он выделил в качестве факторов эволюции взаимопомощь и кооперацию.Так пришли к представлению о том, что органический мир представляет некое единство, имеет свою историю, а его нынешнее состояние есть результат предшествующего. Заслуга Дарвина в том, что из сопоставления фактов борьбы за существование и всеобщей изменчивости свойств и признаков он вывел неизбежность избирательного уничтожения одних особей и размножения других — естественного отбора. Начинаясь с наблюдения, познание жизни продолжалось на уровне мыслительных процессов. В классической биологии эксперимент еще не был методом познания живого. Механистический детерминизм игнорировал функциональное единство живых систем, а телеологический подход основывался на целесообразности организмов. С теории эволюции Дарвина, в основе которой лежал рациональный подход, началось преодоление идеалистической тенденции в биологии.
Учение Дарвина (наследственность, изменчивость и естественный отбор) за несколько лет вытеснило все антиэволюционные и креацианистские концепции. При этом сопоставляли данные палеонтологии, сравнительной анатомии и эмбриологии (метод Г е к к е л я). Данные палеонтологии доказывали существование эволюции живого. Это показал еще В. О. Ковалевский на примере развития вида лошадей, обнаружив существование предка с пятипалой конечностью, жившего 60 млн лет назад. Последовательные ряды ископаемых животных он выстроил в ряд для наглядности эволюционных изменений. (Такие ряды называют филогенетическими.) До конца XIX в. эволюционные идеи овладевали умами, строились филогенетические древа для всех крупных групп растений и животных. Существуют и эмбриологические доказательства эволюции. Закон Геккеля — Мюллера утверждает, что каждая особь в своем индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез). К доказательствам относят и наличие рудиментальных органов, и явления атавизма. Существуют и биогеографические доказательства: сравнение животного и растительного мира разных континентов показывает, что различия внутри вида тем больше, чем дольше длилась их изоляция. Биоразнообразие — неизбежный результат отбора.
492
В результате изучения эволюции групп (макроэволюции) сформировался так называемый классический дарвинизм: установлена необратимость эволюции, принцип мультифункциональности органов и эволюции органов путем смены функции, биогенетический закон и др. Но многие ученые не приняли дарвинизм, и это неприятие и критика особенно усилились в период возникновения генетики. Можно даже сказать, что распространение эволюционных идей сопровождалось в это время острой критикой теории естественного отбора. Отсутствие обнаружения переходных форм препятствовало принятию дарвиновских идей, а с появлением генетики естественный отбор все больше подвергался критике. Так, Ф.Дженкин показывал, что при скрещивании произойдет «растворение признаков», которые были единично поддержаны отбором. Основу такой критики составило отсутствие строгих доказательств наследственной изменчивости, которые давала генетика (теория мутаций, учение о чистых линиях и принцип корпускулярной наследственности). В. Л. Иоганнсен показал неэффективность отбора в чистых линиях (в потомстве одной са-моопл од отворяющейся особи). Г.де Фриз в 1889 г. выдвинул мутационную гипотезу о скачкообразном возникновении новых видов путем крупных изменений наследственности (мутации) без ведущего участия естественного отбора.