
Урбанизация «диких» видов птиц в контексте эволюции урболандшафта
| Вид материала | Документы |
- Красная книга, 61.32kb.
- Тайны диких животных, 61.83kb.
- Редкие и находящиеся под угрозой исчезновения виды диких животных и дикорастущих растений, 3127.94kb.
- Венции должны содействовать восстановлению исчезающих видов, в частности, путем разработки, 98.37kb.
- Правила поведения в лесу: Не лови диких животных и не уноси их домой, 321.38kb.
- Правительство Республики Казахстан постановляет : Утвердить прилагаемую сохранения, 754.5kb.
- «Санитарно-эпидемиологическрое значение мошек (сем. Simuliidae) как переносчиков возбудителей, 86.93kb.
- Правительство Республики Казахстан постановляет: Утвердить прилагаемую программу сохранения, 712.05kb.
- «Красной книге», 31.74kb.
- Классный час на тему: «Многообразие пернатых», 88.89kb.
В результате парадоксального сочетания «проточности» популяционных группировок с очень жёсткими территориальными связями «ядра» резидентов, в каждой группировке поддерживается определённое пропорциональное соотношение между особями-приверженцами альтернативных стратегий (их бывает две и более).
Эта пропорция представительства альтернативных стратегий в конкретной популяции остаётся постоянной и тогда, когда другие популяционные характеристики (численность, персональный состав резидентов, ресурсообеспеченность особей) резко и непредсказуемо колеблются. Дело в том, что она отражает тот аспект разнокачественности особей в популяции, который существен для приспособления популяционной системы к наиболее общим характеристикам новой среды обитания - города.
Третий характерный признак городских популяций - расщепление на несколько альтернативных стратегий, поддержание динамического равновесия между ними, быстрые и адекватные сдвиги равновесия в ответ на направленные изменения среды. Описанная выше дифференциация групп «сверхоседлых» и «сверхподвижных» особей в городских популяциях популяции с противоположным типом реагирования на текущие изменения среды может рассматриваться как основа для формирования альтернативных стратегий, так как зафиксирована у всех «городских» видов.
В наиболее развитом виде стратегии включают не только тип реагирования особи, но также сопряжённые с ним поведенческие, психофизические или морфологические показатели. Поэтому разные стратегии могут рассматриваться как альтернативные варианты специализации особей к противоположным вариантам ландшафтного, топического и т.п. разнообразия урбосреды, которые образуют противоположные полюса «городского градиента» (рис.1). При всей гетерогенности структуры урболандшафта разнообразие мозаик среды «выстроено» по градиенту «центр-периферия города», вдоль него падают уровень урбанизации и концентрации главных элементов техногенного ландшафта (застройка, дороги и т.п. коммуникации), а концентрация участков природных сообществ растёт (Geyer, Kontuly, 1993; Witt, 1996).

Рисунок 1. Городской градиент: падение степени урбанизации территории ( 1) и степени изменённости городом природных ландшафтов, представляющих собой естественные местообитания вида (2-4) от центра города (I) к его границе (II) и далее к периферии зоны влияния города, III. IV – радиус центральной агломерации и потенции его изменения в процессе роста урбанизированного «ядра» (штрих-пунктирные стрелки). Пунктирная стрелка – вектор изменений урбосреды в процессе городского развития,
Трансформация видовых местообитаний после попадания в зону воздействия города и включения соответствующих биотопов отдельными участками в структуру урболандшафта (на следующем этапе расширения города). 2 - фрагментация непрерывных местообитаний вида в «зоне влияния» города, образование «островного ландшафта». 3 - изоляция «островов» улицами, коммуникациями, городскими кварталами, усиление «краевого эффекта», сокращение площади отдельных «островов» и увеличение их изолированности. 4 - дробление и разъединение «островов», переход от закономерного расположения «островов» относительно друг друга к целиком стохастическому.
Figure 1. Urban gradient as declining of the territory urbanization degree ( 1) & habitat transformation degree (2-4) from center (I) to periphery (II) the urbanized area. Out of boundaries the city urban gradient extend to the impact zone of city (III) as a regional urbanization gradient. IV – central conurbation radius and tendencies of their change during urbanization process continued in this region (stroke-dotted arrows). Dotted arrow – the direction of urbanized territories growth during urbanization process which connected with extension of urbanized areas and habitat transformation in its impact zones.
2-4 – different characteristics of habitat transformation of some species after falling into urbanized area impact and including some parts of these habitats in structure of urbanized landscape on the next stage of city expansion. 2 – Fragmentation of continual habitats in impact zones the cities with forming the «island landscape”, 3 - isolation of several «islands» by streets, others communications, buildings, increasing the «edge effect», decreasing the «island’s» area and growth the isolation degree of every «island». 4 – crushing and separation the «islands», transition from natural to stochastic location different «islands» comparatively each other.
Для устойчивых городских популяций выделение пар или троек альтернативных стратегий, их распределение между особями в популяции также типично, как и появление связей каждой из альтернатив с определённым полюсом городского градиента. С одной стороны, это плотно застроенный центр города, с другой - «полусельские», «парковые» окраины (естественно, в случае кольцевой структуры города; при многоядерной картина усложняется).
Там, где это исследовано специально, принадлежность особей городских популяций к одной из альтернативных стратегий было связано (коррелятивно, но жёстко) с принадлежностью к одной из нескольких цветовых морф или, как минимум, к одному из противоположных психофизиологических типов. Последние проявляются в альтернативных реакциях на присутствие и/или активные действия других особей, особенно в условиях переуплотнения. Пример противоположных психофизиологических типов, соответствующих альтернативным стратегиям – «бродячие» и «территориальные» особи большой синицы, «агрессивные» и «толерантные» птенцы сизаря и т.п. (Ксенц, 1990; Москвитин, Гашков, 2000).
Так, в городских популяциях сизаря существуют три альтернативные стратегии «летунов», «собирателей» и «помоечников». В смешанных колониях «летуны» размещаются наверху «лестницы иерархии», «собиратели» – посредине, а «помоечники» – в самом низу. Первые кормятся зерном на комбинатах зернопродуктов, пригородных фермах и пр., летают на расстояние до 15 км от колоний. Вторые подбирают корм с земли в местах скопления людей, третьи посещают свалки и помойки не далее 200-400 м от колонии. «Собиратели» много расхаживают при сборе корма, а «помоечники» малоподвижны (Ксенц, Москвитин, 1983).
Этому делению взрослых птиц соответствуют два противоположных типа защитного поведения птенцов в городских колониях сизаря – агрессивная или спокойно-безразличная реакция. Агрессивной реакции птенцов и взрослых соответствует «боязливое» отношение к исследователю в противоположность безразличному или доверчивому отношению, с минимальной дистанцией вспугивания человеком. Птенцы с «агрессивным» типом реагирования на наблюдателя преобладают в плотных колониях на светлых и достаточно объёмных чердаках старых зданий. Они объединяются в «ясли» из потомства 4-5 пар, кормящих всех птенцов в группе без разбору, чем увеличивают эффективность приноса корма остальными парами. Так возникают крупные, но довольно разреженные колонии.
Напротив, на тесных и тёмных чердаках новых зданий птенцы практически не покидают гнёзд и не перемещаются по чердаку, иначе родители не смогут отыскать их. Здесь голуби кормят только своих птенцов. Возникают небольшие, но очень плотные колонии, в которых получают преимущество птенцы и взрослые со спокойно-безразличным типом реакции (Ксенц, Москвитин, 1983; Ксенц и др., 1987).
Важно подчеркнуть, что соответствующие реакции почти не зависят от поведения самого человека. Тем самым подтверждается предположение, что альтернативность психофизиологических типов свойственна самим особям в популяции.
Спокойный вариант поведения птенцов чётко преобладает в переуплотнённых крупных колониях. Он сильно снижает потери, вносимые социальной дезорганизацией, увеличивает устойчивость репродукции резидентов в условиях постоянных помех со стороны соседей. Соответственно, птенцам «сизой» морфы наиболее свойственен «агрессивный», взрослым - «боязливый» тип реагирования. Среди «боязливых» птиц преобладают «летуны», среди «доверчивых» и «безразличных» – «помоечники». «Летуны» могут переходить от питания зерном на зернопунктах к сбору пищевых отходов на «микросвалках», расположенных вблизи колонии, вслед за брачным партнёром, придерживающимся стратегии «помоечника». Переход «помоечников» в «летуны» не происходил никогда.
В томской популяции больших синиц особи на протяжении всей жизни (несколько лет) сохраняют тот уровень жёсткости территориальных связей в период зимовки, присущий именно и только им. Эта характеристика индивида не меняется в ответ на изменения плотности зимовочных скоплений и/или условий зимовки. Особи в популяции чётко распределяются в две противоположные категории подвижных и «сверхоседлых» птиц (Гашков, Москвитин, 2001). Так, 16,2% птиц (n=1094) зимовали буквально вокруг одной точки в пространстве парка, чаще всего вблизи прикормочных мест. Их не отмечали в других точках контроля в самом парке или за его пределами. По характеру использования пространства они соответствуют «помоечникам» сизаря. 71,6% особей постоянно отмечали на площади 2,2-4,5 га: подобно «собирателям», они постоянно перемещаются с одного места кормления на другое. 12,1% птиц постоянно пребывали в пределах пространства в 20-27 га. Жизненная стратегия этих синиц, а также битерриториальных P.major соответствует «летунам» – дальние вылеты и длительное кормление на удалённых источниках корма.
Битерриториальные P.major имеют (и защищают) две постоянные территории, одну на территории города, другую в загородном лесу (в малом и среднем городе). Другой вариант битерриториальности реализуется в крупных городах: первая территория особи располагается в городском лесу, вторая – в центральных кварталах. Везде обе территории разделены расстоянием в несколько км: птицы ежедневно или раз в несколько дней перелетают с одной на другую, не останавливаясь в промежуточных точках (Гашков, Москвитин, 2001). Поэтому возможно говорить об альтернативных стратегиях как неких дискретных типах реагирования индивидов в популяционной системе на изменения среды.
Помимо разного размера используемого пространства, большие синицы томской популяции различаются по регулярности перемещений. 40,6% птиц образуют группу постоянно кочующих (частью входит в группы «собирателей» и «летунов»). В местах кольцевания они регистрировались непостоянно, периоды отсутствия в течение зимы составляли от 23 до 42 дней. «Кочующие» особи имели разное число зимовочных мест, и периодически жили то в одном, то в другом, преодолевая разделяющее пространство «рывком». «Подвижные» и «оседлые» особи чрезвычайно привязаны к используемым территориям. Они редко встречаются на удалении более 200 м от точки кольцевания, просто у «подвижных» перемещения идут постоянно и территорий более чем одна (Москвитин, Гашков, 2000).
Вообще, что взаимоотношения в контексте заботы о потомстве (внутри выводка с одной стороны, между родителями и потомством с другой) у птиц организованы так, так что дают «на выходе» высокий уровень разнокачественности молодых, распределяя птенцов по альтернативным стратегия, условно «быстрой» и «медленной» (см.ниже).
У птиц взаимодействия птенцов в выводке и родителей с птенцами организованы так, что поведенческая разнокачественность потомства направленно усиливается социальными средствами сверх того уровня, который задан биологической разнокачественностью самих птенцов. Взаимная конкуренция птенцов за получение корма приводит к дифференциации двух альтернативных стратегий, условно «быстрой» и «медленной», в наиболее ярком виде проявляющихся у двух «крайних» птенцов (самого подвижного и сообразительного и «самого тупого, стереотипного» по описанию птенцов ворона Берндтом Хейнрихом, 1994) а все прочие распределяются между ними. Подкрепление от родителей (положительное – в виде приносов пищи, отрицательное – в виде периодической подачи песни, заставляющей насторожиться, и сигналов тревоги, заставляющих затаиваться) воздействует на птенцов так, что подкрепляет дифференциацию стратегий разных птенцов и стимулирует каждого птенца и дальше специализироваться в раз избранной стратегии, не останавливаться и не менять её. Детали процесса подробно описаны в исследованиях С.Н.Хаютина и Л.П.Дмитриевой (1981, 1991), выполненных в основном на мухоловке-пеструшке.
В итоге к моменту вылета поведенческая разнокачественность птенцов максимальна и в целом соответствует таковой взрослых особей в размножающейся популяции. В ходе взаимодействий в гнезде птенцы конкурируют за первоочерёдное получение корма, и в ходе конкуренции дифференцируются две противоположные стратегии, условно говоря «быстрая» и «медленная». «Быстрые» птенцы сильны, активны, не боятся новизны (в том числе готовы получать корм, не пугаясь тревожных криков родителей, шорохов, сотрясений гнезда и т.п.). Они первыми прорываются к летку, перемещаясь через всё дно дуплянки, первыми получают корм, наедаются и засыпают, освобождая место.
Это, так сказать, конкурентная стратегия: птенец сам, в меру своих сил формирует наиболее благоприятные обстоятельства для получения корма, не боясь изменчивости и нестабильности внешнего мира. Альтернативная стратегия – толерантная: птенцы, проигрывающие все попытки рвануться и взять корм первыми, и постепенно привыкающие максимально использовать для приёма корма те отрезки времени, когда наиболее конкурентные птенцы уже наелись и освободили леток или край гнезда. Для этого они не перемещаются и почти всё время сидят под летком, то есть пассивно ждут благоприятных обстоятельств и, когда возникает соответствующая стимуляция (прилетает родитель с кормом) стереотипно реагируют на воздействие.
В начале периода выкармливания конкурентная стратегия резко выгоднее толерантной, и вторые птенцы сильно проигрывают в весе. Но затем ситуация выравнивается и к моменту вылета представители все птенцы имеют примерно одинаковый вес, но стратегии поведения (выпрашивания корма и реагирования на сигналы родителей) оказываются максимально дифференцированы. Намного больше, чем можно предположить, исходя из биологической разнокачественности птенцов в начале периода выкармливания.
Возникает своеобразная «карусель», непрерывный «кругооборот птенцов в гнезде»: индивиды последовательно сменяют друг друга у летка, почти не вступая в физическое столкновение, так что представители «более толерантных» стратегий используют периоды отдыха и сытости птенцов «более конкурентных» стратегий. Поскольку родители активно поддерживают эту систему, подкрепляя кормом всякий шаг на пути дифференциации, логично считать, что разнокачественность птенцов, отражённая в дифференциации стратегий, не дана изначально, а создана социальными средствами. Этот постоянный «круговорот птенцов в гнезде» ведёт к тому, что пространство внутри гнезда оказывается анизотропным и птенцы с разной стратегией занимают разные «позиции» относительно центра приёма пищи – летка или края гнезда. Чем «быстрее» стратегия птенца, тем дальше он находится от летка, когда сыт, и быстрей пробивается к нему, когда голоден. «Медленные» особи всё время находятся под летком.
Этот «кругооброт», дифференцирующий птенцов, впервые был показан для дуплогнёздника мухоловки-пеструшки, птенцы которой находятся в дуплянках, где имеется один чёткий центр принятия пищи – леток и есть возможность дать чёткий сигнал о прилёте родителей уменьшением освещённости от закрытия летка и сотрясением стенок от удара о дуплянку. Но и у открытогнездящихся птиц происходит та же самая дифференциация птенцов на «быстрых» и «медленных». Непосредственно перед вылетом выделяется «смышлёный и шустрый» птенец, готовый во всё лезть и всё исследовать, его противоположность – самый тупой птенец выводка, боящийся новизны и стереотипно реагирующий на стимуляцию. А прочие птенцы находятся посредине. В частности, это описано для птенцов ворона Берндтом Хейнрихом (1994).
Эти поведенческие различия птенцов сохраняются, но не усиливаются у взрослых птиц, проявляясь как альтернативность «быстрых» и «медленных» фенотипов птиц в природных популяциях. На больших синицах (Parus major) показано, что по показателям локомоторной подвижности и ответа подвижностью на новизну в размножающейся популяции выделяются те же стратегии, что у птенцов. У «быстрых» особей в ответ на новизну возрастает подвижность и исследовательские реакции, у «медленных» они, наоборот, подавляются (Drent et al., 2003; Dingemanse et al., 2002, 2003; ссылка скрытассылка скрыта).
«Быстрые» и «медленные» фенотипы у синиц могут быть разделены, например, тестированием в «открытом поле» по методу Dingemanse et al. (2002), или при помощи метода «удвоения вольеры», когда поведенческая стратегия особи проявляется в скорости освоения вдруг появившегося нового пространства (Ильина и др., 2006).
В двойных вольерах вероятность размножения была выше у «быстрых» особей, активней осваивающих новое смежное помещение. У самцов скорость освоения новой территории положительно коррелировала с результатом тестирования в «открытом поле». В отличие от вероятности размножения его успешность и сроки не зависели от размера вольеры (одинарная или удвоенная), но были связаны с результатами тестирования в открытом поле, то есть с дихотомией «быстрых» и «медленных» фенотипов. «Быстрые» самки раньше медленных начинали демонстрировать репродуктивное поведение, но позже приступали к откладке яиц и хуже насиживали отложенные яйца. Влиял и фенотип самца: партнёрши «быстрых» самцов раньше проявляли репродуктивное поведение, «медленных» самцов – раньше начинали насиживать кладку. Наконец, превосходство самца над самкой по показателям тестирования в «открытом поле» повышало вероятность проявлений репродуктивной активности самца в предгнездовой период (Иванкина и др., 2006).
То есть в социальных взаимодействиях животных в сообществе «быстрые» особи лучше стимулируют партнёра, но хуже (менее точно, с большей вероятностью ошибок и сбоев) реализуют свою социальную роль. А в репродукции и социальной коммуникации точность реализации видовых форм сигналов и форм отношений не менее важна, чем сила и интенсивность стимуляции между партнёрами. Разделение на «быстрых» и «медленных» особей в популяции хорошо согласуется с «моим» разделением особей на приверженных конкурентной и толерантной стратегиям как двум взаимоисключающим альтернативам. Первые предпочитают высокую социальную плотность и чувствительных к энвироментальному стрессу, вторые - низкую плотность, неконкурентоспособных в условиях социального стресса.
В голландской популяции больших синиц также показано, что с альтернативностью «быстрых» и «медленных» фенотипов связана также альтернативность стратегий социального, кормодобывающего и репродуктивного поведения – всего того, что требует реакции на новизну, умения контролировать ситуацию, не боясь риска, связанного с этой самой новизной (или наоборот, избегания новизны и самоограничения средой где можно вести себя только стереотипно), см. Drent et al., 2003; Dingemanse et al., 2003.
Авторы, исследовавшие только взрослых особей, полагают дихотомию «быстрых» и «медленных» фенотипов наследственной. Наследуемость была определена косвенным методом и составила по методу регрессии «родители-потомство» 0,22-0,41, по данным анализа сиблингов 0,37-0,41 (Dingemanse et al., 2002).
Однако в исследованиях онтогенеза видно, что обе альтернативные стратегии – не врождённые, а «сделанные», дифференцирующий эффект сигнальной наследственности, социальных влияний внутри выводка и от родителей здесь «мимикрикует» под эффект генетической наследственности. С одной стороны, конкурентные взаимодействия в выводке организованы так, что создают «зародыш» дифференциации стратегий в виде ситуативных уклонений поведения в ту или иную сторону у птенцов, волей случая оказавшихся в определённой роли. С другой стороны, родители положительной и отрицательной стимуляцией (принос корма, заставляющий двигаться – пение, заставляющее насторожиться - тревожный крик, заставляющий затаиваться) воздействуют на дифференцирующееся поведение птенцов в сторону «усиления флуктуаций», оформления и закрепления их в виде дифференцированных стратегий, следование которым с углублением специализации для каждого птенца оказывается выгодней (=эффективней в плане получения корма и минимизации общего риска разорения гнезда), чем корректировка и смена стратегии. Это закрепляет и усиливает общую схему дифференциации внутри выводка.
То есть родительское поведение у птиц как бы «расталкивает» птенцов по разным стратегиям, дифференцирующимся в конкурентных взаимодействиях внутри выводка. Соответственно, выводки у птиц работают как разнообразящие системы, создающие и закрепляющие дифференциацию поведенческих ролей молодых между двумя полюсами альтернатив, существенных для данного вида и популяции. Воздействие родителей на естественную конкуренцию птенцов в процессе развития создаёт положительную обратную связь, поощряющую уклонение моделей поведения от средней на каждом из последовательных этапов созревания. В первую очередь имеется в виду двигательная активность, поведение выпрашивания и затаивания, с соответствующей вокализацией.
Когда альтернативность жизненных стратегий выражена особенно резко, приверженность особи одной из альтернатив обычно маркируется принадлежностью к одной из n цветовых морф, присутствующих в полиморфной популяции. Это может быть ряд от «сизых» к «черночеканным» особям сизаря, от светлых к тёмным самцам мухоловки-пеструшки и пр.
Связь стратегии и цветовой морфы коррелятивная, но жёсткая: особи с «несовпадением» типа окраски и стратегии поведения (+психофизиологического типа) избирательно вытесняются из данной группировки и переселяются туда, где соответствующая корреляция менее жестка. Это всегда поселения на менее урбанизированных территориях (сизарь, Ксенц и др., 1983, 1987, 1990; мухоловка-пеструшка, Куранов и др., 1991; Куранов, 2005).
Главное, что непрерывный ряд цветовых морф в полиморфной популяции может быть поставлен во взаимнооднозначное соответствие с рядом жизненных стратегий индивидов, точней градиентом противоположно направленных изменений концентрации каждой из альтернатив в тех «пропорциях», в каких они присутствуют в локальных поселениях вида. Соответственно, жизненные стратегии особей, существующие в данной популяции, образуют градиент переходов от одного способа решения индивидом проблем социальной (и экологической) адаптации до другого полярного ему, но на равных бытующего с ним в городской или иной популяции.
Соответственно, в популяциях сизаря или мухоловки-пеструшки цветовая морфа особи обращается в сигнал вероятной принадлежности особи к определённой стратегии. Сизари-«помоечники» с высокой вероятностью принадлежат к меланистической или «черночеканной» морфе, «собиратели» – к «черночеканной» же, но с меньшей вероятностью, «летуны» - к «дикому типу» окраски. Даже в исходно гомогенной популяции городская среда способствует выделению противоположных стратегий, она «разводит» приверженцев разных стратегий по районам с полярным уровнем урбанизации (сизарь, Ксенц, 1990).
Значит, приспособленность черночеканных сизарей определяется в основном социальной компетентностью, особенно в условиях интенсивного социального пресса, а птиц «дикого типа» – внешними условиями, при которых происходит размножение. Первые и вторые равно реализуют стратегии, противоположные по структуре поведения и характеру репродуктивного усилия индивида. Развитие поведения молодых особей обнаруживает чёткое тяготение к одному из двух «полюсов». «Помоечники» могут дольше кормиться в холодный период года, но потери от гнездовой смертности у них намного выше.
Репродуктивный выход черночеканных особей максимален в самых урбанизированных районах города. Здесь колонии относительно невелики, но переуплотнены и зависят от корма, собранного в близлежащих кварталах. Связь репродукции этой морфы со степенью урбанизации территории будет заметнее, если для анализа использовать суммарный успех размножения за всю жизнь особи, а не только за отдельный сезон. Успех размножения сизарей «дикого типа» максимален в крупных колониях на окраине, зависящих от зерна зернопунктов, пивоваренных заводов и пр. (Ксенц, Москвитин, 1983; Ксенц и др., 1985, 1987).
Поэтому на окраине города наибольший вклад в воспроизводство популяции вносят особи с синдромом «сизый тип окраски – летуны – боязливые», а птицы с синдромом «спокойно-толерантные – помоечники – черночеканные» - в урбанизированном городском центре. Пропорциональное соотношение трёх стратегий в городских популяциях сизаря (и в отдельных колониях) постоянно колеблется вслед за динамикой урбосреды. Уплотнение застройки, смена ветхих зданий современными, и урбанизация городских окраин увеличивают долю птиц с максимальной «суммой признаков» толерантности к урболандшафту, в первую очередь «спокойных» и «черночеканных». «Сизые», «боязливые» и «летуны» вытесняются на окраины и на новые территории, «отхваченные» у региона при территориальном росте «городского ядра».
Благодаря этому процессу постепенного перераспределения птиц разных стратегий городская популяция оказывается такой же открытой системой, как и сам город, и способной к захвату новых пространств почти одновременно с ним. Одна из альтернативных стратегий имеет полное преимущество над другой в тех районах города (на тех отрезках городского градиента), которые по степени урбанизации и другим характеристикам среды (в первую очередь это связано со скоростью обновления средовой мозаики вокруг поселений вида) соответствуют её собственному оптимуму.
Средовой оптимум каждой отдельной стратегии, в свою очередь, соответствует одному из полюсов общего городского градиента, если вид заселил весь урбанизированный ареал, как сизарь, чёрный дрозд, или полюсам частных биотопических градиентов среды в том типе местообитаний, которые вид заселяет в городе. Это может быть градиент плотности застройки (существенный для гнездящихся на зданиях городских ласточек и хохлатых жаворонков, использующих незастроенные пустыри), или градиент фрагментированности «островов» естественной растительности внутри города (Degen, Otto, 1996; Witt, 1996). Второй градиент значим для таких видов, как зеленушка, мухоловка-пеструшка и пр.: по нему они проникают до самого центра города, используя даже микрофрагменты растительности (куртины голубых елей у административных зданий, группы старых деревьев во дворах и пр.).
Соответственно, каждому отрезку городского градиента (рисунок) присуще своё оптимальное соотношение между альтернативными стратегиями. Оно устойчиво сохраняется в поселениях вида, расположенных в соответствующем районе города, пока урбосреда сохраняет относительное постоянство, и направленно меняется после изменений урбосреды, в первую очередь при любых изменениях городской планировки.
Например, при сносе старых и строительстве современных зданий исчезают крупные колонии сизаря, зависящие от дальних источников корма. Они заменяются россыпью мелких колоний, зависящих от локальных и рассредоточенных источников корма. Важно подчеркнуть, что в составе соответствующих колоний «летуны» сменяются «помоечниками» практически немедленно, без какой-либо задержки во времени (Ангальт, 1989; Ксенц, 1990).
Вообще, рассматривая проблему теоретически, во всех популяциях, расселённых достаточно широко, возникает расслоение особей на приверженцев двух альтернативных стратегий (поведенческих стратегий или, шире, жизненных стратегий, если дифференциация поведения «тянет» за собой психофизиологию и морфологию). Первую наиболее общо можно описать предпочтение социального стресса энвиронментальному, предпочтение стабильных условий среды и высокой социальной плотности, предпочтение конкуренции за социальный ресурс уклонению от конкуренции (получить выигрыш важнее уменьшения риска – конкурентная стратегия).
Скажем в полиморфных популяциях мухоловки-пеструшки Ficedula hypoleuca это самые яркие чёрно-белые самцы. Альтернативная стратегия - предпочтение энвиронментального стресса социальному, разреженных поселений с нестабильными и экстремальными природными условиями плотным группировкам со стабильными условиями только потому что носители этой стратегии при высокой плотности, высокой интенсивности контактов проигрывают "конкурентным особям". Здесь уменьшение рисков, связанных с социальной конкуренцией, важней того выигрыша в виде большей предсказуемости поведения, предсказуемости репродуктивного успеха, который она может дать (настолько важней, что животные терпят и эффективно используют непредсказуемость экологической среды – патиентная стратегия). У той же мухоловки – пеструшки это светлые самцы мухоловки-пеструшки, окрашенные очень сходно с самками (Иванкина и др., 1995, 1996, 2007; Гриньков, Керимов, 1998).
Реально, конечно, есть два полюса - предпочтение социального стресса и предпочтение энвиронментального, со всей гаммой переходов между ними. Но по мере того, как ареал расширяется, градиент центр-периферия становится всё более выраженным, специализация особей в популяциях центра и периферии к одной или другой стратегии становится всё более выраженной. То же самое в случае высокой колеблемости природных условий в ареале одной и той же популяции: если "благоприятные" сезоны размножения периодически сменяются "плохими", а плотность популяционных группировок резко колеблется, расслоение полиморфной популяции на приверженцев двух альтернативных стратегий должно усиливаться, доля особей с переходной и смешанной стратегией - уменьшаться.
Соответственно, в историческом центре расселения в оптимуме ареала преобладают предпочитающие социальный стресс, и те самые предсказуемые удары током (специфическая стимуляция от демонстраций партнёра, конкурирующего с данной особью за социальный ресурс - более сильный травмирующий агент, чем удары током). "Патиентные" особи проигрывают конкуренцию и из переуплотнённых группировок вытесняются в разреженные, из центральных популяций в периферийные. Там энвиронментальный стресс выше, а социальный - пониже. Сходная дифференциация, конечно, бывает и в человеческих популяциях, что хорошо видно на каждой обширной стране. В России были те, кто предпочитал оставаться на месте и служить, тянуть лямку – неважно, чиновником ли, крестьянином и парторгом, и те, кто уходил «приискать новых землиц», в Сибирь, на Дон, на Керженец и Иргиз, предпочитая неустойчивость жизни мощному давлению социума. Интересно посмотреть изменения доли тех и других типов личностей в населении в связи разной степенью устроенности быта и упорядоченностью жизни в РФ, со всем тем, что называется «цивилизацией». Предварительный анализ такого рода провёл А.С.Мартынов (1998) для природоохранных задач (он эти человеческие типы называет «консервативный» и «деятельный»).
В результате при равной ресурсообеспеченности репродуктивный успех за жизнь у ярких приверженцев той и другой стратегии в целом оказывается одинаковым. Но в явном проигрыше - "промежуточные" особи, ещё не специализировавшихся в сторону следования одной или другой стратегии (что мы видим у той же мухоловки-пеструшки, Гриньков, Керимов, 1998). Равенство репродуктивного успеха носителей альтернативных стратегий сохраняется лишь пока популяция благополучна - если условия ухудшаются, но численность стабильна, репродуктивный успех "патиентов" падает сильней чем у "конкурентных" особей, если же начала падать численность популяции, разрушаться структура ареала - то наоборот).
Такая вот модель сбалансированного полиморфизма получается. Сначала расселение особей создаёт градиенты социальной плотности группировок (=социального стресса при конкуренции особей за социальный ресурс вроде распределения самок или дележа территорий) и градиент экстремальности среды обитания (=энвиронментального стресса). Оба градиента, кстати, есть оценка среды "с точки зрения данного вида", с точки зрения необходимости для особей максимально реализовывать свой репродуктивный потенциал в конкуренции друг с другом, а отдельным группировкам и сети группировок в определённой местности - устойчиво воспроизводить соответствующий паттерн структуры.
Появление градиентов вызывает специализацию поведения в отношении двух противоположных стратегий, нарастание крутизны градиентов при расселении ускоряет дифференциацию. Далее дифференциация стратегий начинает работать на устойчивость системы в целом, так как для каждого состояния экологической и социальной среды есть некое оптимальное сочетание двух альтернативных стратегий, которое решает задачу устойчивого воспроизводства паттерна пространственной и социальной структуры лучше, чем "чистые" группировки (т.н. эволюционно-стабильные стратегии по Мэйнарду Смиту, Maynard Smith, 1988).
И как только дифференциация особей начинает работать не только и не столько на приспособленность самих особей, но и на устойчивость воспроизводства структуры системы в целом (ещё точней, первое сопрягается со вторым в каждой отдельной популяции и во всей популяционной системе вида в пределах ареала), сам процесс устойчивого воспроизводства системы в череде поколений обратно усиливает дифференциацию. Этим он способствует ещё большему обособлению альтернативных стратегий и сортировке их носителей между популяциями на разных участках градиента, а внутри популяции - между группировками.
Иными словами, эффект сортировки особей с разными поведенческими потенциями (к развитию разных стратегий) между группировками и популяциями, различающимися положением на том и другом градиенте, точно повторяет эффекты отбора особей с уже выраженными стратегиями, и специализированным поведением. Но сортировка особей с разными потенциями много лучше отбора особей, уже специализировавших поведение.
Лучше тем, что тот же результат достигается много быстрее, в разы, и с существенно меньшим грузом элиминации. Особь, элиминируемая отбором, гибнет или устраняется из размножения, - одни минусы, никаких плюсов. Особь, перемещаемая внутрипопуляционным процессом сортировки конечно, больше рискует по сравнению с резидентами, оставшимися жить там же, где пытались закрепиться, перемещение всегда связано с повышенным риском, но зато эти особи с высокой вероятностью перемещаются именно в ту среду, которая наиболее благоприятна для развития именно их потенций. "Патиенты" - в среду разреженных поселений с высокой колеблемостью экологических условий, "конкурентные особи" - в плотные группировки со стабильными условиями и высокой частотой контактов. Очевидно, тот же самый процесс сортировки особей разного качества между поселениями разных типов связывает группировки потоком нерезидентов, направленно доставляя нужных особей (с нужной жизненной стратегией, психофизологией, экоморфологией и пр.) в "нужное место" внутри популяционной системы, чем способствует адаптивной специализации особей и эволюционной оптимизации всей системы одновременно.
Так что если мы хотим понять механизмы первого и второго, и главное, способ сопряжения "индивидуальных" интересов с "нуждами целого" в реальных популяциях, надо анализировать сортировку вместо отбора.
- Опережающая реакция на сигналы среды как фактор устойчивости городских популяций
Благодаря столь эффективной регуляции перераспределением особей внутри группировок все урбанизированные популяции способны к опережающей реакции на сигналы среды - предвестники долговременных изменений значимых для вида элементов урболандшафта. То есть в сформировавшихся городских популяциях индивиды отделяют сигналы трендов, от флюктуаций и в среднем реагируют только на первые. Например, сизые голуби часто покидают заранее прежние места гнездования, не дожидаясь исчезновения отдельных кормовых точек. В г.Перми много лет подряд местом гнездования голубей было здание детской больницы. Затем оно стало аварийным, и больницу выселили; голуби покинули здание сразу вслед за исчезновением людей. Поскольку в том же городе есть немало случаев обитания сизарей в покинутых домах, реакцию трудно интерпретировать иначе как отклик птиц на сигнал «будущей перестройки» данного местообитания (Ангальт, 1989). Опережающие реакции вида на точечные преобразования среды, на устойчивые изменения местной экологической ситуации – норма для всех специализированных городских популяций (Корбут, 2000).
Точность реакции на такие «сигналы» положительно связана со степенью урбанизации вида, но почти не зависит от его синантропности вида. Так, массовый разлёт серых ворон с ночёвок в Москве происходит значительно раньше выключения уличного освещения. Вороны затемно подлетают на «места сбора» вблизи кормовых участков, и в ожидании рассвета стаями сидят на верхушках деревьев и крышах зданий. Кормление начинается уже в утренних сумерках.
Менее урбанизированные галки вылетают на улицу позже, когда становится достаточно светло, хотя синантропность галки выше, чем серой вороны. Даже на Европейском Севере (окрестности Архангельска и других городов) сто лет назад серая ворона гнездилась исключительно в лесу. Но современный уровень урбанизации городских популяций ворон много выше, чем галок, вытесняемых из современного города (Корбут, 2000).
Опережающее реагирование на средовые тренды зафиксируется также на других уровнях популяционной организации в динамике социальной и демографической структуры группировок, в перестройках сети поселений вида конкретной городской территории вслед (или, точнее, перед) долговременной динамикой урболандшафта и пр. Изменяются характеристики отдельных поселений – «узлов» сети популяционных группировок на территории города (плотность населения, межгодовое постоянство и пр.), и характер отношений между группировками, выраженный в интенсивности и направленности обмена особями между группировками, в качественном составе потока переселенцев «туда» и «обратно» (Hanski, 1999).
Важно подчеркнуть, что направленные перестройки популяционной структуры в ответ на сигнал о существенных долговременных изменениях, трендах, опережают «адаптивные уклонения» в биологии и поведении особей, регулируют их встречаемость в городской популяции или выраженность в поведении индивидов, а не наоборот. Например, в начале 1990-х гг. падение численности московской популяции кряквы стимулировало перераспределение зимующих уток внутри города. В период подъёма (1980-е гг.) зимующая популяция крякв Москвы распадалась на две - на внутренних городских водоёмах и на р.Москве, в районе её выхода из города. Автономность каждой группировки подтверждается достижением ими пика численности в разные годы – в 1990 и 1992 гг. Группировка «на городских водоёмах» демонстрировала наибольшие темпы роста в 1980-е гг. (Авилова, 2001).
Падение численности кряквы в начале 1990-х гг. стимулировало быстрое объединение «городских» птиц с «москворецкими». Подъём численности в 1995-1998 гг. вызвал повторное разделение группировок и восстановление зимовок на городских водоёмах. Кряквы г.Москвы размножаются на городских прудах: особи, зимующие там же займут лучшие места гнездования или первыми начнут репродукцию (Авилова, 2001).
Подчеркнём замкнутость городской популяции крякв в Москве и других крупных городах (Минск, Гётеборг, Упсала). Птицы из Подмосковья даже в период дисперсии не проникают в город дальше окраин (Корбут, 1999). Родившиеся в городе молодые частью выселяются за его пределы на расстояние до 30-60 км, но птицы из области почти не проникают в город. Смертность взрослых здесь снижена до 6% в год, а увеличение среднеожидаемой продолжительности жизни (СОПЖ) резидентов позволяет городской популяции расти даже с низким успехом размножения, большом проценте взрослых, не участвующих в репродукции, и ежегодном выселении из города определённой части молодых (Козулин, 1991).
Неустойчивые и уязвимые городские популяции на сигналы – предвестники средовых трендов реагируют прямо противоположным образом. На всех уровнях организации системы (поведение особей, структура группировок, организация всей системы) происходит консервация сложившихся отношений, уменьшение подвижности особей, увеличение «закрытости» поселений, прекращение колонизации даже ближайших подходящих незаселённых участков. Эти регрессивные изменения ведут к сокращению популяции даже при сильной синантропности вида.
Например, популяция крякв г.Минска испытывает явную нехватку особей, способных активно разыскивать и занимать новые места кормления, появляющиеся по всему городскому ареалу. Утки концентрируются на очистных сооружениях и там кормятся в плотных скоплениях, периодически сменяя друг друга на одних и тех же местах. К марту масса тела самок минских крякв ниже, чем у птиц, вернувшихся в область после зимовки. Поэтому размножение крякв в минской популяции в среднем начинается на две недели позже, чем в области, в московской – наоборот (А.В.Козулин, личн.сообщ.).
Деление на альтернативные стратегии наблюдается лишь в тех популяциях, что успешно колонизировали городской ареал и устойчивы в урболандшафтах, исторически нетипичных для вида. Каждый спад численности городской популяции ликвидирует разделение на стратегии, но неблагоприятные изменения среды скорей усиливают его. Так, чёткость деления томской популяции сизаря на «летунов», «собирателей» и «помоечников» увеличивается в холодный период года, в самых напряжённых условиях сбора корма (Ксенц, Москвитин, 1983; Ксенц, 1990).
Поэтому вышеописанные формы популяционной саморегуляции поддерживаются самой надыиндивидуальной системой, а не вызваны суммой независимых реакций отдельных особей на условия среды. Механизмы регуляции здесь те же самые, что поддерживают открытость и проточность видовых группировок, способствуют выделению альтернативных стратегий в соответствии с противоположными полюсами градиента урбосреды. В итоге хорологическая структура городских популяций, кроме дальнего порядка обмена особей между группировками, определяется регуляторной реакцией популяции в целом на частные и локальные изменения среды. Во «внегородских» популяциях тех же видов реакция на местные изменения среды остаётся локальной, не сопровождается приспособительной перестройкой структуры системы на значительно большем пространственном и временном интервале (табл.3).
Синдром дифференцирующих различий в организации «городских» и «внегородских» популяционных систем показан в таблице 3. Альтернативные способы организации отношений между особями в популяциях первого и второго типа выделены на основании восьми оппозиций (I-VIII, табл.3). Противоположность «городского» и «внегородского» типов организации популяционных систем с наибольшей полнотой проявляется в противоположности откликов на изменения среды (флюктуации или тренды) и в характере реагирования особей – элементов системы на успех / неуспех в прохождении разных стадий жизненного цикла. Городские популяции постоянно и с опережением перестраиваются под воздействием сигналов среды, «внегородские» популяции тех же видов остаются консервативными и закрытыми.
На уровне всей популяции способность к опережающему реагированию на сигналы среды проявляется в форме так называемого динамизма (термин С.С.Москвитина, 2001). Под динамизмом понимается способность птиц (большую у одних и меньшую у других видов) «дистанционно оценивать ландшафтную картину больших пространств, в совершенстве ориентироваться, своевременно и направленно реагировать на конкретную климатическую, экологическую и антропогенную ситуацию и высокоэффективно пользоваться ими для себя» (Москвитин, 2001).
Высокий «динамизм» позволяет популяции быстро изменять вид зависимость плотности населения от площади местообитаний с положительной на отрицательную и обратно. При высоком «динамизме» эти изменения связи вида с собственным местообитанием (или, что то же самое, изменения экологической ёмкости местообитания «с точки зрения вида») происходят в ответ на фрагментацию видовых биотопов или нестабильность условий среды. Локальные изменения инициируют быстрые и резкие перераспределения особей по ареалу на значительных территориях внутри ареала, так что «кружево ареала» значительно изменяется (Connor et al., 2000).
С.С.Москвитин (2001) показал, что успех внедрения новых видов в район г.Томска (19 видов за ХХ в., в том числе закрепление в южнотаёжной зоне «южных» и «лесостепных» видов) вызван разными причинами или их сочетаниями, но необходимым условием всегда является высокий динамизм внедренцев. Его же демонстрируют виды-потенциальные урбанисты, когда легко и успешно осваивают городские экологические ниши в разных городах внутри видового ареала. Следовательно, «динамизм» популяционной системы вида причинно связан с присутствием того же типа организации популяционной системы, который задаёт устойчивость городских популяций («городской тип», табл.3). Успешная урбанизация «диких» видов есть крайнее (наиболее специализированное) проявление этого «динамизма».
Анализ расселения видов дендрофильной авифауны к югу по пойменным лесам таких рек, как Урал, Дон, Северской Донец, по лесополосам, посаженным в рамках Сталинского плана преобразования природы, вполне подтверждает эту гипотезу. Виды, наиболее склонные к урбанизации, показывают и наибольший динамизм в освоении новосозданных «островных ландшафтов», в расселении по ним к югу, в нелесные пространства степи и даже полупустыни. Это большая синица, зяблик (Fringilla coelebs), лазоревка (Parus caeruleus), мухоловка-пеструшка, чёрный дрозд, зарянка, большой и малый пёстрые дятлы, ушастая сова (Asio otus), вяхирь и пр. – всё потенциальные урбанисты (Белик, 1997).
Их скорость продвижения к югу по лесным «островам» и «полосам» в пойме р.Урал достигает 2-6 км/год км в год. Минимальная скорость расселения зафиксирована у самых крупных из названных видов, вяхиря и большого пёстрого дятла (2-4 км/год). Во второй половине ХХ в. резко возросла скорость расселения всех видов, лидирующих по темпам урбанизации: у большой синицы с 2 до 14 км/год, у лазоревки и зяблика с 2-3 до 5-6 км/год. С той же скоростью неморальные и бореальные виды птиц расселялись в ХХ веке в низовьях Северского Донца (Белик, 1997). Большинство городских популяций этих видов в урбанизированных регионах Европейской части СССР возникали в годы «повышенного динамизма»=наибольшей скорости расселения дендрофильных видов в нелесные зоны.
Показательно, что скорость распространения урбанизационных процессов у таких видов, как чёрный дрозд, вяхирь, рябинник или тетеревятник на 1-2 порядка превышает самые высокие темпы расселения, зафиксированные при расширении ареала вида в природных ландшафтах (то же верно для скоростей расселения по лесополосам и лесным микрофрагментам в нелесном ландшафте). Например, скорости, зарегистрированные при расселении овсянки-дубровника (Emberiza aureola), чечевицы (Carpodacus erythrinus), зелёной пеночки (Phylloscopus trochyloides), синехвостки (Tarsiger cyanurus) в ХХ веке. Аналогичные расширения ареалов здесь требуют десятилетий (Зимин, 2001).
В целом, виды-потенциальные урбанисты за годы по антропогенно-фрагментированным местообитаниям расселяются на расстояние, которое другие виды в непрерывных местообитаниях «проходят» за десятилетия. Поэтому они были названы В.В.Леоновичем (1991) «виды-выскочки». Это значит, что потенциальным урбанистам исходно присущ иной тип организации популяционной системы, наиболее приближающийся к «городским» вариантам организации. Благодаря ему именно в «островном ландшафте», созданном и поддерживаемом человеческой деятельностью (неважно, в городе или за его пределами), все эти виды обретают высокую устойчивость популяций и особый динамизм. Им исходно присущ «городской» тип популяционной организации или близкий к нему (табл.3), тогда как прочие виды региональной авифауны лабилизируют популяционную структуру – и жизненные стратегии индивидов – лишь попав «под давление» урболандшафта.
В силу этого виды-выскочки характеризуются лабильной жизненной стратегией даже в негородских местообитаниях и первыми колонизируют урболандшафт. Все остальные виды формируют «городскую» организацию популяционных систем лишь в ответ на трансформацию исходных местообитаний процессом региональной урбанизации, ростом «городского» ядра, причём ответ всегда вынужденный, и в большей или меньшей степени запаздывающий, тогда как у видов-выскочек он носит опережающий характер (Фридман и др., 2008).
- Факторы жизнеспособности городских популяций: как система сохраняет устойчивость в условиях постоянных средовых изменений?
Устойчивость популяционных систем экологи определяют по-разному, в зависимости от того, какой из аспектов этого явления вносит наибольший вклад в жизнеспособность системы (табл.4). Устойчивость популяционной системы проявляется как её «эластичность» (resilience) или «стойкость» (persistence). Первая связана со способностью восстанавливать определённую организацию системы в условиях постоянных флюктуаций среды, и адекватно перестраивать популяционную структуру ради сохранения неких инвариантов организации в ответ на направленные изменения среды (тренды). Вторая - со стабильностью определённого «типичного» паттерна структуры в условиях, когда можно пренебречь его «нарушением» под действием возмущений среды. «Эластичность» сохраняет устойчивость системы в ситуациях, когда нарушением нельзя пренебречь (Holling, 1969; Розенберг и др., 1999).
Очевидно, устойчивость городских популяций связана скорее с «эластичностью», чем со «стойкостью» системы, и тем в большей степени, чем дальше зашла урбанизации вида. «Эластичность» проявляется как способность популяции быстро восстанавливать структуру группировок при постоянных нарушениях местообитаний, постоянно перестраивать сеть поселений вида, чтобы обнаружить и использовать все вновь появившиеся «островки» подходящих местообитаний, сохранить устойчивость всей сети при продолжающейся фрагментации последних. Процессуальным механизмом этих перестроек является «проточность» группировок, «дальний порядок» обмена особями в городсой популяции и сортировка особей в ответ на сигналы среды.
Необходимость «сортировочного» механизма устойчивости городских популяций показывает математическая модель, заимствованная из работы Г.С.Розенберга (2000).
Рассмотрим надёжность статистической системы А (городской авифауны или отдельных видовых популяций). В момент времени она состоит из элементов еi (i=1, n())-видов. В результате взаимодействия с некими факторами среды каждый вид-элемент системы «фауна» (или особь с определённой стратегией - элемент системы «популяция») независимо от других «погибает» - исчезает из фауны с постоянной вероятностью рi Система А может пополниться любым числом элементов или воспроизвести их, причём внешняя среда «не мешает» этому (индифферентна по отношению к системе).
Если цель системы R - выжить, т.е. просуществовать как можно дольше, можно считать, что АR (система А с целью R) «приобретает» у среды время жизни , «оплачивая» его гибелью своих членов n. Для особей в составе популяционной системы «гибель» означает выселение особей за пределы урболандшафта, для видов локальной фауны – прекращение гнездования на отдельном «острове» или на всей территории города.
Тогда можно говорить о существовании обмена (n;) и решать задачу об определении потенциально-эффективного (n;0)-обмена: до какого минимального числа элементов n() должна пополняться система АR, компенсируя элементы, погибшие в момент {-1}, чтобы с вероятностью сколь угодно близкой к единице существовать длительное время 0 .
Потенциально эффективный (n;)-обмен имеет место при 0 = (n) = exp [CR*n(t)], где СR=CR({pi}) - фундаментальная константа, зависящая от вероятности гибели элементов системы А. В нашем анализе СR определяется вероятностью выпадения вида из городской авифауны и вероятностью включения новых видов в состав городской авифауны. Фактически это показатель того уровня стрессирующих воздействий среды на систему, который должен быть преодолён за счёт трансформации популяционной структуры вида, чтобы обеспечить ему устойчивость существования в «архипелаге» городских биотопов.
Та же модель применима к обоснованию устойчивости конкретных фаун – орнитокомплексов, складывающихся на «островах» природных территорий. Аналогом устойчивости популяционной системы будет постоянство ежегодного гнездования N разных видов, составляющих фауну данного «острова» или «архипелага».
Анализ Г.С.Розенберга (2000) показывает, что неограниченно долгое существование городской популяции (или постоянное присутствие N видов в «архипелаге» местообитаний, когда .>0) можно обеспечить только постоянным ростом популяционной устойчивости и, в меру последнего, - направленной колонизацией видом всё более урбанизированных районов города, где нестабильность среды особенно велика. Или (переходя от популяционной устойчивости к устойчивому существованию разных видов на городской территории) успешным освоением города новыми и новыми видами региональной фауны за время , и их включением в существующие орнитокомплексы.
Поэтому надёжность системы «популяция» прямо зависит от темпов колонизации более урбанизированных районов города, системы «фауна» - от темпов урбанизации «диких» видов. Обе зависимости усиливаются пропорционально величине pi (чувствительность элементов системы А к средовому стрессу), меняющейся на градиенте от периферии к центру города.
Условие жизнеспособности популяции на урбанизированной территории и в других типах ландшафта с высокой вероятностью «выпадения» элементов системы аналогично. Необходимы
1) постоянный подток особей из периферийных районов города с меньшими значениями pi, то есть с меньшим уровнем средового стресса, включая подток птиц из пригородов,
2) обратный отток на участки с меньшим pi особей, «выпадающих» из состава группировок «более высоких» участков городского градиента в силу «недостаточно урбанистичной» стратегии,
3) немедленное включение особей, переселяющихся в процессе сортировки, в состав поселений «принимающей» территории, эффективное участие «пришлых» особей в размножении и занятии участков (не менее эффективное, чем у «местных»),
4) лабильность организации популяционной системы вида на территории города должна расти пропорционально степени урбанизации, но быстрее, чем сам городской градиент. В этом случае «конструкция» популяционной системы будет достаточно лабильна, чтобы обеспечить эффективность и точность двусторонней сортировки особей в городской популяции, но недостаточной, чтобы исказить видовой «тип» социальных, территориально-пространственных и биотопических отношений между особями.
Очевидно, городская популяция устойчива и будет расти, если «выигрыш» от заселения новых территорий и успешного гнездования на вновь занятых «островах» превышает «потери» от уничтожения части местообитаний, уменьшения репродуктивного выхода популяции под действием «экологических ловушек» и пр. Для успешного освоения «диким» видом урбосреды необходимо, чтобы на городской территории баланс «выигрышей» и «плат» урбанизации был устойчиво положительным на протяжении тех 20…30 лет, необходимых для необратимости процесса (Фридман и др., 2005, 2008).
Очевидно, что баланс «плат» и «выигрышей» воспроизводства городской популяции положителен тем в большей степени, чем большее развитие получил «городской» тип организации популяционной систем, обеспечивающий устойчивость существования вида при наиболее высоком уровне средового стресса (табл.3, рис.2). Присущая ему "конструкция" популяционной системы достаточно лабильна, чтобы обеспечить эффективность и точность двусторонней сортировки особей в городской популяции, но недостаточной, чтобы исказить видовой "тип" социальных, территориально-пространственных и биотопических отношений между особями.
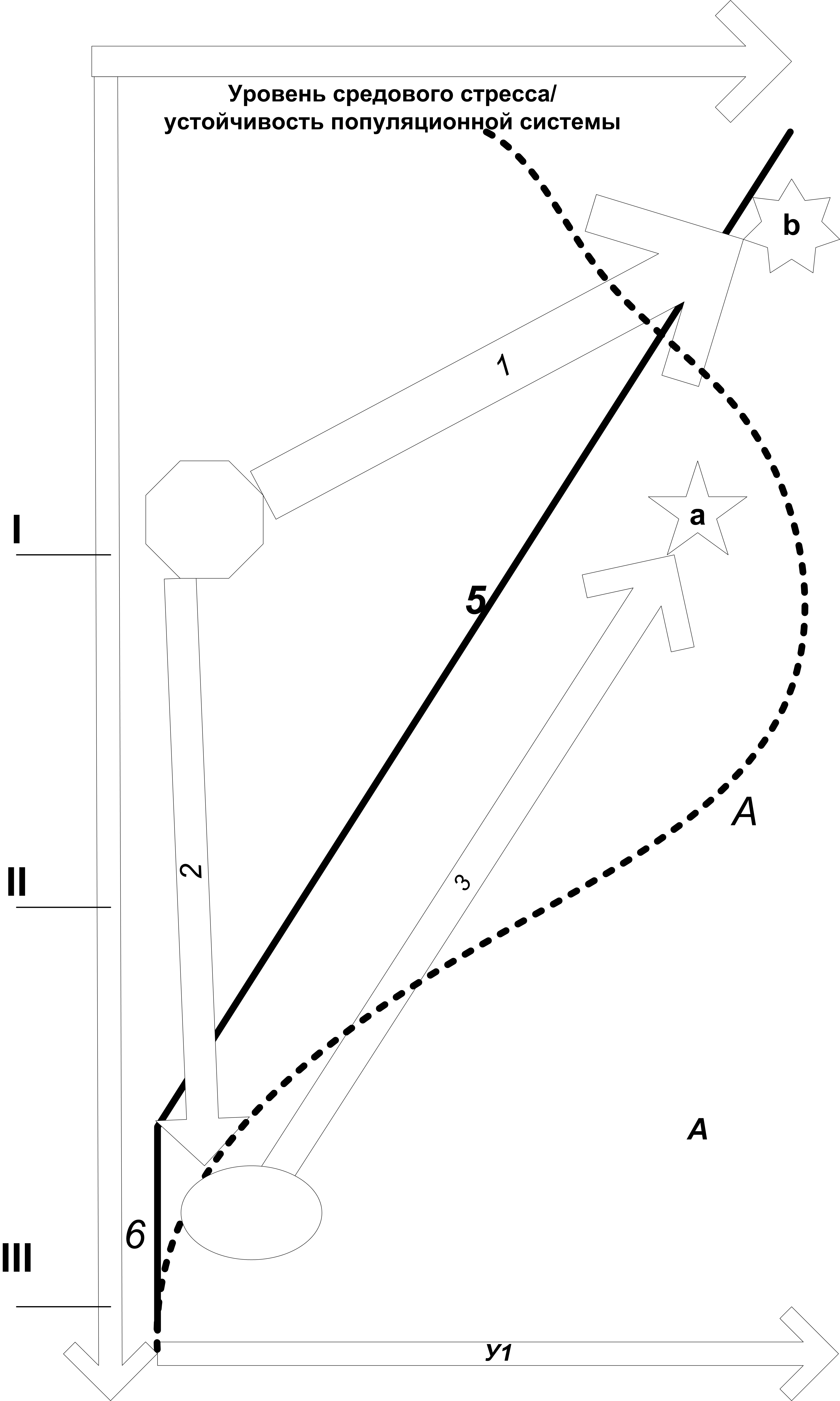
Рисунок 2. Динамика стрессирующих воздействий городской среды в процессе урбанизации территории, рост устойчивости городской популяции в процессе урбанизации вида. 1-3 – разные варианты взаимодействия популяционных систем с растущим урбанизированным ареалом: 1 – немедленная урбанизация (урбофилы), 2 – необратимое вытеснение из города и зоны влияния урбанизированного «ядра» (урбофобы), 3 – «возвратная урбанизация» после определённого периода существования в зоне влияния «ядра» (виды, переходящие от вытеснения к урбанизации через трансформацию популяционной системы вида в зоне влияния города и в самом городе). Абсцисса – городской градиент (I-III рис.1), ордината – уровень воздействия средового стресса на популяцию, если речь идёт о самой урбосреде, и устойчивость воспроизводящейся в ней популяционной системы, когда речь идёт о видах, осваивающих урбанизированный ареал.
Примечание. Уровень средового стресса определяется общей «плотностью нарушений» местообитаний вида вдоль городского градиента (суммируется из воздействий 1-4 рис.1). Σ – изменения «плотности нарушений» местообитаний вида вдоль городского градиента на фоне таких же изменений плотности городского населения (А, Лаппо, 1997).
Когда устойчивость популяционной системы ниже уровня стрессирующих воздействий среды, вид вытесняется на «более низкие» участки городского градиента, характеризующиеся меньшим воздействием средового стресса (процесс вытеснения 2). Когда устойчивость популяций растёт в результате эндогенной трансформации структуры системы, вид осваивает всё более «высокие» участки городского градиента, и в конце концов заселяет весь город (процессы урбанизации 1, 3).
Figure 2. Dynamic of environmental stress level during urbanization the territories, growth of the steadiness in the population systems during urbanization this species ( 1-3 – different schemes of interactions between population systems and growing urbanized areas). 1 – immediate urbanization (urbophilic bird species), 2 – irreversible exclusion from urbanized area and their impact zone (undoubted urbophobic species), 3 – «recurrent urbanization» after certain period existing this species in city impact zone (species transition from exclusion to urbanization with help the transformation of population system). X axis – urban gradient (I-III fig.1). Y axis – environmental stress level as the question is about urban environment yourself, and the steadiness of population systems as the question is about different bird species, which begin to settle new urbanized area. А – changes of citizens population density along of urban gradient (Lappo, 1997), Σ – changes of «distribution density» all habitat disturbance along urban gradient in cities.
Comment. If the steadiness of species population system is lower as the environmental stress level, the species will excluded on the «more lower» parts of urban gradient (exclusion process 2). When the population steadiness increase as a result of transition to «sustainable variants» of system structure, the species can to occupy all «more higher» parts of urban gradient (urbanization processes 1 and 3).
Действительно, анализ различий в демографической структуре городских и внегородских популяций (табл.5) говорит о том, что при образовании первых из вторых происходит резкая лабилизация структуры системы, пространственно-этологической, биотопической, территориальной и пр. Она тем больше, чем выше степень урбанизации территорий, колонизируемых видом внутри города, и при успешной урбанизации вида первое связано положительной обратной связью со вторым. Благодаря лабилизации социодемографических отношений в системе (при сохранении некого видового "типа") индивид в городской популяции обретает свободу выбора той экологической и социальной среды в мозаике урболандшафта, которая наиболее соответствует его жизненной стратегии.
Таблица 5. Основные демографические изменения в городских популяциях разных видов по сравнению с «внегородскими».
Table 5. Basic changes the demographic structure of urban bird population comparing with population of same species out of urban landscape boundaries
| Общие демографические изменения в городских популяциях Some demographical parameters changing in urbanized population the birds and mammals | Вид, источник Species and source |
| Увеличение среднеожидаемой продолжительности жизни резидентов и птиц из «резерва» | A.platyrhynchos - Козулин, 1991; T.merula – Luniak, 2004; P.major – Гашков, Москвитин, 2001; S.vulgaris, F.hypoleuca – Куранов и др., 1991; Куранов, 2005; C.brahyrhynchos – Marzluff et al., 2001а; C.livia – Ксенц, 1990; Обухова, 1999. |
| Сокращение ежегодного успеха размножения индивидов (падение среднего числа слётков на успешно размножавшуюся пару в городе по сравнению с пригородом) | A.platyrhynchos - Козулин, 1991; T.merula – Luniak, 2004; P.major – Гашков, Москвитин, 2001; Куранов и др., 1991; S.vulgaris, F.hypoleuca – Куранов и др., 1991; Куранов, 2005; C.brahyrhynchos – Marzluff et al., 2001а. |
| Повышенный риск необразования пары и отказа резидентов от размножения в городских популяциях | A.platyrhynchos – Корбут, 1999; С.cornix – Baglioni et al., 2005; S.vulgaris, F.hypoleuca, P.major – Куранов и др., 1991; Куранов, 2005 |
| Рост успешности повторной кладки на той же территории после первого разорения гнезда | Дрозды р. Turdus – Черенков и др., 1995; Морозов, 2000; S.vulgaris, F.hypoleuca, P.major – Куранов и др., 1991; Куранов, 2005; C.brahyrhynchos – Marzluff et al., 2001а |
| Увеличение среднего размера кладки в городской популяции (эффект умеренной фрагментации местообитаний) | F.hypoleuca – Huhta et al., 1994; Куранов, 2005; S.vulgaris, Ph.phoenicurus, P.major – Куранов и др., 1991 |
| Повышенный риск гибели гнезда и птенцов от неспециализированных хищников во фрагментированных местообитаниях, особенно при размещении гнезда в «наиболее видоспецифических» микростациях | Искусственные гнёзда - Jokimäki, Huhta, 2000; S.vulgaris, Ph.phoenicurus, P.major – Куранов и др., 1991; F.coelebs, T.philomelos, T.merula, T.pilaris, L.collurio, S.borin, S.communis, S.nisoria, A.trivialis, E.citrinella – Сарычев, 2001; Черенков и др., 1995; Морозов, 2000. |
| Возрастание средней дальности обмена особями между поселениями до размеров самого города | C.livia – Ксенц, 1990; P.major – Гашков, Москвитин, 2001; Юдкин, Юдкин, 2002; С.cornix – Константинов и др., 1981; Baglioni et al., 1998, 2005. |
| Рост доли птиц «подвижного резерва» популяции, в том числе за счёт резидентных особей | A.platyrhynchos – Корбут, 1999; С.cornix – Baglioni et al., 1998; 2005; S.vulgaris, F.hypoleuca, P.major – Куранов и др., 1991; Куранов, 2005 |
| Разделение популяции на группы «сверхподвижных» и «сверхоседлых» особей. Птицы или исключительно привязаны к собственной территории, или легко переселяются в дальние, но не в соседние группировки. | A.platyrhynchos - Корбут, 1999; P.domesticus, P.montanus – Иваницкий, Матюхин, 1990, 1995; С.cornix – Корбут, 2001; Baglioni et al., 2005; P.major – Москвитин, Гашков, 2000; Гашков, Москвитин, 2001; T.merula – Møller, 1995; Luniak, 2004; C.livia – Ксенц, Москвитин, 1983; Ксенц и др., 1985, 1987; Ксенц, 1990. |
| Направленная сортировка особей в городской популяции: птицы с более урбофильной стратегией расселяются «вверх» по городскому градиенту, с урбофобной – отселяются «вниз» | P.major – Юдкин, Юдкин, 2002; Гашков, Москвитин, 2000; C.livia – Ангальт, 1989; Ксенц, 1990; T.merula – Møller, 1995; Luniak, 2004; |
| Направленный обмен особей между городской популяцией и населением ближних пригородов: особи с более урбофильной стратегией включаются в состав городских поселений, наиболее урбофобные из городских птиц выселяются в пригородные биотопы. | A.platyrhynchos – Козулин, 1991; Корбут, 1999; T.merula - Møller, 1995; P.major – Грищенко, 1995; Москвитин, Гашков, 2000; Юдкин, Юдкин, 2002; С.cornix – Константинов и др., 1981; C.livia – Ангальт, 1989; Ксенц, 1990; |