
Влияние возраста и физиологического состояния животных на активность ферментных систем клеток, тканей и органов 03. 03. 01 физиология 03. 01. 04 биохимия
| Вид материала | Автореферат |
- «Физиология и этология животных» Специальность 111201. 65 Ветеринария. Пояснительная, 240.53kb.
- Методические рекомендации к написанию и оформлению рефератов по курсу «физиология, 129.81kb.
- Анатомия и физиология человека Общее знакомство с организмом человека, 174.34kb.
- Задачами физиологии и этологии животных являются, 536.54kb.
- Тема: Двигательная активность и лфк для людей пожилого возраста, 353.11kb.
- «Физиология нервной системы», 172.86kb.
- Пояснительная записка Нормальная физиология наука и учебная дисциплина о жизненных, 578.98kb.
- Влияние пренатального стресса на процессы окислительной модификации белков и активность, 202.57kb.
- Рабочая программа дисциплины «физиология» (Физиология животных), 288kb.
- Программа дисциплины «Физиология с основами анатомии», 721.8kb.
«*» - р0,05 по сравнению с аналогичными показателями в стадии уравновешивания полового цикла
«» - р0,05 по сравнению с аналогичными показателями 1-й трети рогов матки
«+» - р0,05 по сравнению с активностью Mg2+-АТФазы
Таблица 13. Коэффициенты корреляции (rxy) активности Mg2+-АТФазы
с активностями Na+,K+-; Ca2+-; HCO-3-АТФаз
АТФазы | Внутриклеточные органоиды | ||
| Ядра | Митохондрии | Лизосомы | |
| Na+,K+- | 0,92** | 0,02 | 0,96** |
| Ca2+- | 0,94** | 0,98** | 0,99** |
| HCO-3- | 0,85** | -0,25 | 0,87** |
Таблица 14. Коэффициенты корреляции (rxy) активности Na+,K+-АТФазы с активностями; Ca2+-; HCO-3-АТФаз и корреляции активностей Ca2+-АТФазы и HCO-3-АТФазы
АТФазы | Внутриклеточные органоиды | ||
Ядра | Митохондрии | Лизосомы | |
| Ca2+- | 0,73** | -0,19 | 0,99** |
| HCO-3- | 0,99** | 0,96** | 0,97** |
| Ca2+- к HCO-3- | 0,62 | -0,45 | 0,94** |
Активность АТФаз при остром серозно-катаральном эндометрите
Активность ядер клеток эндометрия. При остром эндометрите отмечается достоверное (р0,05) снижение активности всех изучаемых АТФаз ядер клеток эндометрия во всех участках рогов матки по сравнению с аналогичными показателями у свиноматок без патологии. Так, в 1-й трети рогов матки при остром эндометрите наиболее выраженное снижение ферментативной активности отмечалось у Са2+-АТФазы, уровень активности которой снижался на 29% по сравнению с аналогичным показателем у здоровых свиноматок, а наименьшее снижение активности установлено у HCO3-АТФазы 20%. Во 2-й трети рогов матки при воспалительном процессе регистрировалось снижение ферментативной активности Mg2+- АТФазы (33%). В 3-й трети рогов активность Na+, K+-АТФазы, была ниже аналогичного показателя у здоровых свиноматок на 29%. При этом активность HCO3-АТФазы в этом участке была на 20% ниже, чем у контрольных животных.
Активность митохондрий клеток эндометрия. При остром эндометрите происходит достоверное снижение активности АТФаз митохондриальной фракции клеток эндометрия свиноматок во всех участках рога матки по сравнению с аналогичными показателями у свиноматок без патологии.
Так в 1-й трети рогов матки наименьшее снижение ферментативной активности было отмечено у Са2+-АТФазы (21%), во 2-й трети у Na+, K+-АТФазы (17%) и в 3-й трети у Mg2+-АТФазы (13%), а наибольшее снижение активности в 1-й трети у Mg2+-; Na+, K+- и HCO3--АТФаз (26-27%), во 2-й трети рогов для HCO3--АТФазы – 29%, и в 3-й трети – для Na+, K+- и HCO3--АТФаз 28% и 27% соответственно по сравнению с аналогичными показателями у здоровых свиноматок.
Активность лизосом клеток эндометрия. При остром эндометрите отмечается достоверное снижение активности всех изучаемых АТФаз лизосом во всех участках рогов матки по сравнению с соответствующими показателями у свиноматок без патологии. Так, активность HCO3-АТФазы составляла 76,60,9, 92,61,0 и 55,51,2 нмоль Рнмг белка-1мин-1, что соответственно ниже на 36%, 39% и 43% по сравнению с этими показателями у здоровых свиноматок. Наибольшее снижения ферментативной активности установлено для Мg2+-АТФазы лизосом во всех участках рогов матки на 62-71% по сравнению с аналогичными показателями у здоровых животных.
Активность АТФаз при хроническом эндометрите.
Активность АТФаз ядер клеток эндометрия. В 1-й трети рогов матки при хроническом воспалении активность Mg2+-; Na+, K+- и Са2+-АТФаз существенно не изменялась по сравнению с аналогичными показателями у здоровых животных, за исключением HCO3- АТФазы, уровень активности которой снижался на 13%. Во 2-й трети рогов матки активность Mg2+-; Na+, K+-и HCO3- АТФаз была ниже по сравнению с аналогичными показателями у свиноматок без патологии соответственно на 4% 17% и 28%, а активность Са2+-АТФазы, напротив, была выше на 7%. В 3-й трети рогов матки отмечено достоверное увеличения активности Mg2+- и Са2+-АТФаз соответственно на 10% и 12% и уменьшения Na+, K+-и HCO3--АТФаз на 6% и 7% по сравнению с аналогичными показателями у здоровых свиноматок.
Активность АТФаз митохондрий клеток эндометрия. Активность Mg2+-; Na+, K+-и HCO3--АТФаз митохондрий клеток эндометрия свиноматок при хроническом эндометрите в 1-й и 2-й третях рогов матки была достоверно выше по сравнению с соответствующими показателями у здоровых животных. Так, в 1-й трети рогов матки ферментативная активность Mg2+-АТФазы была выше на 7%, Na+, K+- на 10%, Ca2+- на 6% и HCO3АТФазы на 7% по сравнению с этими показателями у здоровых животных, а во 2-й трети - активность Mg2+-; Na+, K+-; Ca2+- и HCO3--АТФаз была выше соответственно на 6%, 16%, 12% и 4%.
В 3-й трети рогов матки активность Na+, K+-; Ca2+- и HCO3-АТФаз наоборот, была ниже, чем у здоровых животных на 4%, 2% и 2,5%, а активность Mg2+-АТФазы на 9% выше.
Активность АТФаз лизосом клеток эндометрия. В лизосомах клеток эндометрия свиноматок при хроническом эндометрите в 1-й трети рогов матки отмечается достоверное повышения активности Mg2+-; Na+, K+-; Ca2+- и HCO3--АТФаз, соответственно на 21%, 12%, 27% и 14%, по сравнению с аналогичными показателями у здоровых животных. Во 2-й трети рогов матки активность Mg2+-; Na+, K+- и HCO3--АТФаз была выше на 15%, 64% и 22%, чем у здоровых животных, а активность Ca2+-АТФазы не имела достоверных отличий. В 3-й трети рогов матки активность Mg2+-; Na+, K+-; Ca2+- и HCO3--АТФаз была на 34%,37%, 32% и 42% выше по сравнению с активностью АТФаз лизосом эндометрии свиноматок без патологии.
Активность ферментов эндометрия.
Активность протеазВлияние рН среды на активность протеолитических ферментов в эндометрии свиноматок. Наибольшая протеолитическая активность (рис. 10) наблюдалась при рН 4,5 и составляла 55,76 мкмоль тирозинамг белка-130мин-1.
В интервале рН от 7 до 10 отмечалось несколько пиков протеолитической активности, при рН 7,41, рН 8,0, рН 8,75 и рН 10,30, что говорит о том, что в эндометрии присутствует сложный набор этих ферментов, имеющих различное происхождение и биологическую роль.
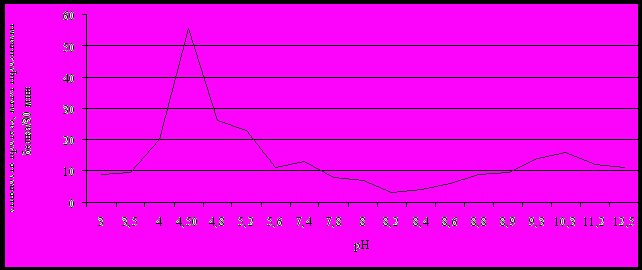
Рис. 10. Активность протеолитических ферментов эндометрия свиноматок в зависимости от рН среды инкубации.
Влияние стадий полового цикла и локализации в роге матки на активность ферментов. Исследованиями выявлены определенные закономерные, зависимые от стадии полового цикла и локализации в матке изменения ферментного профиля эндометрия (табл.15). Максимальная активность протеаз регистрировалась во время стадии торможения полового цикла, а минимальная - во время стадии уравновешивания. Вместе с тем наибольшая активность кислых протеаз независимо от стадии полового цикла отмечалась во 2-й трети рогов матки, что связано с развитой сетью артерий и большим количеством маточных желез в этой части эндометрия.
Наибольшая активность кислых фосфатаз была зарегистрирована в стадии торможения. В зависимости от места локализации высокий уровень общей и простатической кислых фосфатаз наблюдался во 2-й трети рогов матки во все стадии полового цикла.
Высокая активность щелочной фосфатазы в эндометрии и маточной слизи регистрировалась в стадии торможения, а минимальная - уравновешивания.
Активность АСТ и АЛТ имела наибольший уровень в стадии возбуждения полового цикла, что связано с усилением процессов пролиферации в эту стадию. Независимо от стадии полового цикла высокая активность АСТ и АЛТ отмечалась во 2-й трети рогов матки, что также связано с развитой сетью артерий и маточных желез.
Таблица 15. Активность ферментов эндометрия, и маточной слизи в зависимости от стадии полового цикла и локализации в матке, (n=9)
| Стадия | 1-я треть | 2-я треть | 3-я треть | Маточная слизь |
| Катепсины, мкмоль тирозина ·мг белка-1·мин-1 | ||||
| Возбуждения | 2,860,04 | 3,610,08* | 2,900,08 | 3,240,08* |
| Торможения | 3,140,04 | 4,200,09* | 3,430,05 | 3,290,08 |
| Уравновешивания | 1,610,07 | 2,140,04* | 2,090,04* | 4,380,06* |
| Общая кислая фосфатаза, ммоль·мг белка-1*мин-1 | ||||
| Возбуждения | 13,120,41 | 17,300,69* | 18,340,71* | 15,620,98 |
| Торможения | 20,600,90 | 22,200,81 | 18,410,81 | 17,471,21 |
| Уравновешивания | 13,240,61 | 22,460,80* | 20,260,86* | 21,580,89* |
| Простатическая кислая фосфатаза, ммоль·мг белка-1·мин-1 | ||||
| Возбуждения | 5,020,25 | 4,060,21 | 5,840,11 | 4,720,34 |
| Торможения | 10,62022 | 13,300,36 | 6,920,22** | 5,550,27 |
| Уравновешивания | 5,480,15 | 6,890,23 | 7,100,24* | 7,020,21 |
| Щелочная фосфатаза, ммоль·мг белка-1·мин-1 | ||||
| Возбуждения | 25,381,40 | 15,200,98* | 22,301,59 | 9,500,17* |
| Торможения | 42,603,09 | 49,883,38 | 41,723,52 | 24,41,14 |
| Уравновешивания | 11,040,46 | 9,700,25 | 9,570,18 | 12,700,52 |
«*» - р0,05 по сравнению с аналогичным показателем, полученными при анализе материала из 1-й трети рога матки;
«» - р0,05 по сравнению с аналогичными показателями, полученными в стадию уравновешивания полового цикла.
Активность ферментов при остром эндометрите. При остром эндометрите у свиноматок достоверно (р0,05) повышается активность катепсинов эндометрия и маточной слизи. Наиболее выраженное повышение этого показателя отмечалось во 2-й трети рогов матки, где он составлял 5,900,17 мкмоль тирозина·мг белка-1·мин-1. Наименьший прирост активности кислых протез при остром эндометрите был отмечен в 1-ой трети рога матки 2,300,08 мкмоль тирозина·мг белка-1·мин-1.
Активность общей и простатической кислой фосфатаз при остром эндометрите достоверно (р0,05) повышается во всех участках рогов матки по сравнению с аналогичным показателем у здоровых животных. Так, отмечено достоверное увеличение активности этого фермента в 1-ой трети рогов матки в 9 раз, во 2-ой – в 11 раз и в 3-ей – в 6 раз. Активность простатической формы кислой фосфатазы при эндометрите достоверно увеличилась в 1-ой трети рогов матки в 8,4 раза, во 2-ой – в 15 раз и в 3-ей – в 5,8 раза.
При остром эндометрите отмечается достоверное снижение активности щелочной фосфатазы во всех участках рогов матки и в маточной слизи. При этом активность ЩФ составляла в 1-ой трети рогов матки 2,5±0,1, во 2-ой трети 3,04±0,04 и в 3-ей трети 2,3±0,1 ммоль·мг белка-1·мин-1, что соответственно в 4,5, 3 и 4 раза ниже аналогичных показателей у здоровых животных. В данном случае снижение активности ЩФ вероятно связано с нарушением трансмембранного переноса глюкозы и других сахаров в результате резкого уменьшения энергетического обмена и активности транспортных систем при развитии острого воспаления.
Активность АЛТ в эндометрии и маточной слизи при остром эндометрите достоверно увеличивается по сравнению со здоровыми животными. Отмечено, что активность этого фермента повышалась в 1-ой трети в 1,6 раза, во 2-ой трети в 1,4 раза и в 3-ей трети в 1,6 раза по сравнению со здоровыми свиноматками. Активность АЛТ в маточной слизи больных эндометритом животных также была достоверно выше аналогичного показателя у здоровых животных в 2,1 раза. Такое увеличение активности АЛТ, по-видимому, связано с механизмами обезвреживания аммиака, образующегося в очаге воспаления при острой гипоксии и усиленном распаде аминокислот.
Достоверных отличий между показателями активности АСТ при остром эндометрите и у здоровых животных в 1-ой трети рогов матки выявлено не было. В то же время во 2-ой и 3-ей третях рогов матки наблюдалось снижение активности.
В маточной слизи повышение активности АСТ (4,74±0,20 ммоль·мг белка-1·мин-1) было незначительным.
Локализации кислой и щелочной фосфатаз в эндометрии свиноматок в зависимости от стадии полового цикла и развития воспалительного процесса.
Кислая фосфатаза. В стадии возбуждения полового цикла в поверхностном эпителии отмечена диффузная, мелкозернистая, умеренная реакция на КФ - в основном за счет наличия интенсивно окрашенных ядрышковых структур диаметром приблизительно 2-3 мкм, окруженных светлым ореолом нуклеоплазмы и содержащихся по 2-3 в одной клетке. В железистом слое – интенсивная реакция, особенно в базальной и средней части желез, за счет большой концентрации окрашенных ядрышковых структур.
В период стадии торможения полового цикла отмечается слабая реакция в поверхностном эпителии. В железах – умеренная реакция в апикальной части. В базальной части – интенсивная реакция ядрышковых структур.
В стадии торможения отмечена умеренная мелкозернистая реакция на кислую фосфатазу в цитоплазме и ядрах поверхностного эпителия и интенсивная реакция в многочисленных тучных клетках эндометрия.
При развитии воспаления в эндометрии установлено повышение активности КФ в поверхностном эпителии, причем реакция носит неравномерный характер с выявлением активности, не только в ядрах, но и в цитоплазме. Отмечена интенсивная реакция в тучных клетках и железах, в основном диффузного характера.
Щелочная фосфатаза. В стадии возбуждения полового цикла выявлена умеренная реакция в апикальной каемке поверхностного эпителия и слабая – в компактном (подэпителиальном) слое. По контуру желез и в адвентиции сосудов отмечена умеренная реакция. Интенсивная активность ЩФ проявляется в цитоплазме лейкоцитов и в просвете крупных кровеносных сосудов.
В стадии торможения полового цикла отмечена неравномерная диффузная активность ЩФ в поверхностном эпителии и практически полное отсутствие реакции в железах, стромальных и сосудистых структурах.
В стадии уравновешивания наблюдалось снижение активности ЩФ – диффузно-очаговая умеренная реакция в поверхностном эпителии. В эпителии желез выявлена умеренная очаговая диффузная мелкозернистая реакция в виде хлопьевидных масс.
При развитии воспаления наблюдалось неравномерное истончение поверхностного эпителия с диффузно-очаговой активностью ЩФ, не имеющей апикально - базальной зональности цитоплазмы. В железах и капиллярах отмечена умеренная мелкозернистая реакция.
Таким образом, активность и локализация фосфатаз эндометрия меняется в зависимости от стадий полового цикла. Так, если в стадии возбуждения основная активность КФ выявлена в ядрах поверхностного эпителия и желез, то в стадии уравновешивания активность, наблюдалась, в основном только в тучных клетках. Активность ЩФ, наоборот, локализована в апикальной каемке покровного эпителия и по контуру желез, а при переходе от стадии возбуждения к стадии торможения и уравновешивания она перераспределялась по всей цитоплазме, а в железах практически не выявлялась.
Во время развития воспаления активность КФ эндометрия существенно повышается, и в поверхностном эпителии наблюдается ее неравномерность за счет снижения рН цитоплазмы в условиях гипоксии в очаге воспаления, создающего оптимальные условия для функционирования этого фермента и процессов цитолиза. Активность ЩФ при развитии воспаления незначительно повышается, особенно в железах, что, по-видимому, связано с усиленными процессами экссудации и инфильтрации.
ВЫВОДЫ
1. Активность ферментных систем клеток, тканей и органов зависит от возраста и вида животных:
1.1. Активность Na+, K+-АТФазы цитоплазматических мембран эритроцитов цыплят на 54,8% зависела от возраста, активность Ca2+-АТФазы – на 72,5% и HCO3--АТФазы – на 36,1%. Активность ядерных АТФаз была детерминирована возрастом на 91,9% - Na+, K+-АТФазы, 92,3% - Ca2+-АТФазы и HCO3--АТФазы – на 28,3%. Это говорит о существенных перестройках АТФазных ферментных систем ядер и цитоплазматических мембран эритроцитов цыплят с возрастом.
1.2. Активность Mg2+,Na+,K+-, Mg2+- и Na+,K+-АТФаз зависела от возраста свиней. Динамику возрастного снижения активности ферментов, на основании регрессионного анализа, можно расположить в порядке убывания следующим образом: Mg2+-АТФазаMg2+,Na+,K+-АТФаза Na+,K+-АТФаза. Дисперсионный анализ показал, что активность АТФазы была детерминирована на 9,15% возрастом и на 87,34% строфантином-G (Р<0,05). Совместное влияние факторов было незначительным (0,58%).
2. В ядерных и цитоплазматических мембранах эритроцитов цыплят-бройлеров выявлены активности АТФазных ферментных систем, отличающихся по специфичности влияния ионов. Так, максимальная АТФазная активность проявляется при концентрациях ионов: натрия – 115-145, ионов калия – 19-28, ионов магния – 2,0-4,0 ммоль·мл-1. На активность АТФазы не оказывает влияния специфический ингибитор – строфантин-К в диапазоне концентраций 0-100 ммоль*л-1 в среде содержащей ионы Na+ и К+, так и в среде без них.
3. Установлено, что АТФазы, локализованные в ядерных и цитоплазматических мембранах эритроцитов цыплят-бройлеров имеют определенные различия. Так, ионы Na+,K+ детерминируют АТФазную активность цитоплазматических мембран на 97%, ионы Ca2+ - на 87% и HCO3- анион - на 96% с высокой степенью достоверности.
Активность АТФазы ядер не зависела от наличия в среде инкубации ионов Na+,K+ и Ca2+ (P>0,05) и была детерминирована ионами HCO3- на 96% (P<0,01).
4. Активность ферментных систем клеток, тканей и органов зависит от физиологического состояния - эндометрия у свиноматок от стадии полового цикла.
4.1. Максимальная активность АТФаз и АСТ и АЛТ регистрировалась в период стадии возбуждения, а минимальная – в период стадии уравновешивания. Максимальная активность внутриклеточных кислых протеаз, кислых и щелочных фосфатаз в эндометрии свиноматок и маточной слизи регистрируется в период стадии торможения, а минимальная в период стадии уравновешивания. Наибольшая активность ферментов, независимо от стадии полового цикла, отмечена во 2-й трети рогов матки, что связано с развитой сетью спиральных артерий и большим количеством маточных желез в этой части эндометрия.
4.2. Распределение активности АТФаз, КФ и ЩФ в эндометрии, на основании гистохимических исследований, зависит от стадии полового цикла, что согласуется с результатами биохимических исследований:
-в стадии возбуждения проявляется интенсивная реакция на АТФазу в апикальной части покровного эпителия, в эндотелии спиральных артерий и в эпителии желез, так же проявляется хорошо выраженная активность КФ в ядрах поверхностного эпителия и маточных желез, а активность ЩФ, наоборот, локализована преимущественно в апикальной каемке покровного эпителия и по контуру желез;
-в стадии торможения и уравновешивания распределение активности АТФаз становится диффузным, без четко выраженной локализации, а активность КФ преимущественно локализуется в тучных клетках, активность ЩФ перераспределена по всей цитоплазме поверхностного в эпителия.
5. Активность ферментных систем клеток, тканей и органов зависит от кормовых добавок (ПКД и сукцинат натрия). Так, активность Mg2+-АТФазы цитоплазматических мембран была детерминирована ПКД на 28,9%, активность Na+,K+-АТфазы – на 30,7%, Ca2+-АТФаза – 32,8% и HCO3—АТФаза – 10,7%. Сукцинат оказывал наибольшее влияние на активность Ca2+-АТФазы цитоплазматических мембран – 8,2%. Активность АТФаз ядер, практически не зависела от применения ПКД, кроме HCO3--АТФазы, активность которой была на 21,9% детерминирована сукцинатом и на 61,4% при совместном применении ПКД и сукцината.
6. АТФазная активность тканей и органов цыплят-бройлеров неодинакова в различных тканях. По убыванию АТФазной активности ткани и органы можно расположить в следующий ряд: почки, печень, эритроциты, скелетные и сердечные мышцы. В процессе выращивания цыплят (с возрастом) активность АТФазы возрастает. Наибольшие изменения в этом отношении отмечены в почках, далее следует печень и эритроциты и в меньшей степени такие сдвиги наблюдались в мышцах. На динамику АТФазной активности тканей и органов существенное влияние оказывают ПКД и сукцинат. В печени, почках и мышцах наибольший рост активности АТФазы отмечен в группах получавших ПКД. В сердечной мышце наиболее интенсивный рост активности АТФазы происходил в контрольной группе и опытной группе 3, получавшей сукцинат.
7. Активность ферментных систем клеток, тканей и органов зависит от действия чрезвычайного фактора среды - приводящего к воспалительным процессам в эндометрии свиноматок (острому и хроническому).
7.1. Установлено снижение активности АТФаз в мембранных структурах внутриклеточных органоидов. При этом в лизосомах снижение активности АТФаз было более выражено, чем в других органоидах. Протеазы, КФ и АЛТ имеют высокую активность во всех участках тканей рогов матки и в маточной слизи. Активность АСТ снижается во 2-й и 3-й третях рогов матки и в маточной слизи. Активность ЩФ достоверно снижается во всех участках рогов матки и в маточной слизи.
Для хронического эндометрита характерны изменения активности АТФаз в зависимости от локализации в матке и типа внутриклеточных органоидов.
7.2. Воспалительный процесс в эндометрии (острый и хронический) приводит к перераспределению активности АТФаз, КФ и ЩФ:
-в поверхностном эпителии и его апикальной части появляются обширные участки отсутствия АТФазной активности;
-в эндотелии спиральных артерий и в маточных железах выявлено снижение ферментативной активности АТФаз в виде слабой диффузной реакции;
-активность КФ повышается в эпителии и клетках стромы;
-активность ЩФ снижается, а ее распределение в области апикальной каемки поверхностного эпителия и апикальных участках железистых клеток становится прерывистым.
-хронический эндометрит сопровождается повышенной АТФазной активностью, имеющей диффузный характер в поверхностном эпителии, а в маточных железах её распределение носит неравномерный характер.
8. На содержание нуклеиновых кислот – ДНК и РНК и процеесы их аутолиза существенное влияет физиологическое состояние животных, обусловленное половыми циклами. Так, максимальный уровень содержания ДНК и РНК в эндометрии и скорость аутолиза отмечается в стадию возбуждения, а минимальный в стадию уравновешивания полового цикла, что, вероятно связано с циклическими процессами перестройки эндометрия.