
«Физиология нервной системы»
| Вид материала | Реферат |
- Краткая учебно-методическая программа по специальности 030. 301. 65 Психология обоснование, 61.88kb.
- Физиология центральной нервной системы Цель дисциплины, 20.01kb.
- О. В. Физиология центральной нервной системы: Программа, 302.51kb.
- «Анатомия и физиология центральной нервной системы и сенсорных систем», 102.43kb.
- Лекция №1 Общая физиология нервной системы, 311.44kb.
- Пособие содержит словарь физиологических терминов, рисунки, схемы, что поможет студентам, 3193.15kb.
- «Анатомия и физиология центральной нервной системы и сенсорных систем», 256.06kb.
- Программа дисциплины Анатомия и физиология центральной нервной системы для направления, 434.61kb.
- Учебно-методический комплекс дисциплины «анатомия центральной нервной системы» Специальности, 294.67kb.
- Учебно-методический комплекс по дисциплине «физиология центральной нервной системы», 1510.33kb.
Министерство образования и науки Российской Федерации
Новосибирский Государственный Технический Университет
Кафедра инженерных проблем экологии
РЕФЕРАТ
на тему
«Физиология нервной системы»
Выполнила Преподаватель
студентка факультета ЛА Каштанова Е.В.
группы МБ-61
Елисеева Елена
Новосибирск, 2008
Функции центральной нервной системы.
Организм человека представляет собой сложную высокоорганизованную систему, состоящую из функционально связанных между собой клеток, тканей, органов и их систем.
Эту взаимосвязь функций, их согласованное функционирование обеспечивает центральная нервная система. ЦНС регулирует все процессы, протекающие в организме, поэтому с ее помощью происходят наиболее адекватные изменения работы различных органов, направленные на обеспечение той или иной его деятельности.
ЦНС также осуществляет связь организма с внешней средой, путем анализа и синтеза поступающей к ней разнообразной информации от рецепторов. Она выполняет функции регулятора поведения, необходимого в конкретных условиях существования. Это обеспечивает адекватное приспособление к окружающему миру. Кроме того с функциями ЦНС связаны процессы, лежащие в основе психической деятельности человека.
Строение ЦНС.
Структурно-функциональной единицей ЦНС является нейрон (нервная клетка). Он состоит из тела (сомы) и отростков - многочисленных дендритов и одного аксона. Дендриты обычно сильно ветвятся и образуют множество синапсов с другими клетками, что определяет их ведущую роль в восприятии нейроном информации. Аксон начинается от тела клетки аксонным холмиком, функцией которого является генерация нервного импульса, который по аксону проводится к другим клеткам. Аксон сильно ветвится, образуя множество коллатералей, терминали которых образуют синапсы с другими клетками. Мембрана аксона в области синапса содержит специфические рецепторы, способные реагировать на различные медиаторы или нейромодуляторы. Поэтому процесс выделения медиатора пресинаптическими окончаниями может эффективно регулироваться другими нейронами. Кроме того, мембрана окончаний содержит большое число потенциалозависимых кальциевых каналов, через которые ионы кальция поступают внутрь окончания при его возбуждении.
Степень возбудимости разных участков нейрона неодинакова, она самая высокая в области аксонного холмика, в области тела нейрона она значительно ниже и самая низкая у дендритов.
Помимо нейронов в ЦНС имеются глиальные клетки, занимающие половину объема мозга. Периферические аксоны также окружены оболочкой из глиальных клеток - швановских клеток. Нейроны и глиальные клетки разделены межклеточными щелями, которые сообщаются друг с другом и образуют заполненное жидкостью межклеточное пространство нейронов и глии. Через это пространство происходит обмен веществами между нервными и глиальными клетками. Функции клеток глии многообразны:
• они являются для нейронов опорным, защитным и трофическим аппаратом, поддерживают определенную концентрацию ионов кальция и калия в межклеточном пространстве;
• активно поглощают нейромедиаторы, ограничивая таким образом время их действия и другие функции.
Аксоны, помимо функции проведения возбуждения, являются каналами для транспорта различных веществ. Белки и медиатор, синтезированные в теле клетки, органеллы и другие вещества могут перемещаться по аксону к его окончанию. Это перемещение веществ получило название аксонного транспорта. Существует два его вида - быстрый и медленный аксонный транспорт.
Рефлекторная теория.
Основным механизмом деятельности ЦНС является рефлекс. Рефлекс- это ответная реакция организма на действия раздражителя, осуществляемая с участием ЦНС и направленная на достижение полезного результата.
Рефлекс в переводе с латинского языка означает "отражение". Впервые термин "отражение" или "рефлектирование" был применен Р. Декартом (1595-1650) для характеристики реакций организма в ответ на раздражение органов чувств. Он первым высказал мысль о том, что все проявления эффекторной активности организма вызываются вполне реальными физическими факторами. После Р. Декарта представление о рефлексе было развито чешским исследователем Т. Прохазкой, который развил учение об отражательных действиях. В это время уже было отмечено, что у спинальных животных движения наступают в ответ на раздражение определенных участков кожи, а разрушение спинного мозга ведет к их исчезновению.
Дальнейшее развитие рефлекторной теории связано с именем И. М. Сеченова. В книге "Рефлексы головного мозга" он утверждал, что все акты бессознательной и сознательной жизни по природе происхождения являются рефлексами. Это была гениальная попытка ввести физиологический анализ в психические процессы. Но в то время не существовало методов объективной оценки деятельности мозга, которые могли бы подтвердить это предположение И. М. Сеченова. Такой объективный метод был разработан И. П. Павловым - метод условных рефлексов, с помощью которого он доказал, что высшая нервная деятельность организма, так же как и низшая, является рефлекторной.
Структурной основой рефлекса, его материальным субстратом является рефлекторная дуга - совокупность морфологических структур, которая обеспечивает осуществление рефлекса (путь, по которому проходит возбуждение при осуществлении рефлекса).
В основе современного представления о рефлекторной деятельности лежит понятие полезного приспособительного результата, ради которого совершается любой рефлекс. Информация о достижении полезного приспособительного результата поступает в ЦНС по звену обратной связи в виде обратное афферентации, которая является обязательным компонентом рефлекторной деятельности. Принцип обратной афферентации введен в рефлекторную теорию П. К. Анохиным. Таким образом, по современным представлениям структурной основой рефлекса является не рефлекторная дуга, а рефлекторное кольцо, состоящее из следующих компонентов (звеньев):
• рецептор;
• афферентный нервный путь;
• нервный центр;
• эфферентный нервный путь;
• рабочий орган (эффектор);
• обратная афферентация (рис. 8).
Анализ структурной основы рефлекса проводится путем последовательного выключения отдельных звеньев рефлекторного кольца (рецептора, афферентного и эфферентного пути, нервного центра) При выключении любого звена рефлекторного кольца рефлекс исчезает. Следовательно, для осуществления рефлекса необходима целостность всех звеньев его морфологической основы.
Клетки ЦНС имеют многочисленные связи друг с другом, поэтому нервная система человека может быть представлена как система нейронных цепей (нейронных сетей) , передающих возбуждение и формирующих торможение. В этой нервной сети возбуждение может распространяться от одного нейрона на многие другие нейроны. Процесс распространения возбуждения от одного нейрона на многие другие нейроны получил название иррадиации возбуждения или дивергентного принципа распространения возбуждения. Различают два вида иррадиации возбуждения:
• направленная или системная иррадиация, когда возбуждение распространяется по определенной системе нейронов и формирует координированную приспособительную деятельность организма;
• бессистемная или диффузная (ненаправленная) иррадиация, хаотичное распространение возбуждения, при котором координированная деятельность невозможна (рис. 9) .
Рис. 9. Схема явления конвергенции (А) и дивергенции (иррадиации) (Б) возбуждения в центральной нервной системе.
В ЦНС к одному нейрону могут сходиться возбуждения от различных источников. Эта способность возбуждений сходиться к одним и тем же промежуточным и конечным нейронам получила название конвергенции возбуждений (рис.9) .
Основные свойства нервных центров
Нейроны ЦНС для осуществления сложных и многообразных функций объединяются в нервные центры. Нервный центр - это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т. д.) . В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС. Такое объединение (нервный центр в широком смысле слова) позволяет осуществлять наиболее адекватное для конкретных условий осуществления рефлекторной деятельности.
Нервные центры обладают рядом характерных функциональных свойств, обусловленных объединением нейронов в нейронные сети и наличием межнейрональных синапсов.
Основные из этих свойств:
1. Возбуждение в нервных центрах распространяется односторонне - от рецептора к эффектору, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической.
2. Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапс (синаптическая задержка) .
3. В нервных центрах осуществляется суммация возбуждений. Различают два вида суммации:
• временная или последовательная, если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны. В этих условиях ВПСП на постсинаптической мембране суммируются и доводят ее деполяризацию до уровня, достаточного для генерации нейроном потенциала действия;
• пространственная или одновременная - наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы (рис. 10).
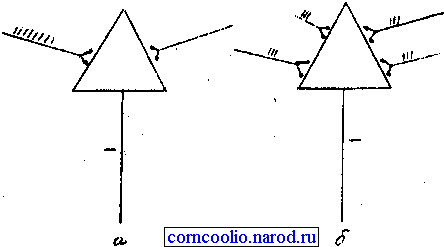
Рис. 10. Схема временной (а) и пространственной (б) суммации возбуждений в центральной нервной системе.
4. Трансформация ритма возбуждения - изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
• понижающая трансформация, в основе которой, в основном, лежит явление суммации возбуждений (пространственной и временной), когда в ответ на несколько возбуждений, пришедших к нервной клетке, в последней возникает только одно возбуждение;
• повышающая трансформация, в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения (рис. 11).
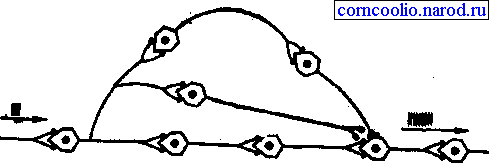
Рис. 11. Схема умножения (мультипликации) возбуждения в центральной нервной
5. Рефлекторное последействие - заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено двумя причинами:
• длительной следовой деполяризацией мембраны нейрона, на фоне которой могут возникать несколько потенциалов действия, обеспечивающих кратковременное рефлекторное последействие;
• пролонгированием выхода возбуждения к эффектору в результате циркуляции (реверберации) возбуждения в нейронной сети типа "нейронной ловушки" (рис. 12). Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление.
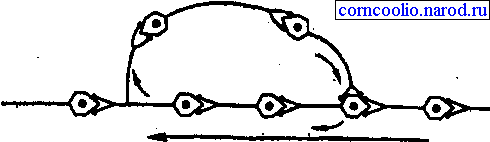
Рис. 12. Схема пролонгирования возбуждения (нейронной ловушки) в центральной нервной системе.
6. Нервные центры, как и синапсы, обладают высокой чувствительностью к недостатку кислорода.
7. Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам.
Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма.
8. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми.
9. Нервные центры, как и синапсы, обладают низкой лабильностью.
10. В нервных центрах легко возникает процесс торможения.
11. Нервные центры обладают тонусом, который выражается в том, что даже при отсутствии специальных раздражении, они постоянно посылают импульсы к рабочим органам.
12. Нервные центры, как и синапсы, обладают низкой аккомодационной способностью, т. е. они способны реагировать на раздражающие факторы, медленно нарастающие по силе.
13. Нервные центры обладают пластичностью - способностью изменять собственное функциональное назначение и расширять свои функциональные возможности.
14. Посттетаническая потенциация - явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Оно обусловлено сохранением определенного уровня ВПСП на нейронах центра, что облегчает проведение последующих возбуждений через синапсы.
Принципы, лежащие в основе координационной деятельности ЦНС.
1. Принцип доминанты был сформулирован А. А. Ухтомским как основной принцип работы нервных центров. Согласно этому принципу для деятельности нервной системы характерно наличие в ЦНС доминирующих (господствующих) в данный период времени очагов возбуждения, в нервных центрах, которые и определяют направленность и характер функций организма в этот период. Доминантный очаг возбуждения характеризуется следующими свойствами:
• повышенной возбудимостью;
• стойкостью возбуждения (инертностью), т. к. трудно подавить другим возбуждением;
• способностью к суммации субдоминантных возбуждений;
• способностью тормозить субдоминантные очаги возбуждения, в функционально различных нервных центрах.
2. Принцип пространственного облегчения. Он проявляется в том, что суммарный ответ организма при одновременном действии двух относительно слабых раздражителей будет больше суммы ответов, полученных при их раздельном действии. Причина облегчения связана с тем, что аксон афферентного нейрона в ЦНС синаптирует с группой нервных клеток, в которой выделяют центральную (пороговую) зону и периферическую (подпороговую) "кайму". Нейроны, находящиеся в центральной зоне, получают от каждого афферентного нейрона достаточное количество синаптических окончаний (например, по 2) (рис. 13) , чтобы сформировать потенциал действия. Нейрон подпороговой зоны получает от тех же нейронов меньшее число окончаний (по 1), поэтому их афферентные импульсы будут недостаточны, чтобы вызвать в нейронах "каймы" генерацию потенциалов действия, а возникает лишь подпороговое возбуждение. Вследствие этого, при раздельном раздражении афферентных нейронов 1 и 2 возникают рефлекторные реакции, суммарная выраженность которых определяется только нейронами центральной зоны (3) . Но при одновременном раздражении афферентных нейронов потенциалы действия генерируются и нейронами подпороговой зоны. Поэтому выраженность такого суммарного рефлекторного ответа будет больше. Это явление получило название центрального облегчения. Оно чаще наблюдается при действии на организм слабых раздражителей.
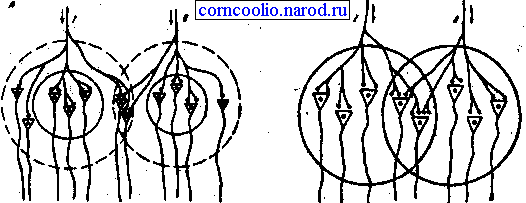
Рис. 13. Схема явления облегчения (А) и окклюзии (Б). Кругами обозначены центральные зоны (сплошная линия) и подпороговая "кайма" (пунктирная линия) популяции нейронов.
3. Принцип окклюзии. Этот принцип противоположен пространственному облегчению и он заключается в том, что два афферентных входа совместно возбуждают меньшую группу мотонейронов по сравнению с эффектами при раздельной их активации. Причина окклюзии состоит в том, что афферентные входы в силу конвергенции отчасти адресуются к одним и тем же мотонейронам, которые затормаживаются при активации обоих входов одновременно (рис. 13). Явление окклюзии проявляется в случаях применения сильных афферентных раздражении.
4. Принцип обратной связи. Процессы саморегуляции в организме аналогичны техническим, предполагающим автоматическую регуляцию процесса с использованием обратной связи. Наличие обратной связи позволяет соотнести выраженность изменений параметров системы с ее работой в целом. Связь выхода системы с ее входом с положительным коэффициентом усиления называется положительной обратной связью, а с отрицательным коэффициентом - отрицательной обратной связью. В биологических системах положительная обратная связь реализуется в основном в патологических ситуациях. Отрицательная обратная связь улучшает устойчивость системы, т. е. ее способность возвращаться к первоначальному состоянию после прекращения влияния возмущающих факторов.
Обратные связи можно подразделять по различным признакам. Например, по скорости действия - быстрая (нервная) и медленная (гуморальная) и т. д.
Можно привести множество примеров проявления эффектов обратной связи. Например, в нервной системе так осуществляется регулирование активности мотонейронов. Суть процесса заключается в том, что импульсы возбуждения, распространяющиеся по аксонам мотонейронов, достигают не только мышц, но и специализированных промежуточных нейронов (клеток Реншоу), возбуждение которых тормозит активность мотонейронов. Данный эффект известен как процесс возвратного торможения.
В качестве примера с положительной обратной связью можно привести процесс возникновения потенциала действия. Так при формировании восходящей части ПД деполяризация мембраны увеличивает ее натриевую проницаемость, которая, в свою очередь, увеличивая натриевый ток, увеличивает деполяризацию мембраны.
Велико значение механизмов обратной связи в поддержании гомеостаза. Так, например, поддержание константного уровня кровяного давления осуществляется за счет изменения импульсной активности барорецепторов сосудистых рефлексогенных зон, которые измененяют тонус вазомоторных симпатических нервов и таким образом нормализуют кровяное давление.
5. Принцип реципрокности (сочетанности, сопряженности, взаимоисключения). Он отражает характер отношений между центрами ответственными за осуществление противоположных функций (вдоха и выдоха, сгибание и разгибание конечности и т. д.). Например, активация проприорецепторов мышцы-сгибателя одновременно возбуждает мотонейроны мышцы-сгибателя и тормозит через вставочные тормозные нейроны мотонейроны мышцы-разгибателя (рис. 18). Реципрокное торможение играет важную роль в автоматической координации двигательных актов.
6. Принцип общего конечного пути. Эффекторные нейроны ЦНС (прежде всего мотонейроны спинного мозга), являясь конечными в цепочке состоящей из афферентных, промежуточных и эффекторных нейронов, могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого числа афферентных и промежуточных нейронов, для которых они являются конечным путем (путем от ЦНС к эффектору). Например, на мотонейронах передних рогов спинного мозга, иннервирующих мускулатуру конечности, оканчиваются волокна афферентных нейронов, нейронов пирамидного тракта и экстрапирамидной системы (ядер мозжечка, ретикулярной формации и многих других структур). Поэтому эти мотонейроны, обеспечивающие рефлекторную деятельность конечности, рассматриваются как конечный путь для общей реализации на конечность многих нервных влияний.
Процессы торможения в центральной нервной системе.
В центральной нервной системе постоянно функционируют два основных, взаимосвязанных процесса - возбуждение и торможение.
Торможение - это активный биологический процесс, направленный на ослабление, прекращение или предотвращение возникновения процесса возбуждения. Явление центрального торможения, т. е. торможения в ЦНС, было открыто И. М. Сеченовым в 1862 г. в опыте, получившим название "опыт сеченовского торможения". Суть опыта: у лягушки на срез зрительных бугров накладывали кристаллик поваренной соли, что приводило к увеличению времени двигательных рефлексов, т. е. к их торможению. Время рефлекса - это время от начала раздражения до начала ответной реакции.
Торможение в ЦНС выполняет две основные функции. Во-первых, оно координирует функции, т. е. оно направляет возбуждение по определенным путям к определенным нервным центрам, при этом выключая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата. Важность этой функции процесса торможения для функционирования организма можно наблюдать в эксперименте с введением животному стрихнина. Стрихнин блокирует тормозные синапсы в ЦНС (в основном глицинергические) и тем самым устраняет основу для формирования процесса торможения. В этих условиях раздражение животного вызывает некоординированную реакцию, в основе которой лежит диффузная (генерализованная) иррадиация возбуждения. При этом приспособителъная деятельность становится невозможной. Во-вторых, торможение выполняет охранительную или защитную функцию, пред охраняя нервные клетки от перевозбуждения и истощения при действии сверхсильных и длительных раздражителей.
Теории торможения. Н. Е. Введенским (1886) было показано, что очень частые раздражения нерва нервно-мышечного препарата вызывают сокращения мышцы в виде гладкого тетануса, амплитуда которого мала. Н. Е. Введенский полагал, что в нервно-мышечном препарате при частом раздражении возникает процесс пессимального торможения, т. е. торможение является как бы следствием перевозбуждения. Сейчас установлено, что его механизм заключается в длительной, застойной деполяризации мембраны, вызванной избытком медиатора (ацетилхолина), выделяющегося при частой стимуляции нерва. Мембрана полностью теряет возбудимость из-за инактивации натриевых каналов и не в состоянии ответить на приход новых возбуждений выделением новых порций медиатора. Таким образом, возбуждение переходит в противоположный процесс - торможение. Следовательно, возбуждение и торможение являются как бы одним и тем же процессом, возникают в одних и тех же структурах, с участием одного и того. же медиатора. Данная теория торможения называется унитарно-химической или монистической.
Медиаторы на постсинаптической мембране могут вызывать не только деполяризацию (ВПСП), но и гиперполяризацию (ТПСП). Эти медиаторы увеличивают проницаемость субсинаптической мембраны для ионов калия или хлора, в результате чего постсинаптическая мембрана гиперполяризуется и возникает ТПСП. Данная теория торможения получила название бинарно-химической, согласно которой торможение и возбуждение развиваются по разным механизмам, с участием тормозных и возбуждающих медиаторов соответственно.
Классификация центрального торможения. Торможение в ЦНС можно классифицировать по различным признакам:
• по электрическому состоянию мембраны - деполяризационное и гиперполяризационное;
• по отношению к синапсу - пресинаптическое и постсинаптическое;
• по нейрональной организации - поступательное, латеральное (боковое), возвратное, реципрокное.
Постсинаптическое торможение развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется. Постсинаптическое торможение может быть деполяризационным, если в его основе лежит процесс длительной деполяризации, и гиперполяризационным, если - гиперполяризации.
Пресинаптическое торможение обусловлено наличием вставочных тормозных нейронов, которые формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению, например, к мотонейрону. В любом случае активации тормозного интернейрона, он вызывает деполяризацию мембраны афферентных терминалей, ухудшающей условия проведения по ним ПД, что таким образом уменьшает количество выделяемого ими медиатора, и, следовательно, эффективность синаптической передачи возбуждения к мотонейрону, что уменьшает его активность (рис. 14). Медиатором в таких аксо-аксональных синапсах является, по-видимому, ГАМК, которая вызывает повышение проницаемости мембраны для ионов хлора, которые выходят из терминали и частично, но длительно ее деполяризуют.
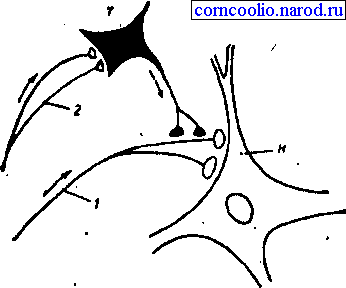
Рис. 14. Пресинаптическое торможение (схема): Н - нейрон, возбуждаемый афферентными импульсами, приходящими по волокну 1; Т - нейрон, образующий тормозные синапсы на пресинаптических разветвлениях волокна 1; 2 - афферентные волокна, вызывающие активность тормозного нейрона Т.
Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения (рис. 15).

Рис. 15. Схема поступательного торможения. Т - тормозньй нейрон
Возвратное торможение осуществляется вставочными тормозными нейронами (клетками Реншоу). Импульсы от мотонейронов, через отходящие от его аксона коллатерали, активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона (рис. 16). Это торможение реализуется за счет тормозных синапсов, образованных клеткой Реншоу на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, которая дает возможность стабилизировать частоту разряда мотонейрона и подавлять избыточную его активность.
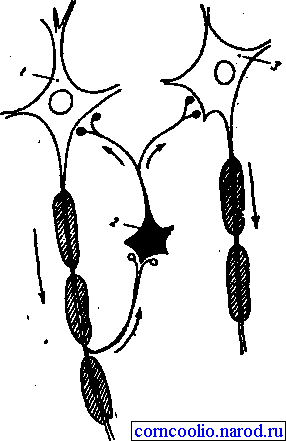
Рис. 16. Схема возвратного торможения. Коллатерали аксона мотонейрона (1) контактируют с телом клетки Реншоу (2), короткий аксон которой, разветвляясь, образует тормозные синапсы на мотонейронах 1 и 3.
Латеральное (боковое) торможение. Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения (рис. 17). В таких случаях возбуждение направляется только по строго определенному пути.
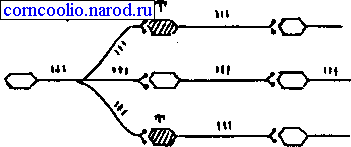
Рис. 17. Схема латерального (бокового) торможения. Т - тормозный нейрон.
Именно латеральное торможение обеспечивает, в основном, системную (направленную) иррадиацию возбуждения в ЦНС.
Реципрокное торможение. Примером реципрокного торможения является торможение центров мышц-антагонистов. Суть этого вида торможения заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны (рис. 18). Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей.
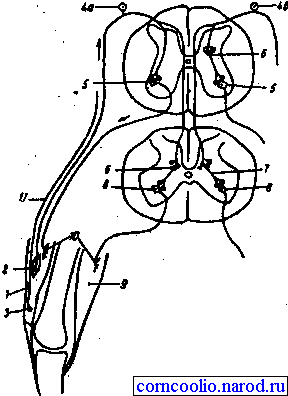
Рис. 18. Схема реципрокного торможения. 1 - четырехглавая мышца бедра; 2 - мышечное веретено; 3 - сухожильный рецептор Гольджи; 4 - рецепторные клетки спиномозгового ганглия; 4а - нервная клетка, воспринимающая импульсы от мышечного веретена; 4б - нервная клетка, воспринимающая имульсы от рецептора Гольджи; 5 - мотонейроны, иннервирующие мышцы-разгибатели; 6 - тормозный промежуточный нейрон; 7 - возбуждающий промежуточный нейрон; 8 - мотонейроны, иннервирующие мышцы-сгибатели; 9 - мышца-сгибатель; 10 - моторные нервные окончания в мышцах; 11 - нервное волокно от сухожильного рецептора Гольджи.