
Н. Н. Алипова, канд биол наук О. В. Левашова и канд биол наук М. С. Морозовой под редакцией акад. П. Г. Костюка москва «мир» 1996 ббк 28. 903 Ф50
| Вид материала | Документы |
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- М. А. Ляшко доц., канд физ мат наук; Т. Н. Смотрова доц., канд, 2299.13kb.
- Современные направления развития физической культуры, спорта и туризма, 4493.6kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Пособие для врачей и среднего медицинского персонала Минск, 5480.63kb.
- Ббк 63. 3(0) Н72, 4378.93kb.
- Тезисы докладов, 4290.75kb.
- Строительные нормы и правила нагрузки и воздействия сниП 01. 07-85* министерство строительства, 1162.86kb.
- Ббк 63. 3(0) Н72, 5546.58kb.
Глава 23. ТКАНЕВОЕ ДЫХАНИЕ. Й. Гроте
23.1. Тканевой метаболизм и потребности тканей в кислороде
Обмен веществ и преобразование энергии в клетках.
Тканевым дыханием называют обмен дыхательных газов, происходящий в массе клеток при биологическом окислении питательных веществ. В ходе окислительных процессов клетки поглощают из капилляров кислород и одновременно выделяют конечный продукт метаболизма - углекислый газ [8]. Здесь термин «тканевое дыхание» использован в более широком смысле, чем в большинстве пособий по биохимии, где им обозначают окислительный распад питательных веществ с участием молекулярного кислорода. Поскольку недостаток О2 лимитирует окислительные реакции значительно сильнее, чем их лимитирует неадекватное удаление СО2, мы будем рассматривать прежде всего процессы, обеспечивающие снабжение тканей кислородом.
Аэробные и анаэробные процессы. Каждая живая клетка нашего организма нуждается в поступлении определенного количества энергии. Эта энергия необходима для поддержания нормальной структуры и жизнедеятельности клеток, а также для выполнения специфических функций. В нормальных условиях клетки получают энергию главным образом путем окислительного (аэробного) разложения питательных веществ. Для осуществления аэробного метаболизма в клетке должны поддерживаться определенные концентрации субстратов (углеводов, белков и жиров) и молекулярного кислорода.
В анаэробных условиях необходимая клетке энергия может быть получена только в процессе гликолиза. Конечный продукт гликолиза-лактатзаключает в себе еще много энергии, поэтому данный путь метаболизма менее экономичен, чем аэробное разложение глюкозы. Для получения одного и того же количества энергии в анаэробных условиях в клетке должно расщепляться примерно в 15 раз больше глюкозы, чем в аэробных.
Согласно Бертону и Кребсу [15], при окислительном распаде 1 моль глюкозы в условиях, примерно соответствующих внутриклеточной среде (t = 25 °С, pH 7,0, РО2 =159 мм рт. ст. = 20 кПа. РСО2 = 50 мм рт. ст. = 5,3 кПа), высвобождается около 689 ккал = 2883 кДж свободной энергии. При разложении того же количества глюкозы в процессе гликолиза выделяется лишь 50 ккал = 208 кДж свободной энергии. Несмотря на низкий энергетический выход, анаэробный распад глюкозы играет важную роль как в анаэробных, так и в аэробных условиях во многих тканях (например, в мозговом веществе почек, хрящах, клетках сетчатки, эритроцитах и работающих мышцах). При изучении обмена веществ в работающих скелетных мышцах, а также во внутренней зоне и сосочках мозгового вещества почек было показано, что в противоположность существовавшим ранее представлениям гликолиз может протекать с высокой скоростью даже в аэробных условиях [17, 18].
Биологическое окисление в митохондриях
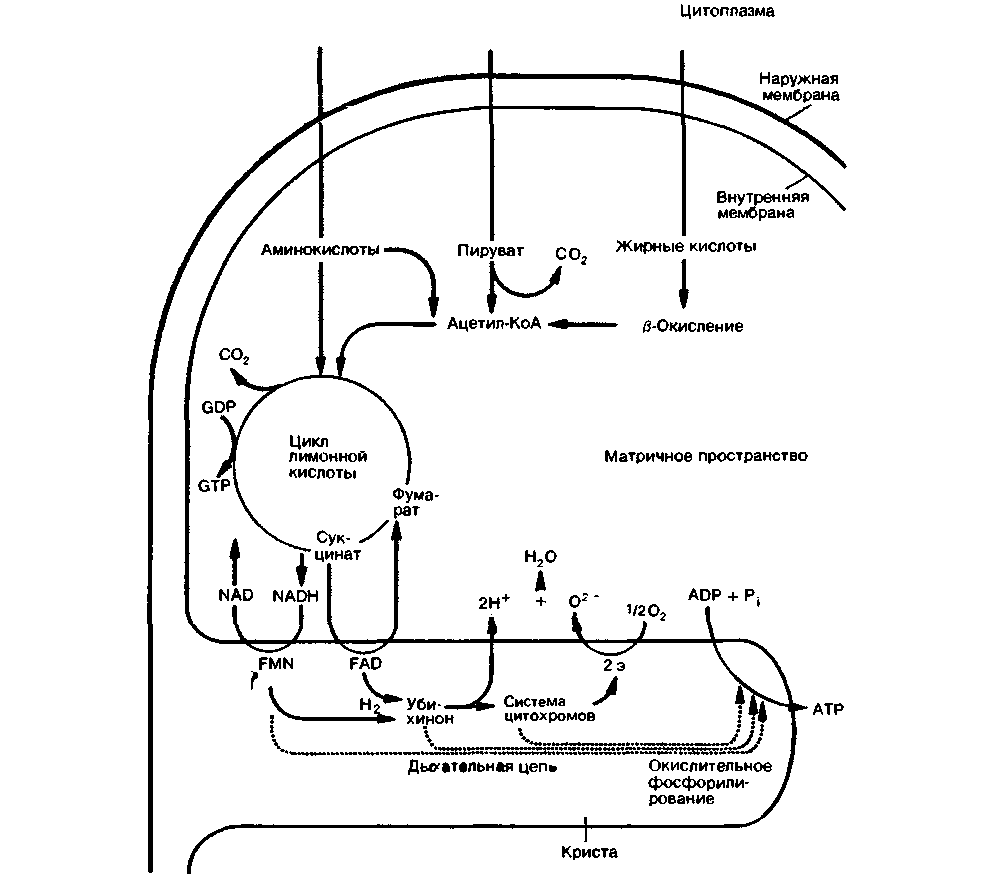
Биологическое окисление происходит в митохондриях. Кроме ферментов цикла лимонной кислоты, дыхательной цепи и окислительного фосфорилирования в этих органеллах были обнаружены также ферменты расщепления жирных кислот и ряда аминокислот [4]. На рис. 23.1 схематично показаны различные пути окислительного метаболизма в митохондриях.
Пируват. жирные кислоты и аминокислоты переносятся из цитоплазмы через митохондриальные мембраны в матрикс митохондрий. После ряда биохимических превращений все эти субстраты распадаются до веществ, которые поступают в цикл лимонной кислоты. Пируват, образующийся главным образом в процессе аэробного расщепления глюкозы в цитоплазме, превращается в результате окислительного декарбоксилирования в матриксе митохондрий в ацетилкофермент А (ацетил-КоА), большая часть которого в нормальных условиях распадается в цикле лимонной кислоты. В отличие от расщепления глюкозы, первый этап которого (гликолиз) протекает в цитоплазме, окислительный распад жирных кислот полностью происходит в матриксе митохондрий. Молекулы
ГЛАВА 23. ТКАНЕВОЕ ДЫХАНИЕ 627
 |
| Рис. 23.1. Схема важнейших этапов биологического окисления в митохондриях |
жирных кислот окисляются в цепи биологических реакций, называемой ß-окислением; образующийся при этом ацетил-КоА поступает в цикл лимонной кислоты либо используется для синтеза жирных кислот. Органические кислоты, образовавшиеся в матриксе митохондрий при метаболизме аминокислот, могут включаться в цикл лимонной кислоты на разных этапах в виде ацетил-КоА, а-кетоглутарата, сукцинил-КоА, фумарата и оксалоацетата.
Образовавшиеся в цикле лимонной кислоты молекулы НАДН (NADH, продукт трех реакций дегидрогенизации) и сукцинат диффундируют из матрикса митохондрий к их внутренней мембране, в которой локализованы ферменты дыхательной цепи и окислительного фосфорилирования. Здесь
НАДН подвергается окислению комплексом ферментов дыхательной цепи, включающим в качестве кофермента флавинмононуклеотид (FMN). Окисление сукцината осуществляется комплексом ферментов дыхательной цепи, в котором роль кофермента выполняет флавинадениндинуклеотид (ФАД, FAD). Оба флавопротеиновых комплекса передают электроны на убихинон (кофермент Q), который затем окисляется с участием цитохромов. В конечном счете из каждой молекулы водорода образуется 2 протона и 2 электрона. Далее электроны переносятся специфическими железосодержащими компонентами дыхательной цепи (цитохром b, FeS-белок, цитохром с1, цитохром с2) на цитохромоксидазу (комплекс
628 ЧАСТЬ VI. ДЫХАНИЕ
цитохромов а и а3), которая передает их на молекулярный кислород. Восстановленный кислород соединяется со свободными ионами водорода с образованием воды. Энергия, высвобождающаяся в процессе переноса электронов по дыхательной цепи, используется для перекачивания протонов из матрикса митохондрий в пространство между их внутренней и наружной мембранами. Перенос протонов приводит к созданию мембранного потенциала на внутренней митохондриальной мембране. За счет тока протонов обратно в матрикс митохондрий индуцируется, согласно теории Митчелла, синтез АТФ (окислительное фосфорилирование). При окислении каждой молекулы НАДН образуются три молекулы АТФ, а при окислении одной молекулы ФАДН2-две молекулы АТФ. В первом случае отношение образования АТФ к потреблению кислорода (коэффициент Р/О) равно 3 (на 1 грамм-атом кислорода образуются 3 моль АТФ), а во втором-2.
Последствия недостаточного снабжения тканей кислородом. При ряде патологических состояний нарушается снабжение тканей кислородом. В этих случаях энергетические потребности клеток могут в течение короткого времени удовлетворяться за счет ограниченных запасов энергии в виде АТФ и креатинфосфата, а также за счет анаэробного процесса-гликолиза. Однако этих источников энергии недостаточно, и они могут использоваться лишь в течение небольшого периода времени, что вызвано двумя основными причинами. Во-первых, в анаэробных условиях потребность клеток в глюкозе настолько возрастает, что чаще всего не может полностью удовлетворяться в течение длительного времени. Во-вторых, в процессе гликолиза образуется в больших количествах лактат, который не столь быстро удаляется из ткани для использования в других органах (например, для расщепления в печени, почках или миокарде или для синтеза гликогена). При сильном недостатке кислорода содержание лактата в тканях и крови постоянно возрастает, что в конечном счете приводит к нереспираторному ацидозу. Когда pH внутриклеточной среды падает ниже уровня, оптимального для активности ферментных систем, наступают резкие нарушения в клеточном метаболизме.
Потребность тканей в кислороде
Потребление О2 в состоянии покоя. Количество кислорода, потребляемого тканью, зависит от функционального состояния входящих в ее состав клеток. В табл. 23.1 приведены данные о потреблении кислорода различными органами и их частями, когда организм находится в состоянии покоя при нормальной температуре. Скорость потребления кислорода тем или иным органом (
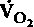 ) обычно
) обычновыражают в мл О2 на 1 г или 100 г массы за 1 мин (при этом учитывается масса органа в естественных условиях). В соответствии с принципом Фика
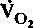 определяют, исходя из кровотока (
определяют, исходя из кровотока (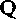 ) через тот или иной орган и разницы в концентрациях О2 в поступающей к органу артериальной крови и оттекающей от него венозной крови (
) через тот или иной орган и разницы в концентрациях О2 в поступающей к органу артериальной крови и оттекающей от него венозной крови (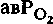 ):
):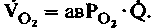 (1)
(1)Когда организм находится в состоянии покоя, кислород относительно интенсивно поглощается миокардом, серым веществом головного мозга (в частности, корой), печенью и корковым веществом почек. В то же время скелетные мышцы, селезенка и белое вещество головного мозга потребляют меньше кислорода (табл. 23.1).
Различия в потреблении кислорода разными участками одного и того же органа. Во многих органах можно измерить кровоток через ограниченные участки ткани путем определения клиренса инертных газов (например, 85Кг, 133Хе и Н2). Таким образом, если возможно взять пробу крови из вены, по которой осуществляется отток от данного участка, то этот метод позволяет определить в нем потребление кислорода. Кроме того, несколько лет назад был разработан метод позитронной эмиссионной томографии (ПЭТ), позволяющий непосредственно измерять кровоток и потребление О2 в отдельных частях органов. Этот метод успешно применяется для исследования головного мозга человека [21, 36]. До внедрения метода ПЭТ, как видно из табл. 23.1, измерить региональное потребление О2 можно было лишь в немногих органах.
При изучении потребления кислорода тканями мозга различных млекопитающих было показано, что кора больших полушарий потребляет от 8 10−2 до 0,1 мл О2 г−1 мин−1. Исходя из потребления О2 целым мозгом и корой, можно вычислить среднее потребление О2 белым веществом мозга. Эта величина составляет примерно 1 10 −2 мл г−1 мин−1. Прямое измерение поглощения О2 участками головного мозга у здоровых испытуемых методом позитронной эмиссионной томографии дало следующие величины: для серого вещества (в различных участках) - примерно от 4 до 6-10−2 мл г−1 -мин−1, для белого вещества-2-10−2 млг−1 мин−1 [21, 36]. Можно предполагать, что потребление кислорода варьирует не только в зависимости от участка, но также в разных клетках одного участка. В самом деле, при измерении (с помощью платиновых микроэлектродов) регионального потребления О2 поверхностными клеточными слоями коры головного мозга было показано, что в условиях слабого наркоза это потребление в пределах небольших участков варьирует примерно от 4-10−2 до 0,12 мл-г−1-мин−1. Результаты радиоавтогра-
ГЛАВА 23. ТКАНЕВОЕ ДЫХАНИЕ 629
| Таблица 23.1. Средние значения скорости кровотока ( 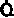 ), артериовенозной разницы по О2 ( ), артериовенозной разницы по О2 (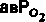 ) и потребления 02 ( ) и потребления 02 (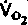 ) в различных органах человека при 37 °С ) в различных органах человека при 37 °С | ||||
| Орган |  | 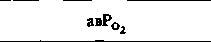 |  | Источник данных |
| Кровь |  |  | 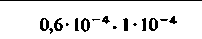 |  |
| Скелетные мышцы: в покое при тяжелой физической нагрузке | 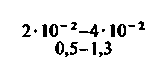 | 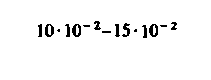 | 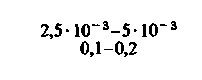 |  |
| Селезенка | 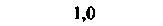 | 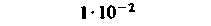 | 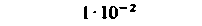 | 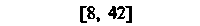 |
| Головной мозг: кора белое вещество | 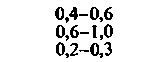 | 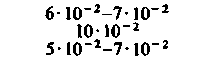 | 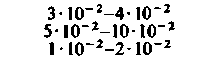 | 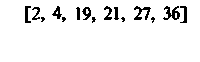 |
| Печень | 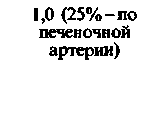 |  | 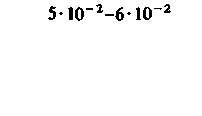 | 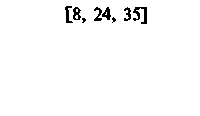 |
| Почки: корковое вещество наружный слой мозгоаого вещества внутренний слой мозгового вещества | 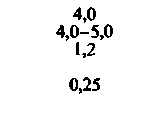 | 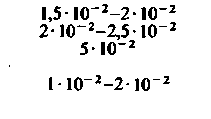 | 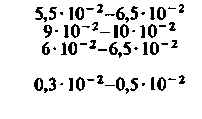 |  |
| Сердце: в покое при тяжелой физической нагрузке | 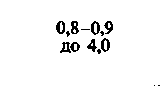 | 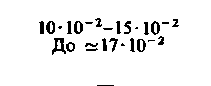 | 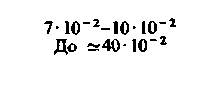 | 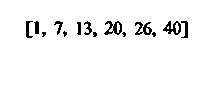 |
 фических исследований регионального кровотока (с использованием иод-14С-антипирина) и регионального потребления глюкозы (с использованием 14С-2дезоксиглюкозы) в коре головного мозга позволяют считать, что эти параметры также существенно различаются в соседних участках [37, 38]. У людей старше 30 лет региональный кровоток и потребление О2 в сером веществе головного мозга с возрастом постепенно снижаются [19, 21, 36]. Примерно такие же различия в потреблении кислорода были обнаружены между отдельными частями почек. В корковом веществе почек среднее потребление О2 в несколько раз больше, чем во внутренних участках и сосочках мозгового вещества. Поскольку потребности почек в кислороде зависят главным образом от интенсивности активной реабсорбции Na+ из просвета канальцев в ткани, полагают, что столь выраженные различия в региональном потреблении О2 обусловлены в основном разницей между величинами этой реабсорбции в корковом и мозговом веществе [11].
фических исследований регионального кровотока (с использованием иод-14С-антипирина) и регионального потребления глюкозы (с использованием 14С-2дезоксиглюкозы) в коре головного мозга позволяют считать, что эти параметры также существенно различаются в соседних участках [37, 38]. У людей старше 30 лет региональный кровоток и потребление О2 в сером веществе головного мозга с возрастом постепенно снижаются [19, 21, 36]. Примерно такие же различия в потреблении кислорода были обнаружены между отдельными частями почек. В корковом веществе почек среднее потребление О2 в несколько раз больше, чем во внутренних участках и сосочках мозгового вещества. Поскольку потребности почек в кислороде зависят главным образом от интенсивности активной реабсорбции Na+ из просвета канальцев в ткани, полагают, что столь выраженные различия в региональном потреблении О2 обусловлены в основном разницей между величинами этой реабсорбции в корковом и мозговом веществе [11].Потребление О2 в условиях повышенной активности органа. В том случае, если активность какоголибо органа по тем или иным причинам повышается, в нем возрастает и скорость энергетического обмена, а следовательно, и потребность клеток в кислороде. При физической нагрузке потребление
О2 тканями миокарда может увеличиваться в 3-4 раза, а работающими скелетными мышцами -более чем в 20-50 раз по сравнению с уровнем покоя. Потребление О2 тканями почек возрастает при увеличении скорости реабсорбции Na+.
В большинстве органов скорость поглощения О2 не зависит от скорости кровотока в них (при условии, что напряжение О2 в тканях достаточно велико). Почки составляют исключение. Существует критическая скорость перфузии, превышение которой вызывает образование ультрафильтрата; при этом уровне фильтрации повышенный кровоток сопровождается повышенным потреблением О2 тканью почек. Такая особенность обусловлена тем, что интенсивность клубочковой фильтрации (а следовательно, и реабсорбции Na+) пропорциональна скорости кровотока.
Зависимость потребления О2 от температуры. Потребление О2 тканями крайне чувствительно к изменениям температуры. При снижении температуры тела энергетический обмен замедляется, а потребность большей части органов в кислороде уменьшается. При нормальной терморегуляции активность органон, участвующих в поддержании теплового баланса, увеличивается, а потребление ими кислорода возрастает. К таким органам относятся, в частности, скелетные мышцы; их терморегуляторная функция осуществляется за счет повышения мышечного тонуса и дрожи (с. 667). Увеличение температуры тела
63β ЧАСТЬ VI. ДЫХАНИЕ
сопровождается повышением потребности большинства органов в кислороде. Согласно правилу Вант-Гоффа, при изменении температуры на 10 oС в пределах от 20 до 40oС потребление тканями кислорода изменяется в том же направлении в 2 3 раза (Q10 = 2-3). При некоторых хирургических операциях бывает необходимо временно остановить кровообращение (а следовательно, снабжение органов О2 и питательными веществами). При этом, чтобы уменьшить потребности органов в кислороде, часто используют гипотермию (понижение температуры тела): больному дают такой глубокий наркоз, при котором терморегуляторные механизмы подавляются.