
Н. Н. Алипова, канд биол наук О. В. Левашова и канд биол наук М. С. Морозовой под редакцией акад. П. Г. Костюка москва «мир» 1996 ббк 28. 903 Ф50
| Вид материала | Документы |
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- М. А. Ляшко доц., канд физ мат наук; Т. Н. Смотрова доц., канд, 2299.13kb.
- Современные направления развития физической культуры, спорта и туризма, 4493.6kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Пособие для врачей и среднего медицинского персонала Минск, 5480.63kb.
- Ббк 63. 3(0) Н72, 4378.93kb.
- Тезисы докладов, 4290.75kb.
- Строительные нормы и правила нагрузки и воздействия сниП 01. 07-85* министерство строительства, 1162.86kb.
- Ббк 63. 3(0) Н72, 5546.58kb.
19.6. Энергетика сокращения сердца
В предыдущих разделах с различных позиций рассматривалась работа сердца. Теперь мы обратимся к количественной стороне этого вопроса и более подробно разберем процессы, отвечающие за снабжение сердца энергией [19]. Прежде всего рассмотрим расход энергии при деятельности сердца.
Мощность и работа сердца
Виды работы сердца. Работа есть произведение силы и расстояния. Единицей работы является джоуль (1 джоуль = 1 ньютон·1 метр, сокращенно Η · м). Эта формула применима, в частности, к работе, совершаемой скелетной мышцей при укорочении и поднимании груза на определенную высоту (работа = вес груза χ высота). Работа сердца в конечном счете также связана с укорочением волокон и развитием усилия. Однако в данном случае происходит не поднятие груза, а перемещение определенного объема крови (V) против сопротивления, создающегося за счет давления (Р). При этом совершается работа по перемещению объема против давления, равная Ρ · V. К этой величине следует прибавить работу по сообщению крови ускорения: эта работа затрачивается на то, чтобы придать инертной массе (m) крови достаточно высокую скорость (ν). Ее вычисляют, исходя из формулы для кинетической энергии: E=1/2mv2.
Расчет работы сердца. В связи с тем что во время систолы отдельные параметры, от которых зависит работа сердца, постоянно меняются, для вычисления этой работы следует проинтегрировать выражения P· V и mv2/2 по времени от начала до конца периода изгнания. Однако мы примем некоторые допущения, упрощающие эту задачу и в то же время позволяющие вычислять работу с достаточной степенью приближения. Можно считать, что P-среднее систолическое давление у выхода из желудочка (1 мм рт. ст. соответствует 133 Н/м2, или 133 Па); V-ударный объем (м2); m - масса крови, которой придано ускорение, т. е. масса ударного объема (кг); ν-средняя линейная скорость выброса (м/с). Значения всех этих параметров для одной систолы следующие.
ГЛАВА 19. ФУНКЦИЯ СЕРДЦА 493
| Работа по перемещению объема против сил давления: P-V |
 |
П
 олная работа А = 1,089 Н-м
олная работа А = 1,089 Н-мРабота по сообщению крови ускорения измеряется в единицах кг-м2-с~2 (Н-м) (см. приложение). В старой литературе работа сердца обычно выражена не в Н-м, а в килограмм-сила·метр (кгс-м). 1 Н-м = 0,102 кгс-м.
Работа левого желудочка по перемещению объема против сил давления значительно больше, чем работа по сообщению крови ускорения: на долю последней приходится лишь около 1 % общей работы. Таким образом, работа сердца в целом во время систолы определяется главным образом величиной ударного объема и давления в аорте; она составляет примерно 1H-м (0,1 кгс-м).
Отношение работы по сообщению крови ускорения к
общей работе сердца может значительно возрастать при увеличении ударного объема, сопровождающемся ускорением кровотока. Это отношение увеличивается также при снижении эластичности аорты в пожилом возрасте, так как уменьшение растяжимости «компрессионной камеры» приводит к падению диастолической скорости кровотока в аорте (рис. 19.31). В этих условиях при систоле сердцу приходится придавать ускорение объему крови, значительно превышающему ударный. Работа по сообщению крови ускорения в таких случаях может становиться почти равной работе по перемещению объема против давления.
Мощность сердца ■ отношение мощности к весу. Мощностью называют работу, совершаемую в единицу времени. Если частота сокращений сердца соответствует одному сокращению в секунду, то мощность сердца равна примерно 1 Вт (Н-м/с), или 0,1 кгс-м/с. Важной характеристикой двигателя любого типа является отношение мощности к весу: для сердца весом около 3 Η это отношение равно
0,3 Вт/Н. Эта величина довольно мала по сравнению с механическими двигателями; так, отношение мощности к весу для автомобильного мотора составляет 15-25 Вт/Н. Однако при мышечной работе мощность сердца может значительно возрастать, и отношение мощности к весу становится почти таким же, как у механических двигателей. Как бы то ни было, приведенные расчеты показывают, что в принципе можно создать такие искусственные насосы, которые при соответствующих условиях могли бы заменить живое сердце, будучи более легкими.
Потребление кислорода я питательных веществ
Энергию, необходимую для совершения механической работы, сердце получает главным образом за счет окислительного разложения питательных веществ. В этом отношении миокард принципиально отличается от скелетных мышц, которые при кратковременных нагрузках могут в значительной мере покрывать свои энергетические потребности за счет анаэробных процессов-образующийся в них «кислородный долг» может быть восполнен в дальнейшем. О значении окислительных процессов для сердца говорит обилие в клетках миокарда митохондрий-органелл, содержащих ферменты окисления.
Потребление кислорода и КПД сердца. Потребление сердцем кислорода in situ обычно определяют, измеряя разницу между содержанием этого газа в артериальной крови и в крови коронарных вен (ав-разница по О2) и умножая полученную величину на объемную скорость кровотока в коронарных сосудах. В покое потребление сердцем кислорода, рассчитанное по этому способу, равно примерно 0,08-0,1 мл г-мин"1. Таким образом, сердце массой 300 г потребляет 24-30 мл О2 в минуту. Это примерно 10% общего потребления кислорода у взрослого человека в покое; масса же сердца составляет лишь около 5% массы тела. При интенсивной работе потребление кислорода миокардом может возрастать в четыре раза по сравнению с покоем. На первый взгляд кажется, что потребление кислорода сердцем должно зависеть в основном от его вклада в общую работу организма за период одной систолы. Однако на самом деле это не так: при одной и той же работе сердце потребляет значительно больше кислорода в том случае, когда оно выбрасывает кровь против повышенного давления, чем когда оно изгоняет больший объем при низком давлении. Таким образом, коэффициент полезного действия сердца (т. е. доля энергии, идущая на совершение механической работы) при нагрузке давлением меньше, чем при нагрузке объемом (рис. 19.35). У здорового сердца КПД составляет в зависимости от преобладания той или иной нагрузки 15-40%.
494 ЧАСТЬ V. КРОВЬ И СИСТЕМА КРОВООБРАЩЕНИЯ
При коронарной недостаточности, когда потребность сердца в кислороде превышает его поступление с кровью, пытаются снизить системное периферическое сопротивление с целью уменьшить артериальное давление и тем самым сократить потребление сердцем кислорода. На этом основано, в частности, терапевтическое действие нитроглицерина во время приступов стенокардии.
Факторы, влияющие на потребление кислорода.
Проведенные исследования показали, что объем кислорода, потребляемый сердцем за одну систолу, зависит прежде всего от напряжения волокон миокарда и что при увеличении периода сокращения этот объем возрастает. Поэтому вошел в употребление показатель время-напряжение, выраженный как произведение среднего напряжения волокон миокарда на длительность систолы. При постоянном размере желудочков можно, исходя из закона Лапласа (с. 480), использовать вместо напряжения волокон среднее систолическое давление в аорте. При колебаниях частоты сокращений сердца потребление О2 изменяется приблизительно в такой же степени, в какой при этом меняется «чистое» рабочее время (произведение длительности систолы на частоту). В связи с этим потребление кислорода примерно пропорционально квадратному корню из частоты сокращений сердца. Остановленное сердце также потребляет некоторое количество кислорода, без которого наступили бы необратимые структурные изменения миокарда. Это так называемое базальное потребление, равное примерно 0,015 мл-г-мин -1 и составляющее лишь незначительную часть от потребления О2 бьющимся сердцем.
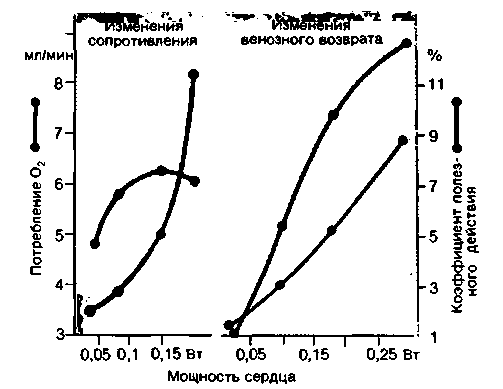 |
| Рис. 19.35. Зависимость потребления кислорода и КПД сердца собаки от развиваемой им мощности при изменениях периферического сопротивления и венозного возврата. Данные получены на сердечно-легочном препарате [по Gollwitzer-Meier, Kroetz. Klin. Wschr., 18, 869 (1939)] |
Потребление питательных веществ. Количественный и качественный состав веществ, используемых сердцем для выработки энергии, может быть определен тем же путем, что и потребление кислорода. Для этого находят разницу концентраций того или иного вещества в артериях и коронарных венах и умножают ее на величину коронарного кровотока. Подобные эксперименты показали, что сердце в отличие, например, от скелетных мышц-«всеядный» орган (рис. 19.36.)
Особенность, которая заслуживает интереса,это значительная доля свободных жирных кислот среди потребляемых сердцем веществ, а также тот факт, что сердце в отличие от скелетных мышц способно использовать молочную кислоту (лактат). При интенсивной физической нагрузке, когда мышцы в результате анаэробного гликолиза высвобождают в кровь большое количество лактата, это вещество служит дополнительным «топливом», необходимым для усиленной работы сердца. Расщепляя молочную кислоту, сердце не только получает энергию, но также способствует поддержанию постоянства pH крови.
Соотношение различных субстратов в общем балансе потребления питательных веществ сердцем зависит главным образом от их поступления (т. е. от их концентрации в артериальной крови). Вследствие такой способности сердца потреблять все доступные вещества главная опасность нарушения коронарного кровообращения заключается не в недостатке субстрата, а в дефиците кислорода.
Макроэргические фосфаты. Метаболическое расщепление различных веществ сопровождается образованием АТФ-непосредственного источника энергии для сокращений сердца. Содержание АТФ в миокарде составляет 4-6 мкмоль/г. Это количество невелико по сравнению с тем, которое требуется для сократительной деятельности миокарда; за несколько секунд работы сердца оно обновляется (т.е. расщепляется с образованием АДФ и неорганического фосфата и вновь ресинтезируется) несколько раз. В сердце обнаружен еще один макроэргический фосфат -креатинфосфат; содержание его примерно равно содержанию АТФ и составляет 7-8 мкмоль/г. Креатинфосфат служит особенно чувствительным показателем снабжения сердца питательными веществами и кислородом, так как от его расщепления зависит вначале метаболический ресинтез АТФ.
Кровоснабжение миокарда
Хотя питающие сердце коронарные сосуды относятся к большому кругу кровообращения (рис. 19.1), они обладают некоторыми особенностями, тесно связанными с деятельностью сердца. В связи с этим мы считаем уместным рассмотреть коронарное кровообращение в настоящей главе. У человека, как
ГЛАВА 19. ФУНКЦИЯ СЕРДЦА 495
 |
| Рис. 19.36. Относительный вклад различных веществ в окислительные обменные процессы сердца при покое и интенсивной работе. Потребление каждого из веществ выражено в процентах как часть общего потребления кислорода, приходящаяся на долю этого вещества [по Keul et al. Pflugers Arch, ges Physiol., 282. 1 (1965)] |
правило, имеются две коронарные артерии, отходящие от основания аорты. Правая коронарная артерия снабжает большую часть правого желудочка, некоторые отделы перегородки и заднюю стенку левого желудочка. Остальные отделы сердца снабжаются с помощью левой коронарной артерии. Отток крови осуществляется преимущественно в венозный синус, остальная кровь оттекает по передним сердечным венам и венам Тебезия.
Величина коронарного кровотока. В опытах на животных коронарный кровоток можно непосредственно измерить при помощи электромагнитных флоуметров. При определении его у человека приходится пользоваться непрямыми методами; некоторые из них основаны на оценке поглощения или разведения в сердце индифферентных газов (ΝΟ2, аргона, ксенона) с учетом известной растворимости этих газов в тканях. Подобные исследования показали, что в покое величина коронарного кровотока у человека равна примерно 0,8-0,9 мл-г · мин-1, что составляет около 5% общего минутного объема. При интенсивной мышечной работе коронарный кровоток может возрастать в четыре раза (табл. 19.2) и примерно во столько же раз увеличивается потребление сердцем кислорода (с. 493).
Колебания коронарного кровотока во время сердечного цикла. Коронарный кровоток в отличие от кровообращения в других органах претерпевает значительные колебания, соответствующие периодам сердечного цикла. Эти периодические колебания обусловлены как пульсирующим характером давления в аорте, так и изменениями напряжения в стенке сердца. Под действием этого напряжения сдавли-
ваются сосуды внутреннего и среднего слоев миокарда. В результате, как показано на рис. 19.37, кровоток в левой коронарной артерии в начале систолы полностью прекращается, и лишь в диастоле, когда напряжение в стенке миокарда падает, он достигает высокого значения. В бассейне правой коронарной артерии внутристеночное напряжение ниже, поэтому кровоток в ней изменяется в основном в соответствии с давлением в аорте. В результате увеличения внутристеночного напряжения отток крови из коронарного синуса во время систолы резко возрастает; во время диастолы он вновь понижается (рис. 19.37).
Регуляция коронарного кровотока. Даже в состоянии покоя сердце извлекает из крови намного больше кислорода, чем другие органы. При содержании О2 в артериальной крови 20 мл/дл экстракция кислорода сердцем составляет около 14 мл/дл (табл. 19.2). Поэтому при повышении нагрузки на сердце возрастающая потребность его в кислороде не может быть покрыта за счет увеличения экстракции кислорода. Повышенная потребность сердца в О2 удовлетворяется главным образом за счет увеличения коронарного кровотока. Это увеличение обусловлено расширением коронарных сосудов, т. е. снижением их гидродинамического сопротивления. Общепризнано, что наиболее мощным стимулом для расширения коронарных сосудов служит недостаток кислорода: дилатация коронарных сосудов наступает уже при снижении содержания О2 в крови на 5%, т.е. примерно на 1 мл/дл. К факторам, вызывающим расширение коронарных сосудов, относят-
| Таблица 19.2 | ||
| А. Нормальные цифры давления в сердце и крупных сосудах у взрослого человека в покое, мм рт. ст. | ||
| Максимальное систолическое давление | Конечнодиастолическое давление | Среднее давление |
| Правое предсердие | | 5 |
| Правый желудочек 25 | 5 | |
| Легочная артерия 25 | 10 | |
| Левое предсердие - | - | 10 |
| Левый желудочек 120 | 10 | |
| Аорта 120 | 70 | |
| Б. Коронарный кровоток и разница по содержанию О2 между артериальной кровью и кровью коронарных вен (авРо ) у человека в покое и при нагрузке | ||
| | В покое При нагрузке | |
| Коронарный кровоток, мл г-1 -мин-1 | 0,8 | 3,2 |
| ав  , мл/дл крови Содержание О2 в коронарных венах, мл/дл крови , мл/дл крови Содержание О2 в коронарных венах, мл/дл крови | 14 6 | 16 4 |
496 ЧАСТЬ V. КРОВЬ И СИСТЕМА КРОВООБРАЩЕНИЯ
 |
| Рис. 19.37. Изменения коронарного кровотока и их связь с систолой, диастолой и давлением в аорте |
ся также аденозин-вещество, играющее важную роль в распаде макроэргических фосфорных соединений [22], и повышение внеклеточной концентрации ионов К+. Прямое действие вегетативных нервов на коронарные сосуды трудно оценить, так как эти нервы одновременно влияют и на другие параметры деятельности сердца. Однако в недавних исследованиях были получены факты, свидетельствующие о прямом сосудосуживающем действии симпатических нервов и сосудорасширяющем парасимпатических нервов.
Факторы эндотелиального происхождения, например окись азота (NO), также способствуют расширению коронарных сосудов. Их выделение из эндотелия повышается при увеличении кровотока (т.е. сил сдвига, действующих на стенки сосудов), а также под действием различных веществ (ацетилхолина, гистамина, серотонина, норадреналина и других), прямой эффект которых тем самым изменяется [15].
Все эти факторы в совокупности образуют регуляторную систему, обладающую высокой надежностью. Недостаток того или другого компонента сильно отражается на регуляции.
Критерии достаточности коронарного кровообращения. Резервы кровоснабжения сердца. Кровоснабжение сердца является достаточным в том случае, если поступление кислорода соответствует потребности в нем. Соотношение этих двух параметров служит критерием достаточности коронарного кровоснабжения. Его значения ниже 1,2 указывают на критическое уменьшение оксигенации сердца (например, при коронарной недостаточности). Для оценки состояния коронарного кровообращения важен не только этот показатель достаточности коронарного кровообращения, но также резервы кровоснабжения сердца, критерием которых служит разница между максимально возможной доставкой О2 и реальным потреблением кислорода в покое, деленная на это реальное потребление. При полноценной способности к адаптации резервы коронарного кровоснабжения в 4-5 раз выше, чем количество, потребляемое в состоянии покоя.
Аноксия и реанимация. В связи с тем что обменные процессы в сердце почти целиком зависят от образования энергии в реакциях окисления, внезапное прекращение коронарного кровотока (ишемия) уже через несколько минут приводит к тяжелым нарушениям деятельности сердца. В опытах с прекращением доставки сердцу кислорода при ненарушенном коронарном кровотоке (аноксии) наблюдаются практически те же изменения: сокращения постепенно ослабляются, полости сердца расширяются и через 6-10 мин наступает остановка сердца. Сильное нарушение системы получения энергии в этих условиях приводит к резкому падению содержания макроэргических фосфатов (креатинфосфата и АТР). Сердце способно в незначительной степени осуществлять анаэробный гликолиз, в результате которого образуется лактат; однако в отсутствие кислорода лактат не метаболизируется, поэтому концентрация его в коронарных венах становится выше, чем в артериях. Если аноксия продолжается больше 30 мин, происходит не только нарушение функции сердца, но и необратимые структурные изменения миокарда. В связи с этим восстановить деятельность сердца по истечении этого срока невозможно. Таким образом, 30 мин -это предельная длительность аноксии, после которой при нормальной температуре тела можно восстановить деятельность сердца. Этот срок, называемый пределом реанимации, может быть значительно увеличен, если снизить скорость обменных процессов путем охлаждения. Такой прием используется в современной кардиохирургии. Если же аноксии подвергается весь организм в целом (например, при удушье), то срок, в течение которого возможна успешная реанимация, меньше, так как уже через 8-10 мин аноксии наступают необратимые повреждения головного мозга, наиболее чувствительного к недостатку кислорода.
ГЛАВА 19. ФУНКЦИЯ СЕРДЦА 497
Сердечная недостаточность
Сердечной недостаточностью называют состояние, при котором сердце не может перекачать столько крови, сколько это необходимо для организма, хотя венозный приток достаточен и компенсаторные механизмы действуют. Такое состояние может наблюдаться даже в покое («недостаточность покоя»), но может становиться ощутимым лишь при физической нагрузке («недостаточность напряжения»). Недостаточность левого желудочка сопровождается застоем крови в легких, который проявляется такими симптомами, как одышка и цианоз; недостаточность же правого желудочка приводит к застою в системных венах, при котором возникают отеки и асцит. К нарушению нагнетательной функции сердца могут приводить различные состояния, не затрагивающие непосредственно сократимость миокарда (клапанные пороки, индурация перикарда, выраженная брадикардия и т.д.).
В узком смысле слова под сердечной недостаточностью понимают снижение сократительной способности сердца (недостаточность миокарда). Такие состояния наблюдаются при хронической перегрузке сердца давлением или объемом (с. 492), а также при его гипоксии (коронаросклероз, инфаркт миокарда), воспалении (миокардит), отравлении некоторыми ядами и передозировке определенных лекарств.
Эти факторы могут приводить к нарушениям в самых различных звеньях процессов возбуждения, электромеханического сопряжения и сокращения в клетках миокарда. Для практических целей особенно важно выделять два типа недостаточности миокарда, различающихся по особенностям метаболизма макроэргических фосфатов, в частности креатинфосфата [15]. Первый тип характеризуется угнетением ресинтеза креатинфосфата вследствие недостаточности выработки энергии; он наблюдается при гипоксии миокарда, действии на него метаболических ядов и т.п. Снижение сократимости в этом случае наступает из-за недостаточного снабжения энергией сократительных белков и связано с уменьшением содержания креатинфосфата. При втором типе недостаточности миокарда запасы макроэргических фосфатов в сердце имеются, но не могут быть эффективно использованы из-за нарушения активации процессов электромеханического сопряжения. Этот тип наблюдается при передозировке антагонистов Са2+, отравлении местными анестетиками, барбитуратами и т. п. Для него характерно высокое содержание креатинфосфата в тканях сердца. Сократимость в этом случае может быть почти полностью восстановлена веществами, улучшающими электромеханическое сопряжение (катехоламинами, сердечными гликозидами). Напротив, при недостаточном снабжении энергией такие препараты могут даже ухудшать состояние; в подобных случаях следует снижать энергозатраты сердца, снижая нагрузку на него.
19.7. Литература
Учебники и руководства
1. Berne R. Μ.. Sperelakis N.. Geiger S. R. (ed.). Handbook of Physiology. Section 2. The Cardiovascular System. Vol. I. The Heart. Bethesda. Amer. Physiol. Soc, 1979.
- Carmeliet E., Vereecke J. Electrogenesis of the action potential and automacity. In. Berne et al. [1].
- Cranefleld P. F. The Conduction of the Cardiac Impulse. Mount Kisco-New York. Futura Publishing Company, 1979.
- Delius W, Gerlach E., Grobecker Η.. Kühler W. (eds.). Catecholamines and the Heart. Berlin, Heidelberg, New York, Springer, 1981.
- Fleckenstein A. Calcium Antagonism in Heart and Smooth Muscle-Experimental Facts and Therapeutic Prospects. New York-Chichester-Brisbane-Toronto- Singapure. Wiley-Interscience Publ., 1983.
- FozzardH.A., Haber E, Jennings R.B., Katz A.M.. Morgan H.E. (eds.). The Heart and Cardiovascular System. New York. Raven Press, 1986.
- Hille B. Ionic Channels of Excitable Membranes. Sunderland Mass. Sinauer, 1984.
- Langer G.A., Brady A.J. (eds.). The Mammalian Myocardium. New York. Wiley, 1974.
- Noble D. The Initiation of the Heartbeat. Oxford. Clarendon Press, 1979.
- Porter R., Fitzsimons D. W. (eds.). Physiological Basis of Starling's Law of the Heart. Ciba Foundation Symposium. Amsterdam-New York. Associated Scientific Publishers, 1974.
- Rüegg J.C. Calcium in Muscle Activation. Berlin-Heidelberg-New York. Springer, 1986.
- Rupp H. (ed.). Regulation of Heart Function - Basic Concepts and Clinical Applications. New York. Thieme Inc., 1986.
Оригинальные статьи и обзоры
- Allessie M.A., Bonke F.. Schopnum F.J.G. Circus movement in rabbit atrial muscle as a mechanism of tachycardia. Circ. Res., 33, 54 (1973).
- Antoni H., Jacob R., Kaufmann R. Mechanical response of the frog's and mammalian myocardium to modifications of the action potential duration by constant pulses. Pflügers Arch., 306, 33 (1969).
- Bassenge Ε., Busse R. Endothelial modulation of coronary tone. Progress in Cardiovasc. Disease, 30, 349 (1981).
- Di Francesco D. A new interpretation of the pacemaker current in calf Purkinje fibers. J. Physiol., 374, 359 (1981).
- Irisawa H., Nakayama Т., Noma A. Membrane currents of single pacemaker cells from pabbit S-A and A-V nodes. In: D. Noble and D. Powell (Edit.). Electrophysiology of Single Cardiac Cells. London. Academic Press, 1987.
- Jacob R., Kissling G.. Ebrecht G.. Holubarsch C. Medugorac I., Rupp H. Adaptive and pathological alternations in experimental cardiac hypertrophy. Adan. Myocardiol., 4, 55 (1983).
- Jacob R., Just H.J., Holubarsch C. (eds.). Cardiac Energetics-Basic Mechanisms and Clinical Implications. Basic Res. Cardiol., 82 (Suppl. 2) (198η.
- Noble D. The surprising heart: A review of recent progress in cardiac electrophysiology. J. Physiol. (Lond.), 353, 1 (1984).
- Reuter H. Exchange of calcium ions in the mammalian myocardium. Mechanisms and physiological significance. Circ. Res., 34, 599 (1974).
- Schroder J. Sites of action and production of adenosine in the heart. In: Burnstock G., Purinergic Receptors. London. Chapmann & Hall, pp. 120 (1981).
- Trautwein W. Membrane currents in cardiac muscle fibres. Physiol Rev., 53, 793 (1973).
- Weidmann S. The diffusion of radiopotassium across intercalated discs of mammalian cardiac muscle. J. Physiol., 187, 323 (1966).