
Альний науковий селекційно-генетичний центр з вівчарства вівчарство міжвідомчий тематичний науковий збірник Випуск 35 Нова Каховка «пиел» 2009
| Вид материала | Документы |
- Науковий вісник, 5717.63kb.
- Збірник «студентський науковий вісник» Довідки, 26.92kb.
- Про затвердження Програми інвестиційного розвитку міста Нова Каховка до 2015 року, 735.83kb.
- Про затвердження Програми інвестиційного розвитку міста Нова Каховка до 2015 року, 740.27kb.
- Перелік публікацій кафедри журналістики за 2009 рік, 185.73kb.
- Bene diagnostitur bene curatur (якісна діагностика якісне лікування)/ М. Денисенко//Науковий, 194.1kb.
- Опис сортів та гібридів сільськогосподарських культур поширених по полтавській області, 2983.38kb.
- Заявка на участь у конференції, 82.92kb.
- Автореферат дисертації на здобуття наукового ступеня, 360.76kb.
- Регіональний науковий юридичний альманах молодих дослідників українська державність, 2685.65kb.
Список використаної літератури
- Законодательство Украины по ведению племенного дела в животноводстве нуждается в гармонизации с мировыми нормами – МинАП / ссылка скрыта
- Проект Закону України “Про внесення змін до деяких законодавчих актів України щодо племінної справи у тваринництві” / ссылка скрыта
- УААН разработан проект новой системы сбора и обработки селекционной информации в животноводстве / ссылка скрыта
- Горлов А. И. Подготовка племенных книг в овцеводстве средствами информационных технологий / А. И. Горлов, Е. А. Ивина, Е. П. Чичаева, И. А. Мокеев, Т. Г. Герасименко // Сб. науч. тр. – Ставрополь: СНИИЖК, 2007 г. - Ч. ІІІ. – С. 102-105.
- Вимоги Європейського Співтовариства щодо запровадження системи ідентифікації і реєстрації тварин /
www.kpf-zorya.com.ua/site/page1539.phpl
- Міністерство аграрної політики України. Наказ № 498 від 31.12.2004 «Про запровадження ідентифікації та реєстрації овець і кіз» / .ua/doc/?uid=1092.112.1&nobreak=1
- Dr. Robert Bambauer. Стратегический подход к разработке систем идентификации животных и контроля их перемещения / www.icar.org/Documents/Kuopio_Presentations/FAO_ICAR_Seminar/Bambauer%20&%20Domenech%20in%20Russian.pdf.
- F.Schmitt. Планирование эксперимента по разработке проектов идентификации и регистрации животных и формирования крупного рогатого скота в странах Центральной и Восточной Европы. – FAO-ICAR Seminar on Animal I&R Development of animal identification and recording systems for veterinary surveillance and livestock development in countries of Eastern Europe Kuopio, Finland, 6 June 2006. – P. 83-90 / tec_series_12_kuopio.pdf.
УДК 636.32/.38: 611.67.018.1
ДО ПИТАННЯ застосування цитовагінального методу при діагностиці РЕПРОДУКТИВНого статусу овець
О.С. Жулінська,
І. В. Лобачова, канд. с.-г. н.
Інститут тваринництва степових районів ім. М.Ф.Іванова „Асканія-Нова” – Національний науковий селекційно-генетичний центр з вівчарства
Наведено характеристику вагінальних мазків вівцематок різних порід, яку пов’язували з функціональністю їх репродуктивної системи. Зокрема, встановлені особливості зміни кількості без’ядерних (ороговіваючих) та суперфіційних (неороговіваючих) епітеліоцитів протягом естрального циклу та у різні сезони року. Також виявлено, що кількість зруйнованих епітеліоцитів та лейкоцитів у мазках залежить від породи овець та зазнає сезонного коливання. Показники часток зруйнованих клітин, голих ядер, клітин з пікнозом ядра та лейкоцити можуть слугувати індикаторами наявності патологічного процесу у статевих органах вівці. У мазках деяких вівцематок виявлялись так звані «пінисті» клітини, зростання кількості яких співпадало з порушенням репродуктивної функції.
Ключові слова: вівці, порода, відтворення, цитовагінальний метод, епітеліоцит
Своєчасне виявлення відхилень у функціонуванні будь-якої системи організму є необхідною умовою успішного проведення профілактичних та лікувальних заходів.
На сьогодні існує багато методів обстеження, які дають можливість визначити стан репродуктивної системи самиць сільськогосподарських тварин - біохімічне тестування сироватки крові, аналіз виділень з статевих шляхів тощо. Але ці методи потребують використання специфічних реактивів, які, на жаль, є дефіцитними на вітчизняному ринку діагностикумів.
Відомо, що гістологічна структура маткового та вагінального епітелію під дією гормонів зазнає змін, які можуть слугувати індикатором функціонального стану яєчників тварин [1]. Це стало підставою для розробки методу, який базується на аналізі зміни характеру кількісного та якісного розподілу епітеліальних клітин у цервікальному слизу. Цей метод був адаптований нами для овець [2]. Доцільність застосування цитовагінального дослідження при обстеженні самиць дрібної рогатої худоби, насамперед овець, обумовлена невеликими розмірами статевої системи, що унеможливлює ректальне обстеження їх внутрішніх статевих органів, як у великих тварин.
Метод вагінального мазка також зручно використовувати у випадках, коли відсутня необхідність визначення точного значення концентрації статевих гормонів або їх похідних у крові, сечі, молоці.
Метою даного дослідження було проаналізувати кількісний та якісний розподіл епітеліальних клітин вагінального мазка і встановити його зв’язок зі станом репродуктивної системи овець.
Матеріал і методика досліджень Дослідження проведено протягом 2005-2008 років на поголів’ї вівцематок асканійської тонкорунної, м’ясововнової та каракульської порід (ДПДГ «Асканія Нова», ДПДГ «Маркеєво»). Відбір цитоматеріалу (серійно або одноразово) здійснювали: у парувальний сезон - під час осіменіння (одноразово) та протягом статевого циклу (серійно з інтервалом у 2-3 доби); у ранній післяродовий період (серійно, починаючи з 12-ої доби після ягніння з інтервалом 2-3 та 7-10 днів); під час глибокого анеструсу – у червні (одноразово) та у червні-липні з інтервалом 3-4 доби і на початку естрального сезону - у липні-серпні (серійно з інтервалом 7-10 діб). Відбір, приготування та мікроскопію вагінальних мазків проводили за розробленою методикою [2]. Усі клітини мазка за своєю морфологією були поділені на такі групи: функціональні епітеліоцити, зруйновані епітеліоцити, без’ядерні (ороговіваючі) епітеліоцити, епітеліоцити з пікнотизованим ядром та лейкоцити. Група функціональних клітин включала в себе підгрупи базальних, проміжних та суперфіційних (неороговіваючих) епітеліоцитів; група зруйнованих – це функціональні клітини з порушеною цілісністю цитоплазми і ядра та голі ядра. Критерієм оцінки вагінального мазка була наявність або відсутність певного типу клітин і їх кількісна частка у мазку, який пов’язували з функціональним станом репродуктивної системи тварин (норма, знижена запліднюваність, відсутність ознак статевого збудження тощо).
Результати досліджень. Обов’язковим елементом вагінального мазка овець є функціональні клітини – епітеліальні клітини типової морфології зі світлою цитоплазмою та чітко окресленим ядром, діаметр якого не менше 6 мкм (Рис.1). Найбільша кількість таких епітеліоцитів відмічалась у мазках ярок та 3-4-річних вівцематок під час статевої охоти – в середньому до 60-75 % від загальної кількості клітин мазка (у окремих тварин до 90 %). Кількість функціональних клітин сягала мінімуму на 3-4 тиждень після ягніння – 40-60 % (залежно від породи), коли йшло деяке гальмування процесів диференціації в епітеліальних тканинах статевих органів. У вівцематок з ознаками порушення репродуктивної функції (перегули, пізній прояв статевого збудження) їх частка під час статевої охоти не перевищувала 35 %.
Показовим компонентом при характеристиці вагінальних мазків овець є підгрупа базальних/парабазальних (клітини глибоких шарів епітелію) (Рис. 1., а) та підгрупа суперфіційних (неороговіваючих або ядерних) епітеліоцитів (Рис. 1., в). Найбільшу кількість клітин глибоких шарів виявляли протягом перших тижнів після ягніння, коли йшов інтенсивний регенераторний процес у слизовій оболонці внутрішніх статевих органів. Особливо це чітко було видно у тонкорунних вівцематок у ранній післяродовий період, у яких частка таких клітин сягала 25-35 % від загальної кількості клітин мазка, або 35-50 % від кількості функціональних клітин. Під час еструсу частка клітин глибоких шарів у мазках овець у нормі не перевищувала 20-22 %. Збільшення цього показника співпадало зі зниженням запліднюваності у вівцематок м’ясововнової породи.
Згідно нашої методики, неороговіваючі ядерні (суперфіційні) епітеліоцити були відокремлені від ороговіваючих епітеліоцитів і разом з підгрупою базальних/парабазальних клітин та підгрупою клітин проміжних шарів (Рис. 1., б) умовно склали групу функціональних клітин. Нами встановлено, що їх відносна частка досить суттєво залежала від фази статевого циклу та сезонності.
Без’ядерні епітеліоцити (ороговіваючі клітини поверхневого шару) не є обов’язковим елементом мазка вівці (Рис. 2., б). Це пов’язано зі специфікою розташування багатошарового плоского неороговіваючого епітелію, який вистилає піхву вівці і обмежується чіткими границями – від складки сечового каналу до входу у цервікальний канал, на відміну від корів та телиць, у яких ороговіваючий епітелій передвір’я “язиками” заходить подекуди аж до вістя шийки матки [3]. У такого типу епітелію кутанізація (ороговіння) більш виражена і у жуйних не пов’язана з фазами статевої активності, як у гризунів [4].
Як показали попередні дослідження, для овець наявність без’ядерних епітеліоцитів є більше індивідуальною ознакою, - вони можуть бути відсутні взагалі або сягати 12-15 % від загальної кількості клітин мазка. У ярок, переярок та деяких вівцематок під час вагінального огляду на початку естрального сезону, а також при осіменінні виявляли пастоподібні білі виділення. При мікроскопії цього секрету поряд із скупченнями клітин різних груп з пікнотизованими ядрами завжди виявляли деяку кількість ороговілих епітеліоцитів – базофільні темні великі без’ядерні клітини із залишками пікнотизованого ядра. Збільшена кількість без’ядерних клітин була ознакою надмірного ороговіння слизової піхви, що зрідка зустрічається у старих вівцематок з порушенням відтворної функції. Слід зазначити, що присутність ороговілих епітеліоцитів у мазках молодих овець, на нашу думку, може бути спричинена складністю відбору цитоматеріалу (з-за вузькості тазу), коли введення інструментів супроводжується деяким зусиллям; з цієї причини клітини епітелію передвір’я можуть захоплюватись тампоном. Такі клітини дещо більші від клітин вагінального епітелію, мають чітко окреслену голубувату або безколірну цитоплазму з рихлим рожевим ядром; ці клітини були виявлені у мазку вівцематки з укороченою піхвою.
 |  |  |
| а | б | в |
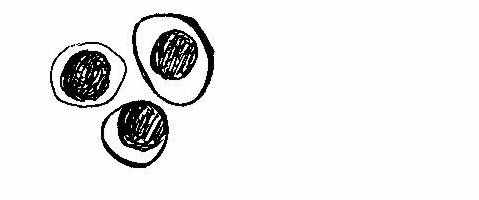
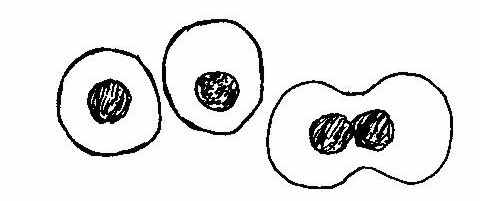
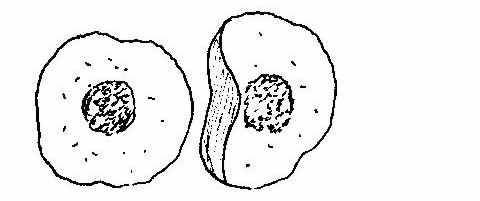
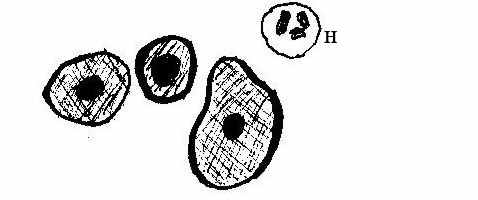
Рис. 1. Функціональні клітини : а - базальні та парабазальні;
б – проміжні епітеліоцити; в – суперфіційні (ядерні) неороговіваючі епітеліоцити.
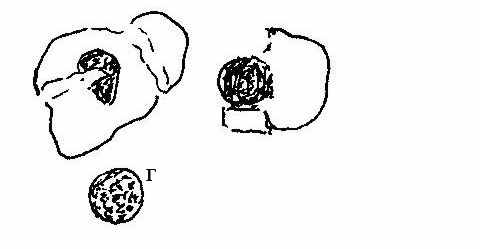
Зруйновані епітеліоцити присутні у мазках усіх тварин (Рис. 2., а). Для тонкорунних вівцематок підвищена їх кількість у післяродовий період була фізіологічною нормою і коливалась від 10-12 до 25 30 %. Взагалі, підвищена кількість зруйнованих епітеліальних клітин у мазках є породною особливістю тонкорунних вівцематок асканійської селекції. Але під час статевої охоти відносна кількість зруйнованих епітеліальних клітин у мазках вівцематок все ж не повинна перевищувати 10 12 %. Зростання частки цих елементів до 25 % і більше під час еструсу виявляли у тварин з ознаками деструктивних процесів у слизовій піхви і порушеннями репродуктивної функції.
При вивченні цитологічної картини вагінальних мазків у тонкорунних вівцематок у естральний сезон відмічене деяке зростання відносної кількості зруйнованих клітин та голих ядер на 11-13-у добу циклу. Припущено, що це може пояснюватись максимальною концентрацією прогестерона в крові овець у цю фазу циклу [5].
У медицині застосовують термін “брудний” мазок, що означає наявність у мазку поряд з епітеліальними клітинами скупчень мікробів, решток дейтрину (внутрішньоклітинної маси, яка утворюється внаслідок руйнування клітин). Все це змінює сприйняття барвника клітинними елементами мазка, ядра клітин при цьому часто розплавлені. “Брудний” мазок при статевій охоті виявляли у кожної другої вівцематки з нерезультативним осіменінням.
 |  |  |
| а | б | в |
| | 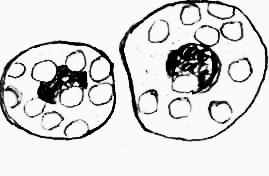 | |
| | г | |
Рисунок 2. Групи клітин: а – зруйновані епітеліоцити; б – без’ядерні; в – клітини з пікнолітизованим ядром, г – «пінисті» клітини.
Голі ядра представляють собою епітеліоцити проміжних або поверхневих шарів вагінального епітелію без видимої цитоплазми (Рис. 2., а-г). Вони або є ознакою порушення формування епітеліоцита, або утворюються внаслідок руйнування клітини з розпаданням цитоплазми навколо ядра. Ці неповноцінні клітини у нормі зрідка зустрічались у мазках вівцематок. Проте, у дослідах 2005-2006 років, коли забезпеченість отари тонкорунних вівцематок кормами у парувальний сезон була нижче 40 %, такий елемент вагінального мазка зустрічався майже у кожної другої тварини. У подальшому, з певним покращенням рівня годівлі, голі ядра у естральних мазках цервікального слизу виявляли зрідка і, в більшості випадків, у старих вівцематок.
Зазвичай в анеструсі голі ядра у вагінальних мазках майже не зустрічались, або їх частка не перевищувала 1-1,5 % від загальної кількості клітин у окремих (частіше старих) тварин. Збільшена кількість голих ядер на 10-12 добу після ягніння (до 3,7±0,86 %) була однією з ознак порушення відтворної функції – 55 % таких вівцематок не запліднились у наступний парувальний сезон [6]. У дослідах 2008 року під час глибокого анеструсу голі ядра були виявлені у 10 % обстеженого поголів’я тонкорунних вівцематок, майже всі ці тварини за результатами попереднього ягніння були яловими.
Епітеліальні клітини з пікнотизованим ядром. До цієї групи клітин мазка належать клітини з базофільним зафарбуванням, темні, з чітким контуром і невеликим щільним ядром діаметром менше 6 мкм (Рис. 2., в). За формою такі клітини відносяться до усіх трьох підгруп функціональних клітин, хоча дещо менші за своїм розміром. У мазках м’ясововнових та каракульських вівцематок протягом місяця після ягніння, коли йшли інтенсивні регенераторні процеси у слизовій оболонці статевих органів, частка таких клітин сягала більше 30 %, що є фізіологічною нормою для цих тварин. Певно в цей час у тварин цих порід відбувається швидке нарощування клітинної маси глибоких шарів епітелію при сповільненні диференціації, яке супроводжується фізіологічно прискореним її відмиранням; при цьому клітини з пікнозом ядра мали розмір базального епітеліоцита і у мазках часто розміщувались у вигляді скупчень. Можливо тому вже через три неділі після ягніння кількісний розподіл клітин у мазках м’ясововнових вівцематок наближався до естрального.
Проте, для тонкорунних вівцематок кількість пікнотизованих ядер у мазку вища за 15 % на 12-14 день після родів була неблагоприємним прогнозом щодо наступного осіменіння.
Під час еструсу частка епітеліоцитів з пікнозом ядра у нормі не повинна перевищувати в середньому 22-25 % від загальної кількості клітин у мазку.
Збільшення кількості пікнотизованих ядер у вагінальному мазку може вказувати на зміну кислотності середовища у піхві вівці і наявність запального процесу або різку зміну гормонального фону організму. Збільшена кількість таких клітин (до 24,8±4,14 %) у мазках вівцематок, віком більше 5 років, прямо співпадала зі зниженням запліднюваності. В окремих дорослих вівцематок поряд з нормальними епітеліоцитами виявляли маленькі темні клітини (розміром як базальний епітеліоцит) з пікнотизованим ядром, які у мазку розташовувалися хаотично або групками. Ця особливість мазка свідчила про порушення формування шарів епітелію, тобто дисплазію. Такі епітеліальні клітини діляться, але не диференціюють і передчасно відмирають. Вівцематки, у мазках яких виявлено такий тип клітин, проявляли ознаки повноцінної статевої охоти, але не запліднювались. В анеструсі загальна цитологічна картина мазка у них залишалась незмінною. На нашу думку, наявність вищезазначених клітин вказує на атрофічні процеси в слизовій оболонці, які можуть бути зумовлені віком або патологічним процесом у статевих органах чи в організмі взагалі. При післязабійному огляді однієї з таких вівцематок було виявлено ущільнені яєчники з ознаками гіпофункції та зменшену матку.
Крім вищезазначених видів клітин, у вагінальних мазках тестували так звані “пінисті” епітеліальні клітини, назву яких взято з кінології. Вони представляють собою епітеліальні клітини проміжних та поверхневих шарів з вакуолізованою цитоплазмою. Зростання частки цих клітин у мазках тонкорунних вівцематок під час статевої охоти супроводжувалось порушенням відтворювальної здатності – такі тварини перегулювали або залишались неплідними. У 18,3 % м’ясововнових вівцематок у вагінальних мазках під час пізнього анеструсу було виявлено “пінисті” епітеліоцити, проте не доведено, що їх присутність була ознакою порушення відтворної функції.
Протягом перших двох тижнів після родів у мазках деяких вівцематок виявляли клітини слизової оболонки цервікального каналу або маткового епітелію різного розміру. Ці клітини розміщувались ізольовано або групками, у вигляді залозистих структур, мали призматичну форму, овальне чи округле ядро, що мало відцентрове розміщення. При збільшенні у 40х на апікальній частині таких клітин були помітні зерна слизу і навіть війки. Їх наявність у мазках через місяць або пізніше після ягніння та під час статевої охоти вказувала на те, що патологічний процес поширився на шийку і матку.
Одним з елементів мазка вагінального слизу вівці також є лейкоцити, які представлені палочкоядерними та сегментоядерними нейтрофілами. Ці клітини потрапляють до вагінальних виділень з міжклітинного простору глибоких шарів багатошарового плоского епітелію піхви, де вони виконують роль складової місцевого тканинного імунітету. Нами спостережено, що у тонкорунних вівцематок лейкоцитів у мазках було завжди більше, ніж у мазках вівцематок інших порід. У сук кількість лейкоцитів є ознакою певної фази статевого циклу. Так, перед тічкою у собак проходить надмірне розростання епітелію у вигляді численних складок, які видно під мікроскопом. Паралельно з цим іде його розрихлення та відшарування, що призводить в подальшому до масового виходу лейкоцитів [7]. З огляду на вищевказане можна зробити припущення про притаманну слизовій оболонці піхви тонкорунних овець складчастість, яка обумовлює вихід значної кількості лейкоцитів. Оскільки попередніми дослідженнями встановлено, що тонкорунні вівцематки, порівняно з м’ясововновими та каракульськими, мають довший період інволюції і більше схильні до виникнення післяродових ускладнень, то цілком ймовірно, що для тварин цієї породи існує більша потреба у присутності клітин, які відповідають за місцевий імунітет [8].
Велику кількість лейкоцитів лімфоїдного ряду у мазках вівцематок виявляли у перші два тижні після родів.
У деяких вівцематок з ознакою гострої форми субінволюції (значні кров’янисті виділення через 2-3 тижні після родів) у вагінальних мазках знаходили навіть клітини Тюрка, що свідчило про наявність гострого запалення.
У разі хронічного запального процесу у статевих органах інколи під час еструсу у мазках певних тварин знаходили гнійні тільця (великі лімфоцити).
Під час статевої охоти лейкоцити нейтрофільного ряду у мазках цервікального слизу не завжди були присутні. За нашими дослідженнями суттєва їх кількість часто співпадала з наявністю у мазках волокон густого слизу. Присутність великої кількості сегментоядерних лейкоцитів під час статевої охоти (більше 2-3 десятків у полі зору, або більше 5 лейкоцитів на один епітеліоцит) є ознакою запального процесу у статевих органах вівці.
Також встановлено, що динаміка кількості лейкоцитів у вагінальних мазках тонкорунних вівцематок залежала від сезону і не залежала від репродуктивного статусу тварини.
Таким чином, виявлено особливості залежності кількісного та якісного розподілу клітин мазка овець від породи, сезону року та репродуктивного статусу.
Висновки. 1. Цитовагінальне дослідження може використовуватись як самостійний метод обстеження стану репродуктивної системи овець.
2. Показники кількості суперфіційних та базальних парабазальних епітеліоцитів змінюються протягом статевого циклу та можуть бути використані для виявлення статевої охоти.
3. Зменшена кількість суперфіційних та збільшена кількість базальних парабазальних клітин під час статевої охоти свідчить про знижену здатність вівці до запліднення.
4. Показник присутності та відносної кількості зруйнованих клітин, голих ядер та клітин з пікнозом ядра є індикатором наявності патологічного процесу у статевих органах вівцематок і може бути використаний при прогнозуванні відтворної функції у овець.
5. Кількість зруйнованих клітин та лейкоцитів у мазку залежить від породи овець та сезону року.
Список використаної літератури
1. Большая медицинская энциклопедия. Т. 5; под ред. В. М. Бакулева. – 2-е изд., перераб. – М.: Мед. лит., 1958. – С. 636-644.
2. Жулінська О. С. Цитовагінальний метод оцінки репродуктивної системи овець / О. С. Жулінська // Вісник аграрної науки. – 2007. - № 4. – С. 75-78.
3. Александровская О. В. Цитология, гистология и эмбриология : учеб. пособие / О. В. Александровская, Т. Н. Радостина, Н. А. Козлов. – М.: Агропромиздат, 1987. – 448 с.
4. Викторов К. Р. Физиология домашних животных / Викторов К. Р. – М.: ОГИЗ – СЕЛЬХОГИЗ, 1948. – 572 с.
5. Прокофьев М.И. Регуляция размножения сельскохозяйственных животных / М. И. Прокофьев – Л.: Наука, 1983. – 264 с.
6. Жулінська О.С. Зв’язок запліднюваності овець з цитолгічною характеристикою вагінальних мазків / О. С. Жулінська // Вівчарство: міжвід. темат. наук. зб. – Нова-Каховка: «Пиєл», 2007. – Вип. 34. – С. 129-134.
7. Ниманд Х.Г., Сутер П.Р. Болезни собак. Практическое руководство для ветеринарных врачей с разделом о болезнях полового аппарата сук Сусанны Арнольд: пер с англ. – М.; «Аквариум», 1998. – 786 с.
8. Жулінська О.С. Особливості перебігу післяродового періоду у вівцематок різних порід / О. С. Жулінська // Проблеми зооінженерії та ветеринарної медицини : зб. наук. праць // ХДЗВА. – Х., 2008. – Вип.16, - Т. 2, Ч.2 – С.257-263.
УДК 636.32/.38.082.451:591.465.1
Вплив тривалості обробки вівцематок фолікулостимулюючим гормоном на кількість і якість прижиттєво вилучених ооцитів
І.В. Лобачова, канд. с.-г. наук
Інститут тваринництва степових районів ім. М.Ф. Іванова «Асканія-Нова» – Національний науковий селекційно-генетичний центр з вівчарства
Досліджена ефективність триденної (дослідна, Д) і дводенної (контрольна, К) схем гормональної стимуляції вівцематок фолікулостимулюючим гормоном («ФСГ-супер», Росія) при серійних операціях з прижиттєвого лапароскопічного вилучення ооцитів. Аспірацію фолікулів проводили на 3-ю добу після вилучення вагінальних песаріїв (МАМ, 30 мг/гол) і на 3-ю (група К) або 4-у (група Д) добу від початку введення ФСГ. Загальна кількість і кількість придатних для подальшого культивування ооцитів, а також їх якість були вищі у контрольних тварин.
Ключові слова: вівця, фолікулогенез, гормональна стимуляція, ооцит, ФСГ, прижиттєве вилучення
Спосіб прижиттєвого лапароскопічного вилучення ооцитів (LOPU) стає прийомом отримання великої кількості гамет від певної вівцематки для проведення робіт з трансплантації ембріонів, особливо при відсутності реакції у тварини на гормональну стимуляцію суперовуляції. Для збільшення кількості вилучених ооцитів дослідниками застосовуються гормональні схеми стимуляції фолікулогенезу в яєчниках, які варіюють від лабораторії до лабораторії. Конкретна схема обробки залежить від сезону року, а також від того, чи планується піддавати вівцематку повторній операції і через який час. Так у дослідах Stangl M. зі спів. [1] для збільшення загальної кількості ооцитів у анестральний сезон вівцематок піддавали повторним LOPU-операціям з інтервалом у сім діб при додатковій обробці гонадотропіном сироватки жеребної кобили (ГСЖК) за 48 годин до операції. Збільшення інтервалу між операціями не покращувало результативність дослідів, проте вело до зменшення кількості ооцитів, вилучених за весь період досліду. Головним недоліком такої схеми була необхідність застосування антисироватки до ГСЖК.
Gibbons A. зі співавт. [2] показали можливість скорочення інтервалу між операціями до 4 діб при додатковій стимуляції фолікулогенезу в яєчниках овець одноразовою ін'єкцією суміші фолікулостимулюючого (ФСГ) і лютеїнізуючого (ЛГ) гормонів без зниження результативності вилучення ооцитів.
Окремим питанням процедури застосування гормональної стимуляції при LOPU-операціях залишається якість вилучених ооцитів. Так Baldassarre H. зі співавт. [3] у дослідах з одноразової аспірації фолікулів овець демонстровано збільшення показника формування хоча б однієї повноцінної бластоцисти на одну оброблену тварину при дробному введенні ФСГ і ЛГ порівняно з їх одноразовою ін’єкцією у тій самій загальній дозі.
Метою наших досліджень є розробка схеми підготовки тварин, яка б поєднували високу ефективність та низьку вартість обробки з можливістю триразового вилучення ооцитів з інтервалом між операціями 10-15 діб. Такий період, на нашу думку, є оптимальним з огляду на необхідність надання тварині часу для заживлення ранок і поновлення стану репродуктивної системи. У попередні роки нами визначено ефективність різних схем підготовки, за якими тварин не стимулювали, або піддавали обробці гестагеними препаратами, чи сумісній обробці гестагенами і одним з гонадотропних препаратів - ГСЖК або ФСГ. Останній вводили за дводенною схемою, що сприяло отриманню достатньо великої загальної кількості ооцитів, але супроводжувалось зниженням кількості вилучених клітин при третій аспірації. За нашим припущенням, причиною цього було збідніння пулу антральних фолікулів в яєчниках через прискорення їх росту під впливом підвищених доз ФСГ і теоретично може бути подолане більш пролонгованим введенням ФСГ.
Метою нашого досліду було порівняння ефективності дво- і триденної схеми обробки вівцематок фолікулостимулюючим гормоном за показниками кількості і якості вилучених ооцитів.
Матеріал і методика досліджень Дослід проведено у травні-червні 2008 року в умовах фізіологічного двору Інституту тваринництва “Асканія-Нова” на 8-ми вівцематках 3-5-річного віку асканійської тонкорунної породи, чотири з яких слугували контролем, а інші чотири склали дослідну групу.
Загальна схема маніпуляцій з тваринами складалася з триразової процедури прижиттєвого вилучення ооцитів з інтервалом між вилученнями 13 діб. Підготовку тварин до першої операції започатковували за 13 діб до неї, до другої і третьої - в день першої та другої аспірації ооцитів.
Випробовані дві схеми підготовки тварин. Схема обробки усіх тварин включала в себе вставку внутрішньовагінальних песаріїв з 30 мг моноацетату мепрогенолу (МАМ) строком на 11 діб, які вилучали за два дні до операції з аспірації ооцитів. Песарії виготовляли самостійно з кусочків паралону діаметром 1,5 см і довжиною 2 см, в середину яких додавали по 30 мг МАМ. Тваринам контрольної групи перед першою та другою операціями, починаючи з дня вилучення песаріїв, ін′єкували через кожні 12 годин препарат “ФСГ-супер” (Росія) за схемою 3+3+3+3 Арморовських одиниць (АО) на вівцематку. Тваринам дослідної групи перед першою та другою операціями ін’єкції фолікулостимулюючого гормону започатковували за 24 години до вилучення песаріїв і вводили шестиразово у дозах 2,25+2,25+2+2+1,5+1,5 АО через кожні 12 годин. Загальна доза ФСГ для тварин обох груп при першій та другій обробках була рівною і становила 12 АО. Перед третьою операцією усім тваринам ін’єктували по 600 ІО препарату ГСЖК («Intergonan», «Intervet», Голандія), який вводили контрольним вівцематкам у день вилучення песаріїв, дослідним - за 24 години до вилучення песаріїв.
Операції з прижиттєвого вилучення проводили з застосуванням лапароскопічної техніки фірми “Richard Wolf” і власно розроблених пристроїв за апробованою методикою з певними змінами: за 15 хвилин до початку операцій тваринам внутрішньом’язево вводили по 0,2 мл препарату “Рометар”. Фолікулярну рідину з ооцитами збирали у середовище Дюльбеко з гепарином (0,01 мг/мл, “Sigma”). Пошук і оцінку ооцитів проводили під мікроскопом при збільшенні у 20 100 разів.
Якість вилучених ооцитів оцінювали за їх морфологією, використовуючи розроблені раніше критерії [4]. Придатними до культивування ооцитами вважали ооцит-кумулюсні комплекси без ознак дегенерації у цитоплазмі та шарі кумулюсних клітин і без розриву зони пелюциду. Оскільки у цитоплазмі більшої частини оголених ооцитів тестували наявність зародкового міхурця, вважали, що такі ооцити позбавлялися кумулюсних клітин в процесі аспірації і, за певних умов, можуть бути використані для культивування. До пустих зон відносили ооцити з розривом зони пелюциду і зони, в середині яких була відсутня внутрішньоклітинна маса.
Результати досліджень обраховували статистично за М.А. Плохінським з обчисленням критерію Стьюдента.
Результати досліджень За результатами дослідів триденна схема введення ФСГ виявилася гіршою за всіма показниками. Так, середня кількість отриманих клітин за операцію на тварину дослідної групи була вірогідно менша від показника тварин контрольної групи як при першому вилученні (p<0,05, td=2,02), так і при другому (p<0,05, td=2,79), а також за результатами двох операцій разом (p<0,005, td=3,78) (табл. 1). За показником придатних до культивування ооцитів вірогідність різниці між групами становила p<0,05 (td=1,89), p<0,05 (td=2,85) і p<0,005 (td=3,69) відповідно.
Таблиця 1. Кількісні показники прижиттєвого вилучення ооцитів контрольних і дослідних овець
| Порядковий номер аспірації | Схема обробки | n | Кількість вилучених ооцитів, шт/гол | |
| всього | придатних до культивування | |||
| перша | дводенна (контрольна) | 4 | 8,5±1,97 | 7,75±1,68 |
| триденна (дослідна) | 4 | 3,75±1,28 | 3,75±1,28 | |
| друга | дводенна | 4 | 9,75±0,87 | 9,25±0,87 |
| триденна | 4 | 4,5±1,67 | 4,25±1,52 | |
| в середньому за двома операціями | дводенна | 8 | 9,13±0,96 | 8,5±0,86 |
| триденна | 8 | 4,13±0,91 | 4,0±0,86 | |
| загалом за дві операції | дводенна | 4 | 18,25±2,37 | 17,0±2,16 |
| триденна | 4 | 8,25±2,23 | 8,0±2,16 | |
| | ||||
| третя | дводенна | 4 | 9,5±2,43 | 7,5±2,24 |
| триденна | 3 | 8,0±3,08 | 4,0±0,71 | |
| | ||||
| в середньому по трьох операціях | дводенна | 12 | 9,25±0,9 | 8,17±0,83 |
| триденна | 11 | 5,18±1,06 | 4,0±0,62 | |
| загалом за три операції | дводенна | 4 | 27,75±3,14 | 24,5±2,89 |
| триденна | 4 | 14,75±1,96 | 11,5±2,69 | |
Загалом, за перші дві операції від тварин дослідної групи отримано вірогідно менше як загальної кількості (p<0,05, td=3,07), так і придатних для культивування ооцитів (p<0,05, td=2,94). Було припущено, що причиною цього могло стати зниження добової дози ФСГ.
При третьому вилученні показник середньої кількості вилучених на тварину ооцитів між групами майже не різнився (p>0,05, td=0,38), але від тварин дослідної групи отримано менше ооцитів, придатних для подальшого культивування (p>0,05, td=1,48). Оскільки при третій обробці доза гонадотропіна між групами не різнилась, скоріше за все основною причиною зниження кількості отриманих ооцитів при триденній схемі введення ФСГ є гальмуючий вплив гестагенів песаріїв, а не зниження добової дози фолікулостимулюючого препарату. Можна припустити, що введення ФСГ під час продовження дії гестагенів веде або до гальмування фолікулогенезу в яєчниках, або до формування неякісних фолікулів, які потім дегенерують.
Аналіз розподілу вилучених ооцитів за морфологічними категоріями за загальними результатами перших двох операцій показав тенденцію до збільшення частки ооцитів І-ої (p>0,05, td=0,38) і зменшення часток ооцитів ІІ-ої категорії (p>0,05, td=1,46) і пустих зон (p>0,05, td=0,91) у тварин дослідної групи, що може свідчити про уповільнення розвитку фолікулів в їх яєчниках (табл. 2). Вірогідне зростання частки ооцитів ІІІ-ої і ІІІа-ої категорій (p<0,05, td=2,13), тобто клітин з ознаками дегенерації у цитоплазмі або оточуючих кумулюсних клітинах, підтверджує наше припущення про негативний вплив гестагенів на розвиток стимульованих дією ФСГ фолікулів. Крім того, припущено, що у тварин контрольної групи під впливом більшої добової дози розвиток фолікулів відбувався більш прискорено, що позначилось зниженням кількості оточуючих ооцитів гранульозних клітин.
Застосування гонадотропіна перед третьою операцією вело до зниження якості ооцитів у тварин обох груп порівняно з використанням очищеного фолікулостимулюючого гормону, що позначилось зниженням майже удвічі частки ооцитів з кумулюсом (І-ої та ІІ-ої категорій), а також значним збільшенням частки пустих зон, і може свідчити про недостатню стимуляцію гонадотропіном проліферації гранульозно-кумулюсного шару в антральних фолікулах.
Таким чином, дводенна схема підготовки тварин-донорів ооцитів за показником кількості і якості отриманих ооцитів виявилася більш ефективною. Враховуючи, що при дводенній схемі ін’єкування ФСГ кількість маніпуляцій з тваринами майже на 30 % менша, ця схема підготовки тварин-донорів ооцитів є більш ефективною і за такими показниками, як затрати праці та зниження стресового впливу на тварину.
Таблиця 2. Розподіл вилучених ооцитів морфологічними категоріями
| Поряд-ковий номер аспірації | Схема обробки | n | Категорія ооцитів, % | ||||
| І | ІІ | ІІІ + ІІІа | IV | пусті зони | |||
| перша | дводенна (контрольна) | 34 | 5,88 | 52,94 | 0,0 | 32,35 | 8,82 |
| триденна (дослідна) | 15 | 13,33 | 33,33 | 20,0 | 33,33 | 0,0 | |
| друга | дводенна | 39 | 7,69 | 61,54 | 0,0 | 25,64 | 5,13 |
| триденна | 18 | 5,56 | 50,0 | 5,56 | 33,33 | 5,56 | |
| разом за дві операції | дводенна | 73 | 6,85 | 57,53 | 0,0 | 28,77 | 6,85 |
| триденна | 33 | 9,09 | 42,42 | 12,12 | 33,33 | 3,03 | |
| | |||||||
| третя | дводенна | 38 | 0,0 | 28,95 | 0,0 | 50,0 | 21,05 |
| триденна | 26 | 3,85 | 19,23 | 3,85 | 26,92 | 46,15 | |
Слід відмітити, що у значної кількості тварин під час операції на яєчниках були присутні сліди свіжих овуляцій. Вказане свідчить про недоцільність такого прийому, як пролонгування дії фолікулостимулюючого гормону до трьох діб з вилученням песаріїв в день початку обробки тварин гонадотропіном. Сама процедура вилучення песаріїв може провокувати передчасну овуляцію і вести до зниження кількості антральних фолікулів на день аспірації. У разі збільшення терміну обробки тварин це може вимагати додаткового застосування препаратів, які б обмежували виділення лютеїнізуючого гормону із гіпофізу, що збільшить загальну вартість обробки.
Результати даного досліду, крім виявлення більш ефективної схеми підготовки вівцематок до операції з прижиттєвого вилучення ооцитів, дають підставу для розуміння причин безуспішності гормональної стимуляції суперовуляції у певних вівцематок. Оскільки за звичай гормональну обробку на суперовуляцію започатковують під час дії вагінальних песаріїв, то однією з причин низької реакції тварин може бути гальмуючий вплив гестагенів песаріїв, що веде до розвитку неповноцінних ооцитів.
Висновки: 1. Дводенна схема введення фолікулостимулюючого гормону при підготовці вівцематок до операцій з прижиттєвого вилучення ооцитів є більш ефективною за показником кількості вилучених ооцитів і їх якості.
2. Причиною зменшення ефективності дії фолікулостимулюю-чого гормону при триденній схеми його введення є гальмуючий вплив гестагенів вагінальних песаріїв.