
Комплекс по дисциплине Чебоксары 2007
| Вид материала | Документы |
- Комплекс по дисциплине Cпециальность 050102 Биология Чебоксары, 1288.79kb.
- Комплекс по дисциплине «генетика» Чебоксары 2006, 821.07kb.
- Учебно-методический комплекс по дисциплине Специальность 100110 Домоведение Чебоксары, 1578.72kb.
- Учебно-методический комплекс по дисциплине Специальность 033100 Физическая культура, 2212.97kb.
- Учебно-методический комплекс дисциплины специальность 033100 (050720) «Физическая культура», 243.87kb.
- Учебно-методический комплекс по дисциплине Cпециальность 050102 Биология Квалификация, 1401.18kb.
- Учебное пособие чебоксары 2007 ббк 81. 02 Б-649, 1167.27kb.
- Учебно-методический комплекс по дисциплине специальность 032300. 00 (050101)- химия, 764.83kb.
- Учебно-методический комплекс по дисциплине специальность 050102 Биология, со специализацией, 615.82kb.
- Учебно-методический комплекс дисциплины «гандбол» Разработан в соответствии с требованиями, 337.21kb.
2.7.5. Тематика курсовых работ
- Изучение флоры дубрав водоохранной зоны Чебоксарского водохранилища
- Изучение флоры дубрав водоохранной зоны р.Волга в пределах Чувашской Республики
- Изучение прибрежной флоры озер ЧР
- Изучение флоры Калининского заказника ЧР
- Изучение флоры дубрав-памятников природы ЧР
- Изучение флоры национального парка «Чаваш вармане»
- Изучение флоры остепненных лугов Присурского степного района ЧР.
- Изучение флоры Заволжского борового района ЧР. Изучение флоры остепненных дубрав ЧР
2.8. Методические указания по организации
изучения дисциплины
Курс «Систематика растений» является составной частью дисциплины «Ботаника с основами фитоценологии» и изучается студентами биологических специальностей педагогических вузов при очной форме обучения в 3 и 4 семестрах, каждый из которых заканчивается экзаменом. По окончании летней сессии 4 семестра студенты проходят полевую практику.
Курс связан с такими дисциплинами, как «Цитология», «Гистология», «Анатомия и морфология растений», «Экология».
Для обеспечения должного уровня преподавания дисциплины необходимо наличие микроскопов, набора готовых микропрепаратов по систематике растений и др. оборудования, указанного в разделе «Материально-техническое обеспечение дисциплины».
2.9. Полевая практика
Летняя полевая практика в природе является одним из важнейших разделов учебных занятий по ботанике. Программа рассчитана на студентов, обучающихся по специальности 050102 – «Биология с дополнительной специальностью». Основными видами деятельности в ходе полевой практики являются экскурсии в природу и последующая камеральная обработка собранного материала в лаборатории.
Основные задачи заключаются в закреплении, расширении и углублении знаний, полученных студентами во время лабораторных занятий по систематике растений с учетом основных вопросов, рассматриваемых при изучении смежных дисциплин; приобретении необходимого научного кругозора и навыков для проведения флористических исследований и геоботанических работ, составление флористических списков, овладение методикой и приемами анализа фитоценозов; овладение навыками природоохранительных мероприятий при проведении экскурсий и сборе ботанического материала, уделяя особое внимание ботаническим объектам, изучаемым в школьном курсе ботаники и редким видам, требующим охраны.
Организационные вопросы проведения практики. Летняя учебная практика по систематике растений с основами фитоценологии осуществляется путем проведения экскурсий в природу, камеральной обработки материала, выполнения самостоятельных наблюдений и индивидуальных работ (оформление рефератов, тематических коллекций и гербария). Базой полевой практики являются разнообразные фитоценозы Чувашии.
Снаряжение и оборудование. Для экскурсий необходимо иметь следующее оборудование: а) папку с гербарной бумагой; б) копалку; в) записную книжку и карандаш; г) лупу;. д) рулетку; е) метровую линейку, ж) весы; з) ведерки или пакеты для растений; и) саперную лопатку, к) 20-и метровые шнуры, л) пресс для сушки растений.
Для проведения камеральной обработки материала необходимо иметь следующее: а) дневник студента; б) определители растений; в) бинокулярные микроскопы МБС-9, препаровальные иглы; г)гербарную бумагу; д) этикетки для гербария; е) бланки описания леса, луга, поля; ж) 10% КОН; з) клей.
Методика изучения растительного покрова
1. Задачи науки о растительности.
2. Определение растительного покрова. Определение фитоценоза. Определение биогеоценоза.
3. Компоненты биогеоценоза.
4. Признаки фитоценоза: а) структура фитоценоза (вертикальное сложение, ярусное расчленение, горизонтальной сложение, сезонная ритмика, смена аспектов); б) динамика и устойчивость фитоценоза (сукцессии, флюктуации, сезонная ритмика).
5. Простейшие методы изучения растительного покрова (пробные площадки, описание рельефа и условий водоснабжения, описание почвенного разреза, структура почвы, формула древостоя, обилие видов, проективное покрытие, ярусность, жизненность видов, бланки описания, хозяйственное использование фитоценозов).
Раннецветущие растения. Особенности строения цветков и соцветий представителей семейств лютиковые, березовые, ивовые, бурачниковые, осоковые, волчеягодниковые, дымянковые. Особенности строения спороносных колосков хвощей. Грибы дискомицеты.
Фитоценоз соснового леса. Географическое положение леса. Условия увлажнения, состав почвы, особенности рельефа, типы леса, методы заложения пробных площадок, формулы древостоя, жизненность. Сомкнутость крон (полнота насаждения). Возраст насаждений. Методы определения высоты дерева. Бонитет. Возобновление, его происхождение. Характеристика подлеска и травянистого яруса. Морфология сосны. Методы определения хозяйственной характеристики леса (запас древесины). Видовой состав наиболее характерных мхов и лишайников. Бланки описания леса.
Фитоценоз елового леса. Морфология ели. Ельники-долгомошники, ельники-черничники, ельники сфагновые. Возобновление ельников. Смены пород. Вертикальная и горизонтальная ярусность. Мхи елового леса.
Растительность болот. Низинные, верховые и переходные болота. Растения болот. Их возникновение. Сфагновые болота. Микрорельеф (кочковатость, средняя высота кочек, мочажины). Особенности морфологии растений сфагновых болот. Болота ивовые, березовые, ольшаники, березово-елово-сосновые. Осоки. Болотные мхи.
Растительность широколиственного леса. Ярусность. Эфемероиды. Аборигенные и заносные виды. Видовой состав травянистых растений лесные злаки и осоки. Возобновление, жизненность. Болезни леса. Бланк описания леса.
Растительность луга. Пойменные и суходольные луга. Способы закладки пробной площади на суходольном луге. Ярусность травостоя. Злаковые и разнотравные луга. Характер злаков по типу кущения. Присутствие мохового покрова. Присутствие мертвой подстилки. Раункьеровские площадки. Хозяйственная характеристика луга: запасы травы и сена, качество сена. Бланк описания луга.
Растительность культурного поля. Искусственные сообщества растений. Их особенности (кратковременность), доминанты – культурные растения, содоминанты – сорные растения. Чистые посевы. Ярусность. Яровые, озимые и многолетние сорняки. Семенная продуктивность. Энергия прорастания сорняков. Определение процента засоренности культурного поля. Бланк описания культурного поля.
Фенологические наблюдения за лекарственными, медоносными, интродуцированными (кормовыми) и редкими видами растений, выращиваемых на АБС (самостоятельная работа).
Выполнение индивидуальной работы по изучению развития вегетативных и репродуктивных органов растений. Агротехника выращивания и уход (валериана лекарственная, родиола розовая, любисток, душица, тимьян ползучий, иссоп, ландыш майский, чистотел, ваточник, мята перечная, тмин, пупавка, гречиха сахалинская, горец Вейриха, сильфия пронзеннолистная, топинамбур, ваточник, окопник, купальница европейская, медуница, лунник оживающий, диплазий сибирский, синюха голубая).
3. ОБЕСПЕЧЕННОСТЬ ДИСЦИПЛИНЫ ЛИТЕРАТУРОЙ
| Ф.И.О. автора (редактора, составителя) Название | Место изд-я | Издательство | Год издания | Кол-во экз. |
| 1 | 2 | 3 | 4 | 5 |
| Гордеева Т.Н. и др. Летняя полевая практика по ботанике | Л. | Просве-щение | 1954 | 1 |
| Куданова З.М. Определитель высших растений Чувашской АССР | Чебоксары | Чув.кн. из-во | 1965 | 35 |
| Методы фенологических наблюдений при ботанических исследованиях под ред. Шульца Г.Э. | М.-Л. | Наука | 1966 | 1 |
| Комарницкий Н.А. и др. Ботаника. Систематика растений | М. | Просве-щение | 1975 | 87 |
| Теплова Л.П. Осенние и весенние дискомицеты | Чебоксары | Чебоксары | 1975 | 10 |
| | | | | |
| Старостенкова М.М.и др. Учебно-полевая практика по ботанике | М. | Просве-щение | 1977 | 30 |
| Старостенкова М.М. и др. Практические работы по систематике растений. Часть 1. Низшие растения Ч.2 Высшие растения | М. | Просве-щение | 1981 | 28 |
| Великанов Л.Л., Гарибова Л.В. и др. Курс низших растений Высшая школа | М. | Высш. школа | 1981 | 2 |
| Тахтаджян А. Система магнолиофитов | Л. | Наука | 1987 | 2 |
| Губанов И.А. и др. Определитель сосудистых растений центра Европейской России | М. | Аргус | 1995 | 19 |
| Теплова Л.П. Мхи разнообразных местообитаний | Чебоксары | Чебоксары | 1997 | 10 |
| Еленевский А.Г. и др. Ботаника высших ,или наземных, растений | М. | Академия | 2000 | 24 |
| Миркин и др. Современная наука о растительности | М. | Логос | 2000 | 4 |
| Миркин Б.М. и др. Высшие растений | М. | Логос | 2001 | 3 |
| Миркин и др. Современная наука о растительности | М. | Логос | 2001 | 2 |
| Программа педвузов. Кол.авторов | Чебоксары | ЧГПУ | 2001 | |
| Красная книга Чувашской Республики. Ч.1. Растения и грибы.Кол.авторов. | Чебоксары | | 2001 | |
4. ФОНДОВЫЕ ЛЕКЦИИ
Тема. Видоизменения мицелия грибов
У грибов иногда можно наблюдать известную дифференцировку гиф мицелия. Отдельные гифы в результате приспособления к выполнению определенных функций несколько изменяют свой вид. Так, у некоторых мукоровых грибов можно обнаружить воздушные дугообразные гифы, которые называют столонами, с помощью которых гриб быстро распространяется по субстрату. Столоны прикрепляются к субстрату ризоидами. Ризоиды – это пучок коротких разветвленных гиф, напоминающих по внешнему виду корни. Однако служат они только для прикрепления к субстрату и развиваются как реакция на соприкосновение с твердым телом. Как только столон перебросился и соприкоснулся с субстратом, то сразу возникают ризоиды. При этом надо подчеркнуть, что ризоиды возникают не только при соприкосновении с питательным субстратом, например, с агаровой средой, но и с любым другим субстратом, даже стеклом. Такую картину можно наблюдать у ризопуса. Таким образом, ризоиды представляют собой пучок коротких пигментированных разветвленных нитей, имеющих форму корней и выполняющих функцию прикрепления.
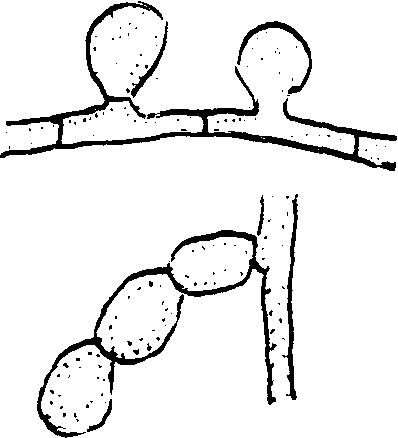
У многих грибов мы наблюдаем образование, так называемых аппрессориев, которые, так же как и ризоиды, выполняют функцию прикрепления. Это тоже специализированные части грибницы. Они встречаются у грибов с эпифитной (поверхностной) грибницей, как, например, представители пор. Эризифовые. Аппрессории имеют вид плоских утолщений на ветвях гиф и могут быть простыми и лопастными и представлять собой одну видоизмененную клетку мицелия, либо могут быть сложными, многоклеточными, как, например, у гриба склеротиния (рис.1). Наблюдать образование аппрессориев довольно просто: если на предметное стекло поместить споры (напр. эризифовых), то через несколько часов спора даст росток, кончик которого при соприкосновении со стеклом образует аппрессорий.
Р

ис. 1 Столоны, ризоиды, аппрессории А – столоны и ризоиды у ризопуса:
1 –ризоиды; 2 – столоны; 3 –5 – спорангий; б – аппрессории эризифы.
 Рис. 2 Несколько напоминают аппрессории так называемые гифоподии (рис.2). Гифоподии имеют вид вздутых одноклеточных коротких боковых ветвей гифы. Одни авторы считают их аналогами аппрессорииев, которые служат для прикрепления к субстрату грибницы, другие – рассматривают гифоподии запасающими органами, где скапливаются питательные вещества.
Рис. 2 Несколько напоминают аппрессории так называемые гифоподии (рис.2). Гифоподии имеют вид вздутых одноклеточных коротких боковых ветвей гифы. Одни авторы считают их аналогами аппрессорииев, которые служат для прикрепления к субстрату грибницы, другие – рассматривают гифоподии запасающими органами, где скапливаются питательные вещества. Различают два типа гифоподиев – головчатые, в виде одноклеточных ветвей и шиповатые, состоящие из короткой боковой ножки, на которой располагается округлая головка из нескольких клеток.
Рис. Гифоподии
Как поверхностный, так и внутренний межклеточный мицелий часто развивает особые специальные ветви – гаустории. Впервые этот орган был обнаружен А.Де Бари, он и ввел это понятие. Гаустории представляют собой своеобразные органы питания и встречаются исключительно у грибов-паразитов (мучнеросных, ржавчинных, пероноспоровых и др.). У мучнеросных грибов, имеющих поверхностный мицелий, гаустории развиваются следующим образом: аппрессории, прикрепляясь к клетке растения-хозяина, выделяет особые ферменты, которые разрыхляют кутикулу, и через эти участки от основания аппрессориев выходит росток, который внедряется в полость клетки растения-хозяина. В гаусторию проникает ядро. Питание паразитного гриба осуществляется только через гаустории, которые выбирают из клетки питательные вещества осмотическим путем, путем всасывания.
У паразитов другой группы – ржавчинных грибов – грибница никогда не бывает поверхностной, а является межклеточной, эндофитной. Ветви такой грибницы в полость клетки развивают гаустории. Гаустории в этом случае являются продолжением вегетативной нити, которая, пройдя в клетку хозяина, изменяет свой внешний облик. Для растительной клетки гриб – это инородное тело, внедрение которого не проходит для нее бесследно. Растительная клетка-хозяин реагирует на процесс внедрения гриба и не дает возможности развиваться гаустории дальше.
Гаустории состоят из узкой части, пронизывающей оболочку клетки в расширенной части, расположенной в полости клетки. Цитологическая картина гаусторий следующая: прежде всего в гаусториях обычно много митохондрий, а также большое число рибосом, что свидетельствует о том, что в гаусториях протекают процессы синтеза; кроме того, между клеточной стенкой гаустории и плазмолеммой клетки-хозяина образуется аморфный слой-чехол (футляр). Гаустория обычно растет по направлению к ядру клетки-хозяина (рис. 3).

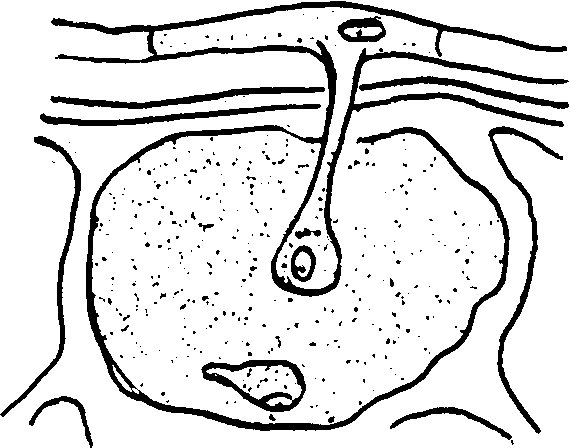 Рис.3. Различная форма гаусторий: а – пероноспора; б – эризифа.
Рис.3. Различная форма гаусторий: а – пероноспора; б – эризифа.Форма гаусторий (ее расширенной части) бывает различной: булавовидной, спиральной и лопастной, встречаются ветвистые ленточные гаустории, они заполняют всю полость клетки (например, у видов Пероноспоровых).
Аналогами гаустории грибов-паразитов можно считать арбускулы грибов-микоризообразователей. Арбускулы представляют собой многократно дихотомически разветвленные гифы, проникающие в паренхимные клетки корня. Считают, что эти видоизмененные гифы образуются под влиянием защитной реакции клеток растения-хозяина.
При обильном ветвлении гифы мицелия могут расти по направлению друг к другу, и в местах соприкосновения нередко происходит слияние их с образованием мостиков. Они носят особое название – анастомозы (рис.4). При обильном развитии анастомозов мицелий принимает вид сеточки. Чаще всего анастомозы развиваются при недостатке питания. Их назначение сводится к следующему: 1) через них происходит обмен веществ; 2) нередко через анастомозы происходит переливание ядер из клетки одной гифы в клетки другой, что приводит к половому процессу у некоторых групп грибов; 3) служат для скрепления гиф мицелия и разрыв грибницы при наличии большого количества анастомозов сильно затруднен.
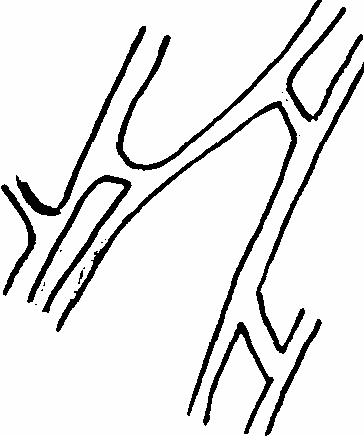
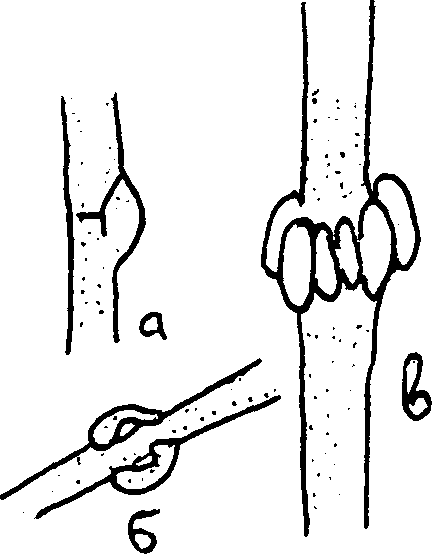 Рис.4 Анастомозы Рис.5. Типы пряжек: а – парные б – мутовчатые В ряде случаев гифы мицелия бывают снабжены пряжками. Это маленькие клетки, лежащие сбоку гифы, против поперечных перегородок. Возникают пряжки следующим образом. На многоклеточной гифе образуется боковой вырост, который тотчас же загибается к основанию клетки и вновь с ней сливается. Пряжка затем отделяется перегородками и становится слепым протоком. Такие пряжки встречаются преимущественно у базидиальных грибов. Их наличие служит систематическим признаком, устанавливающим принадлежность к этой группе грибов. Роль пряжек состоит в том, что через них происходит передача ядра из одной клетки в другую (рис.5).
Рис.4 Анастомозы Рис.5. Типы пряжек: а – парные б – мутовчатые В ряде случаев гифы мицелия бывают снабжены пряжками. Это маленькие клетки, лежащие сбоку гифы, против поперечных перегородок. Возникают пряжки следующим образом. На многоклеточной гифе образуется боковой вырост, который тотчас же загибается к основанию клетки и вновь с ней сливается. Пряжка затем отделяется перегородками и становится слепым протоком. Такие пряжки встречаются преимущественно у базидиальных грибов. Их наличие служит систематическим признаком, устанавливающим принадлежность к этой группе грибов. Роль пряжек состоит в том, что через них происходит передача ядра из одной клетки в другую (рис.5).У некоторых грибов мицелий в присутствии соответствующих животных (личинок нематод) формирует липкие петли, которые, соединяясь друг с другом, образуют сети. Эти сети рассматриваются как приспособления грибов-хищников к улавливанию своей жертвы. К ним следует отнести крючковидные отростки гифы, вздутые клетки, развивающиеся на коротких ветвях мицелия, состоящих из 1-3 клеток, а также разные типы колец-ловушек (пассивное и сжимающее кольцо) (рис.6).
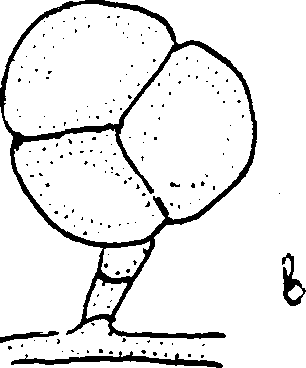

К видоизменениям мицелия относят оидии, представляющие собой различной формы (цилиндрические, эллипсоидальные и др.) клетки, на которые распадается мицелий (рис.7). Чаще всего оидии образуются при неблагоприятных условиях, связанных с кислородным голоданием. При подходящих условиях оидии прорастают в новый мицелий. Таким образом, возникновение оидий это защитное приспособление, служащее для сохранения организма.
Из видоизменений мицелия следует упомянуть еще почкующийся мицелий. В отличие от типичного мицелия, почкующийся мицелий характеризуется тем, что его клетки размножаются не обычным делением, а почкованием. При почковании на поверхности клеток появляется бугорок, или почка, которая постепенно увеличивается в размерах и отделяется затем перегородкой от производящей клетки. В ней, в свою очередь, развиваются такие же почки, и все это повторяется несколько раз. В результате такого почкования получается непрочное соединение круглых или овальных клеток, которое и представляет собой почкующийся мицелий.
Рис.7
В
 некоторых случаях такой мицелий остается коротким и состоит из 2-3 клеток, в других случаях он достигает больших размеров и даже ветвится. Почкование встречается у многих грибов. У одних грибов почко-вание есть результат влияния неблагоприятных условий, как, например, у мукоровых. У других видов – это нормальное, наследственное явление. У дрожжей, например, почкующийся мицелий заменяет грибницу.
некоторых случаях такой мицелий остается коротким и состоит из 2-3 клеток, в других случаях он достигает больших размеров и даже ветвится. Почкование встречается у многих грибов. У одних грибов почко-вание есть результат влияния неблагоприятных условий, как, например, у мукоровых. У других видов – это нормальное, наследственное явление. У дрожжей, например, почкующийся мицелий заменяет грибницу. Грибница нередко образует так называемые хламидоспоры. Они представляют собой толстостенные клетки, возникающие одиночно или группами на вегетативном мицелии. Иногда они обособляются и становятся свободными, не связанными с мицелием. Образуются хламидоспоры следующим образом. Вначале, так же как и при образовании оидий, на мицелии появляются более многочисленные дополнительные поперечные перегородки. Затем содержимое одной клетки или нескольких клеток переливается в соседнюю или соседние. После этого протоплазма отслаивает собственную оболочку под старой оболочкой гифы, образовавшиеся клетки и представляют собой хламидоспоры. После разрушения оставшихся пустыми частей гиф они освобождаются. Хламидоспоры выдерживают высыхание и действие других неблагоприятных условий и могут сохраняться жизнеспособными до 10 и более лет. Они содержат значительные запасы питательных веществ (жиров, углеводов), оболочка их плотная, часто пигментированная и имеет различную скульптуру. Отличие хламидоспор от оидий заключается в том, что, во-первых, хламидоспоры имеют собственную оболочку и, во-вторых, прорастают они чаще всего с образованием органов спороношения, тогда как оидии прорастают в мицелий, Хламидоспоры распространены у очень многих грибов.
Хламидоспоры могут быть в виде, отдельных клеток или в виде клубочков (спорокучек). Так, у одних видов хламидоспоры одиночные, очень мелкие с гладкой или шиповатой оболочкой, тогда как у других видов они представлены клубочками. Обычно клубочки состоят из 1-3 или более типично пигментированных клеток, вокруг которых располагаются мелкие, тонкостенные, бесцветные, лишенные содержимого клетки, последние служат для более легкого распространения хламидоспор, т.е. выполняют роль парашюта (рис.8).
Рис.8
Хламидоспоры
головневых грибов
 Аналогами хламидоспор являются везикулы, встречающиеся у грибов-микоризообразователей. Везикулы - это пузыревидные вздутия, содержащие капли жира (рис.9). Им приписывают функцию запасания питательных веществ, возникают они как вне корня (внекорневые везикулы), так и внутри корня (внутрикорневые везикулы). После разрушения корня они могут служить также и органами размножения.
Аналогами хламидоспор являются везикулы, встречающиеся у грибов-микоризообразователей. Везикулы - это пузыревидные вздутия, содержащие капли жира (рис.9). Им приписывают функцию запасания питательных веществ, возникают они как вне корня (внекорневые везикулы), так и внутри корня (внутрикорневые везикулы). После разрушения корня они могут служить также и органами размножения. 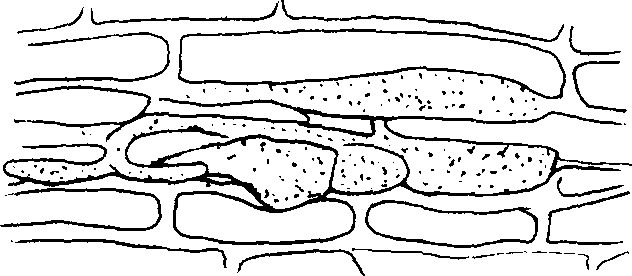 Г
Г рибы нередко формируют пленки. Они представляют собой слой плотно переплетенных гиф, причем гифы здесь расположены в разных направлениях. Пленки бывают разной толщины – от нескольких миллиметров до 0,5 см.
рибы нередко формируют пленки. Они представляют собой слой плотно переплетенных гиф, причем гифы здесь расположены в разных направлениях. Пленки бывают разной толщины – от нескольких миллиметров до 0,5 см.Рис.9. Везикулы у грибов-микоризообразователей
1- интеркалярные везикулы; 2- несколько везикул в межклеточном пространстве
Они образуются на поверхности субстрата или в трещинах древесины. Пленки формируют очень многие грибы, особенно на поверхности жидких питательных сред в чистых культурах. Однако в природных условиях они встречаются преимущественно у трутовиков.
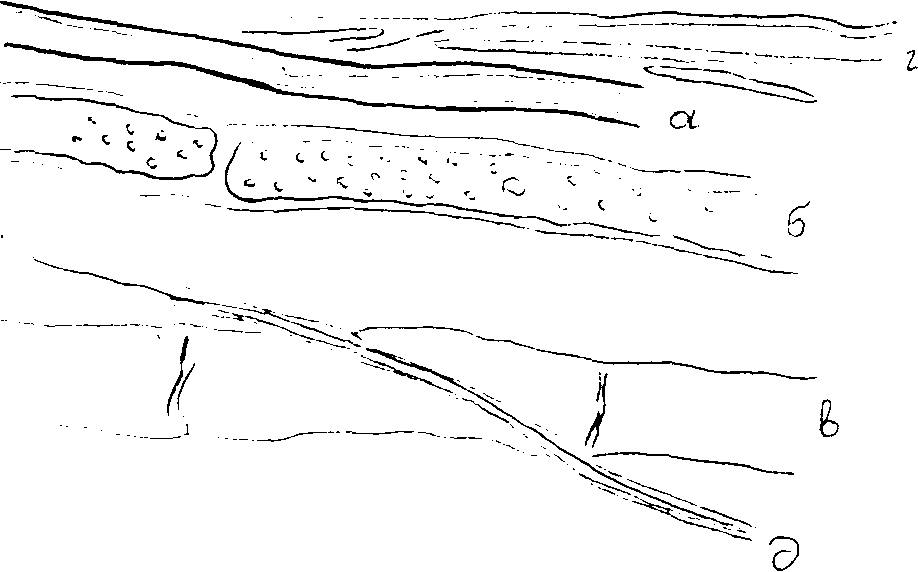 У грибов в качестве сложного и специализированного сплетения грибницы следует указать шнуры и тяжи. Они состоят в противоположность пленкам из параллельно расположенных гиф мицелия. Здесь уже наблюдается либо срастание гиф вегетативного мицелия, либо скрепление этих гиф с помощью коротких анастомозов. Можно выделить несколько типов шнуров. В простейшем случае они состоят из небольшого числа параллельно идущих однородных (одинакового строения) гиф, срастающихся друг с другом ослизненными наружными оболочками или образующих короткие аностомозы. В других случаях, гифы, составляющие шнуры, получают известную дифференцировку. Она выражается в том, что наружные элементы, образующие кору, слагаются из мелких, но прочных, окрашенных в темный цвет гиф, а внутренние, образующие сердцевину, состоят из более широких бесцветных гиф. Наибольшей дифференцировки достигает строение шнура у домового гриба (рис.10).
У грибов в качестве сложного и специализированного сплетения грибницы следует указать шнуры и тяжи. Они состоят в противоположность пленкам из параллельно расположенных гиф мицелия. Здесь уже наблюдается либо срастание гиф вегетативного мицелия, либо скрепление этих гиф с помощью коротких анастомозов. Можно выделить несколько типов шнуров. В простейшем случае они состоят из небольшого числа параллельно идущих однородных (одинакового строения) гиф, срастающихся друг с другом ослизненными наружными оболочками или образующих короткие аностомозы. В других случаях, гифы, составляющие шнуры, получают известную дифференцировку. Она выражается в том, что наружные элементы, образующие кору, слагаются из мелких, но прочных, окрашенных в темный цвет гиф, а внутренние, образующие сердцевину, состоят из более широких бесцветных гиф. Наибольшей дифференцировки достигает строение шнура у домового гриба (рис.10).Рис.10. Часть шнура домового гриба: а, б, в – сосудовидные гифы; г – гифы основной ткани, д – механические гифы
Здесь различают обыкновенные тонкостенные гифы, они служат для продвижения питательных веществ; гифы толстостенные с узким просветом и сильно утолщенными оболочками придают шнуру прочность; гифы с широким просветом служат для продвижения воды, у таких гиф нет поперечных перегородок, они растворены, иногда вместо них сохраняются небольшие бугорки, выросты, представляющие собой остатки поперечных перегородок.
Рис.11
Ш
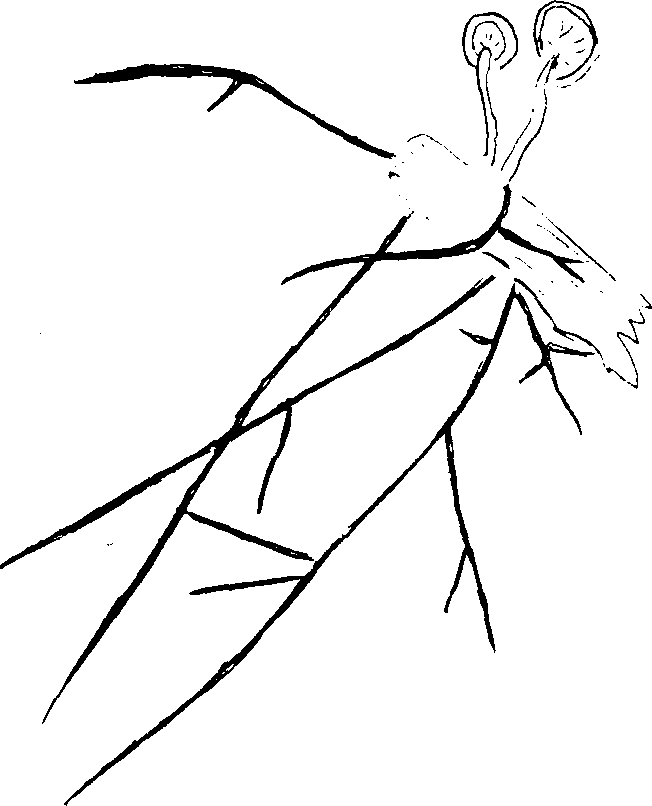 нуры, имеющие сходство с корнями высших растений носят название ризоморфы. Они развиваются чаще всего в почве и простираются на большие расстояния. Ризоморфы, как и шнуры, состоят из вегетативных гиф, которые располагаются параллельно. Поверхностные слои ризоморф состоят из одного или нескольких слоев отмерших клеток, окрашенных в темный цвет, а внутренние – из живых, тонкостенных, бесцветных гиф. Часто такие ризоморфы встречаются у опенка (рис. 11).
нуры, имеющие сходство с корнями высших растений носят название ризоморфы. Они развиваются чаще всего в почве и простираются на большие расстояния. Ризоморфы, как и шнуры, состоят из вегетативных гиф, которые располагаются параллельно. Поверхностные слои ризоморф состоят из одного или нескольких слоев отмерших клеток, окрашенных в темный цвет, а внутренние – из живых, тонкостенных, бесцветных гиф. Часто такие ризоморфы встречаются у опенка (рис. 11).Рис.11. Ризоморфы опенка
Шнуры и ризоморфы нарастают, как и гифы, из которых они состоят, своими вершинами, и могут достигать нескольких мм толщ. и нескольких метров длины. Благодаря своему значительному разра-станию они способствуют вегетативному распространению гриба от дерева к дереву по тяжам и ризоморфам происходит передвижение питательных веществ к плодовым телам; ризоморфы, защищенные плотной корой служат для сохранения гриба при неблагоприятных условиях. Они могут высыхать и при наступлении подходящих условий прорастают и развивают новый мицелий.
Под названием склероций понимают такое уплотнение грибных гиф, при котором получается твердое тело. Склероции представляют собой темноокрашенные тверды тела, состоящие из плотного сплетения мицелиальных гиф. Размер склероциев может быть различным – от микроскопически малых до 20-30 см в диаметре и до 20 кг весом. Некоторые склероции, образованные тропическими трутовиками, употребляются в пищу. Склероции могут быть различной формы – шаровидной, овальной, в виде рожков или иметь совершенно неправильную форму. Очень любопытна, например, форма склероциев сформированных в корзинках подсолнечника. Образующиеся между семянками склероции сливаются между собой и формируют сетчатого строения тело.
Образование склероциев можно проследить на моркови. Несколько немытых морковок следует поместить во влажную камеру, где через несколько дней будет образован белый мицелий гриба. На уплотненном рыхлом мицелии формируются клубочки белого цвета, которые постепенно уплотняются, выдавливая на поверхность капли воды, затем эти клубочки пигментируются и становятся твердыми темноокрашенными телами-склероциями. Такие склероции служат для сохранения гриба в неблагоприятных условиях и для его распространения.
Склероции могут сохраняться очень долго и затем при благоприятных условиях прорастать.
Склероции развиваются следующим образом. Через повреждения в кожице или через укусы насекомых споры гриба попадают в мякоть плода и прорастают в ней в мицелий. На поверхности плода в это время обнаруживается небольшое бурое пятно, которое очень быстро разрастается и охватывает почти всю поверхность плода. Мякоть плода становится рыхлой, губчатой, теряет аромат и вкус. На поверхности плода в это время можно обнаружить в виде концентрических кругов органы бесполого размножения гриба. Мицелий первоначально распространяется в ткани более или менее равномерно, проходя через клетки растения-хозяина, а позже, к концу лета, начинает сосредотачиваться преимущественно в периферических частях плода. Поверхность плода приобретает черную окраску, становится блестящей, как бы лакированной и отвердевает, или, как говорят, мумифицируется. Мумифицированный плод может храниться в течение нескольких лет. В его образовании принимает участие гриб и ткань мякоти плода, пронизанная гифами гриба.
Образование такого типа склероциев происходит не только при поражении растительных тканей, но и при поражении животных. Так, например, при поражении шелковичного червя грибом из личинок этих насекомых получаются мумии, насквозь пропитанные грибницей. Следует подчеркнуть, что участие грибных тканей и тканей субстрата (хозяина) в образовании склероциев может быть различным. В приведенном выше случае мумификации яблока преобладающая масса склероция состоит из мякоти плода, из тканей самого плода. Но иногда бывает и наоборот, когда склероций-мумия образован преимущественно мицелием гриба. такой случай имеет место, например, у гриба, поражающего завязь различных злаков (рис.12.).Грибница здесь пронизывает завязь и превращает ее в удлиненное тело – рожок. В образовании принимают участие ткани завязи, но к моменту созревания склероция от них почти ничего не остается, и он оказывается плотностью состоящим из гиф гриба. Есть еще и третий тип склероциев – так называемые псевдосклероции или микросклероции. Они состоят из окрашенных толстостенных клеток, которые обычно образуются внутри тканей пораженных растений или на мицелии в условиях культуры.


Рис.12. Склероций на колосе ржи и его прорастание стромами.
Роль склероциев всех трех типов сводится к выживанию в неблагоприятных условиях, т.к. обычно грибы зимуют в виде склероциев. С наступлением благоприятной температуры и влажности мицелий во внутренней части склероция пробуждается к жизни. Он начинает использовать запасные вещества, находящиеся внутри склероция, пробивает оболочку, которая к тому времени становится эластичной, и прорастает чаще всего с образованием органов плодоношения.
Тема. ОСНОВЫ ФИТОЦЕНОЛОГИИ
Введение
Фитоценология – наука о фитоценозах. Эта наука еще называется геоботаникой. История этой науки начинается с трудов ботаников и географов 18 века, т.е. геоботаника зародилась в недрах ботанической географии, которая изучает закономерности географического распространения отдельных видов растений и растительных сообществ.
Отдельные положения этой науки мы видим в работах М. В. Ломоносова о происхождении черноземов. О фитоценозах знали еще К. Линней и М. Вильденов, последний ввел в науку термин ассоциация.
В России зарождение геоботаники как науки относится к 60 г.г. 19 века. К числу первых русских геоботаников относится академик Рупрехт Ф.И., а также сотрудник Киевского университета Борцов И.Г. Последний был членом экспедиции на берег Аральского моря, а в 1863 г. была издана работа «Материалы по ботанической географии Арало-Каспийского края». В работе не было еще терминов «сообщество» и «геоботаника». Позднее вышла работа Рупрехта «Геоботанические исследования о черноземе». Впервые в ней был введен термин «геоботаника». В 1863 году агроном Бажанов A.M. опубликовал статью «Об искусственно возделываемых лугах», где была изложена геоботаническая теория. Корелыциков С.П. опубликовал в 1865 году «Луговые и сорные травы», в которой рассмотрены взаимоотношения между луговыми травами, а также вопросы смен растительных сообществ.
Значительное развитие получила геоботаника в 80-х и, особенно,в 90-х г.г. 19 века и в начале 20 века.
Работы Коржинского С.И., Краснова Л.П., Танфильева Г.И. («Пределы лесов на юге России», «Пределы лесов в полярной России»), Сукачева В.П. («Растительные сообщества»), Пачоского И.К. («Стадии развития флоры»), Морозова Г.Ф. («Учение о лесе»), Докучаева В.В. («Русский чернозем», «Наши степи прежде и теперь», «К учению о зонах природы»), Раменского Л.Г (предложил использовать при изучении лугов методы массового учета и статобработки) получили всемирную известность.
В Казанском университете оформилась геоботаничекая школа под руководством Коржинского С.И. Долгое время этой школой руководил Гордягин А.Я., который изучал вместе с сотрудниками видовой состав сообществ Чувашии.
В отделе геоботаники ботанического института под руководством Кузнецова Н.И. была составлена первая карта в мире, охватившая очень большую территорию В 30-50 г.г. 20 века велись долгие научные дискуссии по многим вопросам: что такое ассоциация, что такое фитоценоз. В 30 годах вошел в употребление термин «фитоценология», второе название которой – геоботаника.
Геоботаника, фитоценология – наука описательная. Главная ее задача – изучение структуры растительных сообществ и ее изменяемости во времени и пространстве в зависимости от природных и антропогенных факторов.
Леса, степи, болота, пашни, сады, газоны – все это объекты фитоценотического исследования и поэтому знание фитоценологии позволяет управлять их продуктивностью и качеством накапливаемой биологической продукции (например, определение системы ухода и рубок лесов, допустимых норм и режима пастьбы скота, решение вопроса об оптимальных дозах удобрений естественных и сеяных лугов и т.д.). Фитоценолог рассматривает любые посевы, древостой или укосы луга не как механическую сумму растений, а как сложную систему, участники которой взаимодействуют друг с другом.
Современная фитоценология – наука с развитой методологией и достаточно мощным математическим аппаратом, который помогает исследователю выявлять закономерности, носящие статистический характер. В мире существуют десятки институтов и издается много журналов и монографий, где излагаются результаты фитоценологических исследований.
Понятие о фитоценозе, растительном покрове,
биоценозе, биогеоценозе
Растительное сообщество, или фитоценоз – это закономерное соотношение нескольких или многих видов растений на данном участке территории, находящихся в состоянии взаимозависимости как между собой, так и с окружающей средой.
Фитоценозы могут быть просто устроены, как, например, агрофитоценозы, где взаимные влияния изучены хорошо. Естественные фитоценозы устроены более сложно, здесь сложнее выяснить все взаимоотношения.
Совокупность фитоценозов образует растительный покров какой-либо территории.
Растительные сообщества тесно связаны с животным населением и микроорганизмами. Это сочетание образует биоценоз. Биоценозы находятся во взаимодействии с неживыми, или косными, элементами (горными породами, влагой, газовым составом атмосферы, а также почвой), образуя биогеоценоз.
Биоценозы образованы различными в трофическом отношении группами организмов: автотрофами и гетеротрофами. Автотрофы, или продуценты, способны на основе использования энергии солнечных лучей создавать органическое вещество (зеленые растения). Продуцентами являются также хемотрофы, которые используют энергию восстановления химических соединений. Гетеротрофы (животные, бактерии, актиномицеты, грибы, цветковые паразитные и сапротрофные растения) используют для своей жизни готовые органические вещества, созданные автотрофами, т.е. гетеротрофы не могут существовать без автотрофов. Особую группу организмов в составе биоценозов представляют азотофиксаторы.
Косная среда в наземных биоценозах подразделяется на почвенную и надземную среды: на эдафотоп и аэротоп.
В водных биоценозах различают гидротоп и аэротоп, или только гидротоп (если биоценоз состоит только из водных организмов).
Биоценоз – открытая система, являющаяся частью биосферы, в которой составные части ее находятся в постоянном взаимодействии друг с другом. Огромное воздействие на биоценоз оказывает человек. В результате его деятельности на месте одного биогеоценоза могут возникать другие (например, на месте леса – луг и т.д.).
Признаки фитоценоза
Набор видов растений, входящих в фитоценоз, называют флористическим составом, или флористической насыщенностью. Особенности жизненных форм этих видов и размещение в пространстве их тел (органов) создают структуру фитоценоза. Облик, внешний вид сообщества называют физиономичностью, или аспектом. Эти перечисленные признаки используются при изучении фитоценозов.
Повторяемость нахождения одних и тех же видов в разных участках фитоценоза называют константностью.
Высокая флористическая насыщенность и большое число константных видов свидетельствует об устойчивости фитоценоза (например, сныть в разных типах дубрав).
Виды в фитоценозах неравноценны, они делятся на ценоэлементы: эдификаторы и ассектаторы. Эдификаторы – это строители фитоценоза, они сильно влияют на среду (например, ель). Ассектаторы –- это соучастники, «наполнители» фитоценоза. В геоботанике часто пользуются другим понятием – доминанты. Под этим термином понимают виды, доминирующие в сообществе. Доминанты обычно являются эдификаторами. Но в то же время доминанты не всегда являются эдификаторами (ель – доминант и эдификатор; сосна - доминант, но не эдификатор).
В состав фитоценоза входят виды, относящиеся к различным жизненным формам. Раункиер различает следующие жизненные формы, которые он называет биологическими типами:
а) фанерофиты (почки возобновления, находятся на стеблях)
б) хамефиты (почки возобновления, находятся на побегах, которые лежат на поверхности земли)
в) гемикриптофиты (побеги отмирают)
г) криптофиты (почки под поверхностью почвы)
д) терофиты (зимуют в виде семян).
В состав фитоценоза входят виды, относящиеся к различным экологическим группам
- по отношению к свету (светолюбивые, тенелюбивые, теневыносливые, гелиофиты (сциофиты).
- по отношению к воде (гигрофиты, гидрофиты, ксерофиты, суккуленты, мезофиты, гелофиты - «амфибии»),
- по отношению к кислотности почвы (ацидофилы, базифилы, нейтрофилы),
- по отношению к богатству почвы (эутрофные, олиготрофные, мезотрофные),
- по отношению к солевому режиму (галофиты),
- по отношению к механическому составу субстрата (псаммофиты, литофиты),
- по отношению к металлоносным участкам (металлофиты).
При геоботанических исследованиях приходится иметь дело с группами особей одних и тех же видов, развивающихся в различной географической и экологической обстановке, в различных климатических условиях, в различных фитоценозах.
Естественная смесь особей одного вида называется популяцией. Каждая популяция имеет свой ареал. Наименьшая совокупность особей вида называется ценопопуляцией.
Совокупность однородных ценопопуляций одного и того же вида, называется экотипом. Среди биотических экотипов различают сенокосные, пастбищные, ценотические и т.д.
Ценотические экотипы, или фитоценотипы, – группы родственных популяций, сложившихся под влиянием фитоценотических условий, т.е. в пределах схожих фитоценозов.
Ценопопуляция состоит из неодинаковых в морфолого-экологическом отношении особей. Так, биотипы одуванчика различаются по окраске плодов, форме роста, опушению и т.д.
Как правило, виды сложены значительным числом битипов. Особи одного и того же вида, входящие в состав ценопопуляций, различаются по возрасту: латентный период (период первичного покоя) – семена, плоды; виргинильный (девственный) – период до образования цветков; генеративный – период размножения (семенами); сенильный (старческий) – период вегетации после размножения.
Пространственная структура фитоценоза
В фитоценозах принято различать элементы вертикальной и горизонтальной структуры. Вертикальное распределение в типичном ельнике таежной зоны, на лугу и в тропическом лесу различное. В ельнике четко различимы, по крайней мере, два яруса: древесный и кустарничково-травянистый. Иногда различают 4 яруса: деревья, кустарники, травы, мхи.
В то же время луговые сообщества, тропические леса часто не имеют разделения на ярусы (иногда же в тропических лесах выделяют 5-6 ярусов древесных пород). В таких сообществах характер распределения фитомассы от верхней границы до поверхности земли окажется более или менее постоянным. В этих случаях для описания таких сообществ пользуются условными горизонталями, горизонтальными слоями. Эти зоны (слои) могут различаться очень значительно. Распределение растений зависит от светообеспечения, вдоль градиента которого виды занимают свои экологические ниши, т.е. необходимые для существования вида условия.
В разных слоях фитоценоза обитают разные представители животного мира, что дает основание говорить о биогеоценогоризонтах.
Вертикальная дифференциация свойственна и подземным частям сообщества. 70-80% корней растений располагается в приземном слое почвы (не глубже 20-30 см), однако корни могут углубляться на несколько метров. Вертикальная дифференциация фитоценоза напоминает слоеный пирог.
Горизонтальное сложение фитоценоза подобно калейдоскопу с различными вариантами узора, т.е. это особенность размещения особей и их групп по горизонтали. Неравномерность размещения растений зависит от многих причин: неоднородности экотопа, особенностей вегетативного размещения особей и т.д. Если неоднородность состава охватывает участок сообщества во всех его ярусах, то в таких случаях говорится о микроассоциациях, а все сообщество, включающее несколько микроассоциаций называют комплексным.
Если же горизонтальная слоистость обнаруживается только в отдельных ярусах сообщества, то говорят о микрогруппировках, или микроценозах, или мозаичных пятнах. Например, клевер может формировать пятна диаметром 5-6 м. Вокруг пятен клевера разрастаются злаки, образуя густой темно-зеленый травостой. Здесь говорится о двух микрогруппировках, или мозаичных пятнах. В дубово-еловом лесу количество микрогруппировок бывает большим: елово-волосистоосоковое, елово-кисличная, елово-кру-пнопапоротниковая, дубово-снытевая, елово-костянично-паль-чатоосоковая, елово-щитовниковая, березово-елово-мертвопо-кровная.
При выделении фитоценотических горизонтов возникает вопрос: куда относить лианы,эпифиты? Лианы и эпифиты входят в состав тех горизонтов, к которым принадлежат части деревьев и кустарников, служащие им опорой.
Рассматривая структуру сообщества выделяют также синузии – группы видов, сходных по жизненным формам и экологическим свойствам. Каждая синузия занимает в сообществе свою экологическую нишу. Например, синузии мхов и лишайников, синузия травянистых растений «дубравного широкотравья».
Взаимоотношения между растениями в фитоценозе
Отношения между видами в фитоценозе многообразны. Симбиозом называют такие отношения между организмами, при которых один или оба организма извлекают из этих отношений пользу и ни один не испытывает ущерба (азотфиксирующие бактерии и цветковые растения). Первый тип отношений, когда оба организма извлекают пользу, называется мутуализмом, второй – когда пользу извлекает лишь один из организмов – комменсализмом (например, эпифиты на деревьях, лианы на деревьях).
Паразитизм – второй тип отношений. Один организм проводит более или менее длительное время на теле или в теле другого (омела, повилика).
Полупаразитизм. Один организм использует другой только как источник вещества, а растение – хозяин угнетается (марьянник луговой, зубчатка, погремок, очанка).
Антибиоз – один из организмов причиняет вред другому, например, выделением вредных веществ-колинов. Колины бывают газообразными (этилен, эфирные масла), жидкими, твердыми.
Конкуренция – борьба за пищу, за место (пиретрум вытесняет ковыль в степях).
Микориза – симбиотические отношения растения с грибом.
Средообразующие взаимоотношения
Они есть во всех фитоценозах, особенно наглядно проявляясь в лесах, где деревья создают затенение для кустарников, трав, мхов. Кроме того, в лесу всегда выражен свой специфический фитоклимат, отличный от фитоклимата луга, степи, болота.
Велико влияние на почву опадающей листвы или хвои, образующей на поверхности почвы подстилку, которая преобразуется в почвенный гумус (перегной). Величина опада в разных сообществах различна. Разные типы почв формируются под разными типами растительности; черноземы – под степями, светлокаштановые – под полупустынями, серобурые – под пустынями, подзолистые почвы – под хвойными лесами, бурые лесные почвы – под широколиственными, красные – под саваннами и т.д.
Фитоценозы могут снижать силу ветра (например, лесные массивы), уменьшают количество СО2 в дневное время суток; растения выделяют летучие вещества, которые могут убивать простейших, создавать специфический запах. Фитоценоз леса влияет на температуру воздуха и почвы, она бывает значительно ниже, чем на открытом месте.
Фитоценоз влияет на количество влаги и осадков: в еловом лесу большую часть дождя задерживает крона, а в буковом большая часть стекает по стволам и достигает земли. В то же время, количество осадков зависит от формы и строения крон, от возраста леса (в старом количество осадков, достигающих земли, увеличивается). Количество выпадающих осадков также больше над лесом, чем над открытым местом. Это связано с повышением влажности воздуха из-за транспирации воды деревьями.
Ценопопуляции также влияют на среду. Например, в негустой заросли хвоща влияние растений друг на друга довольно слабое. В густой смешанной заросли растения сильнее влияют друг на друга, в связи с чем виды, не способные к произрастанию в густой заросли, постепенно заменяются другими, более приспособленными. Меняется и характер грунта. Под хвощами был песок. По мере накопления перегнивших остатков появляется гумусовый горизонт почвы. Наносимые ветром и водой частички глины и пыли смешиваются с песком и образуют комочки почвы.
Изменчивость фитоценозов
Фитоценоз – явление динамичное. В нем непрерывно осуществляется сложная система взаимопереплетенных изменений. Поэтому фитоценоз меняет свой облик в зависимости от условий среды и этим самым способствует его устойчивости.
К числу признаков фитоценоза относится их изменение в пределах суток, года и от года к году. В соответствии с этим различают суточную, сезонную, разногодичную и возрастную изменчивость.
Суточная изменчивость проявляется, в основном, в повороте листьев, соцветий, складывании листьев на ночь, перераспределении планктонных водорослей в ночное и дневное время, суточном колебании массы органов растений, суточном ритме цветения (осоковые и ситниковые цветут утром) и т.д.
Сезонная изменчивость особенно ярко проявляется в широколиственных лесах, где напочвенный покров занят эфемероидами- подснежниками. Эти виды успевают отцвести до распускания листьев. Неодновременность зацветания цветков характерна для растений лугов и степей. В этом случае говорится о смене аспектов.
Знание сезонной ритмики луговых растений очень важно для организации сенокосного хозяйства. Ведь именно к моменту цветения в растениях наблюдается накопление питательных веществ (протеина) - важнейшего элемента корма.
Разногодичные изменения сообществ, обусловленные колебаниями различных факторов, называют флюктуациями. Причины флюктуации разные: климат года, характер разлива рек, срок становления пастбищ. Различают также фитопаразитарные и зооциклические флюктуации, например, массовое размножение полевок приводит к исчезновению ковылей и т.д. Разногодичные флюктуации выражены в пустынях, где в благоприятные годы наблюдаются вспышки растений-однолетников.
При флюктуации в сообществах многолетних растений флористический состав фитоценоза обычно не меняется, но изменяются размеры особей, число образуемых ими за сезон побегов и листьев, интенсивность вегетативного перемещения и размножения, семенная продуктивность.
Если же флюктуационные изменения захватывают и флористический состав сообщества, они могут привести к необратимым и стойким перестройкам в нем и приводят к смене его другим сообществом.
Смены фитоценозов. Под сменой понимают необратимые изменения, происходящие в фитоценозах, – сукцессии. Сукцессии подразделяются на кратковременные (частные), вековые (общие). Сукцессии вызываются эндогенными и экзогенными причинами.
Эндогенные сукцессии вызываются необратимыми изменениями внутренней среды самих фитоценозов при мало изменяющихся внешних условиях. Сообщества, в которых эндосук-цессионные процессы идут очень медленно и, следовательно, устойчивы, называют коренными, первичными. Они как бы завершают собой смены менее устойчивых сообществ, их называют еще климаксовыми, заключительными.
Неустойчивые сообщества, жизнь которых быстро приводит к сменам, называются вторичными (временными). Пример: при формировании растительного покрова на площади, не занятой растительностью, например, на свежих песчаных наносах реки, на отвалах породы, возле шахт, на насыпях, дамбах, а также при зарастании водоемов.
Экзогенные смены вызываются постепенными или резкими (катастрофы) внешними воздействиями на фитоценозы – климатическими, антропогенными. Скорость смен, вызываемых климатическими изменениями, очень мала, они протекают веками. Антропогенные воздействия протекают очень быстро. Пример частных (временных) смен – зарастание водоема. Существует 2 способа зарастания: со дна и с поверхности.
Дальнейшее развитие заросших водоемов путем зарастания со дна приводит или к образованию лесной растительности, или к образованию верхового болота. Развитие заросшего водоема с поверхности приводит после заполнения водоема илом к образованию болота, с «окнами», с трясиной.
Общие смены (вековые). Растительный покров любой страны претерпевал в течение тысячелетий сильные изменения. Например, если климат местности претерпевает иссушение, то леса сменяются пустынями. Таким образом, можно вообще говорить об истории фитоценозов. Эту историю можно изучать различными методами. Один из них – спорово-пыльцевой анализ слоев торфа или почвы, дающий представление о чередовании растительности в данной местности за ряд прошедших эпох. Другой метод – палеоботанический.
Формирование растительности
Многие вопросы до сих пор вызывают спор. Например, одни ученые считают степь первичной (она возникла после отступления ледника), которая затем сменяется лесом (Коржинский С.И., Танфильев Г.И.). Другие (Талиев В.И.) считают леса первичным типом, а степь – вторичным (т.к. кочевые племена выжигали лес).
Классификация фитоценозов
Основная единица в фитоценологии – ассоциация (Брюссельский конгресс, 1910 г.). Понимание ассоциации разными учеными разное. На конгрессе принято следующее толкование: «Ассоциация есть растительное сообщество определенного флористического состава с особыми условиями местообитания, особой физиономией». Сукачев дополняет «...ассоциация объединяет фитоценозы, характеризующиеся однородным составом, строением и в основном одинаковым сложением составляющих их синузий, сезонной и разногодичной динамикой».
Так, к одной ассоциации относятся все фитоценозы елового леса, образованные елью европейской с кислицей и зелеными мхами. Это ассоциация ельника-зеленомошника-кисличника.
Ассоциации объединяются в группы ассоциаций. К одной группе ассоциаций относят все ассоциации, различающиеся по составу одного из ярусов при тождестве основных особенностей остальных ярусов, в т.ч. одного главного образованного эдификатором. Так, к одной группе ассоциаций относят все ассоциации, где древостой образован елью европейской без кустарникового яруса с моховым ярусом из зеленых мхов. В нем господствуют в одних сообществах брусника, в других – черника, в третьих – кислица, в четвертых – его нет. Иными словами, к группе ассоциаций ельников зеленомошников относятся ассоциации ельник-зеленомошник-брусничник, ельник-зеленомошник-черничник, ельник-зеленомошник-кисличник, ельник-зеленомошник чистый.
Примеры групп ассоциаций для лугов: горные вейниковые луга с осокой, горные вейниковые луга с крупным разнотравьем, влажные разнотравные луга с купальницей, осоками и т.д.
Группы ассоциаций объединяют в формации. При выделении формаций должен быть общий эдификатор: формации ели европейской, формации ели сибирской, леса из черешчатого дуба.
Формации образуют группы формаций. Критерием для формирования группы является тип жизненных форм. Например: группы формаций темнохвойных лесов, светлохвойных листопадных, буковые леса, дубовые леса.
Для лугов: крупнозлаковые луга, мелкозлаковые, низкозлаковые.
Группы формаций образуют классы формаций. В этом случае к одному классу относят все группы формаций, относящиеся к разным жизненным формам: например, класс формаций хвойных лесов, летне-зеленые леса.
Для лугов: настоящие луга, остепненные луга.
Классы формаций формируют тип растительности: древесный тип, кустарниковый тип, травянистый тип.
5. ФОНДЫ КОНТРОЛЬНЫХ РАБОТ