
Варшавский В. И., Поспелов Д. А
| Вид материала | Документы |
СодержаниеГ л а в а 6 ДИАЛЕКТИКА ПРОСТОГО И СЛОЖНОГО В и Г. Для удобства будем обозначать такой автомат как А |
- Герберт Александер Саймон Исследователи ии: Лотфи Заде Исследователи ии: А. А ляпунов,, 9.34kb.
- «Как привлечь средства государственных институтов развития» Варшавский Владислав Римович, 48.54kb.
- Аннотация к научно-образовательному материалу, 114.81kb.
- 141. Поспелов В. И., Стальнов В. С. Содружественная аккомодация глаз при дисбинокулярной, 167.36kb.
- Тезисы докладов участников III международного конгресса «Россия и Польша: память империй, 1372.37kb.
- Д. А. Поспелов, Г. С. Осипов, 487.33kb.
- Варшавский А. С. Следы на дне, 1828.32kb.
- Рабочая программа учебной дисциплины Для направления 080100. 62 «Экономика» (программа, 562.39kb.
- Диспут с Пирром: прп. Максим Исповедник и христологические споры VII столетия / Отв, 73.89kb.
- Программа дисциплины: Имитационные модели для направления Прикладная математика и информатика, 120.53kb.
Именно так называл свое научное выступление на одной из школ по коллективным моделям поведения известный советский кибернетик М. М. Бонгард. В этом выступлении он говорил о том, что излишняя централизация в биологических организмах может нанести огромный вред. При возрастании централизации организм все большие ресурсы будет затрачивать на обработку информации для принятия решений, ему будет оставаться все меньше времени на поисковую и адаптационную деятельность. И М. М. Бонгард привел в качестве примера адептов учения йогов, которые в своей практике часто достигают того, что «вытаскивают наверх, в сознание» управление теми физиологическими процессами, которые протекают у человека на уровне автономных и полуавтономных систем управления. Они, например, могут сознательно регулировать ритм биения
173
сердца, сокращать и расслаблять желудок, сознательно управлять температурой тела и т. п. Но к чему это приводит? В пределе, когда все автоматизмы подавлены, йог должен тратить все свое время и ресурсы мозга на то, чтобы все эти процессы протекали без срывов, иначе жизнь его может оказаться под угрозой. Но тогда ему уже не хватит времени ни на что другое, ни на размышления, ни на созерцание. Конечно, индийские йоги не попадают в подобное положение. Автоматизмы они сохраняют. И вмешиваются в течение физиологических процессов лишь изредка. Да и цель их иная. В овладении секретом управления автономными процессами, забота о которых вытеснена из сферы сознания, они видят еще одну ступень в овладении законами управления своим телом. Но аналогия, подмеченная М. М. Бонгардом, очень ярка и поучительна.
Мы много говорили о параллельных процессах и методах их взаимодействия. В' человеческом организме формы этого взаимодействия куда богаче. Но суть явления сохраняется. Процессы текут почти автономно, синхронизуясь во времени за счет редких периодических или специфически определяемых ситуацией сигналов.
Однако децентрализация, при которой подсистемы работают практически автономно, обладает одним весьма существенным недостатком, о чем мы еще не говорили, хотя читатели могли бы и сами догадаться о нем. По крайней мере во многих наших моделях, функционирующих в быстро меняющихся средах, он был явно заметен. Этот недостаток связан с тем, что за децентрализацию управления приходится платить увеличением времени адаптации. То, что по единому приказу из центра можно сделать в системе за весьма короткое время, если центральное звено заблаговременно получит информацию об изменениях свойств среды, в децентрализованной системе будет осуществляться весьма медленно. Наверное, поэтому в биологических организмах (и у человека, в частности) имеются как бы два уровня: децентрализованный и централизованный по управлению. Однако эти уровни не дублируют друг друга.
Пока окружающая среда почти неизменна и вполне устраивает человека, децентрализованное управ-
174
ление реализуется в полном объеме. Отдельные его подсистемы функционируют автономно и почти не взаимодействуют между собой. Но вот произошло резкое изменение состояния среды, грозящее человеку неприятными последствиями. Требуется как можно быстрее перевести все подсистемы в состояние «боевой готовности». И тогда срабатывает централизованное управление, переводящее организм в состояние, которое можно назвать ситуацией стресса. Основная особенность этой реакции — ее неспецифичность. Она осуществляется в любых опасных ситуациях и направлена на взаимодействие со всеми подсистемами организма. В' кровь начинают выделяться гормоны, стимулирующие адаптационные реакции, повышается готовность организма к отдаче энергии, подпитываются мышцы и т. п. После этого либо наступает период адаптации, либо стрессовая ситуация исчезает. В наихудшем случае организм так долго стоит в готовности номер один, что наступает истощение, а, возможно, и гибель.
Таким образом, между децентрализованной и централизованной частями системы управления мы наблюдаем весьма интересное распределение функций. В медленно меняющихся или неизменных средах децентрализованная часть системы управления успешно справляется с адаптацией поведения к среде и достижением глобальных целей организма, а при резких изменениях среды организм включает некоторую систему всеобщего назначения.
Специалисты по управлению интегральными роботами (в отличие от узкоспециализированных роботов. последние должны действовать в широком классе сред, точное описание которых сделать невозможно) сейчас находятся в весьма нелегком положении. С одной стороны, совершенно очевидно, что в роботе имеется немало подсистем, которые должны функционировать автономно или почти автономно, получая сигналы из центрального блока управления (например, подсистемы «глаз» и «рука», позволяющие роботу найти нужный предмет, взять его и совершить с ним какую-либо операцию, должны действовать параллельно и автономно, согласуя свои действия лишь не слишком частыми синхронизующими сигналами). С другой стороны, возникает проблема создания неспецифических глобальных видов воздействий
175
от центрального блока, способного обеспечить целесообразное поведение робота. Общие законы такого поведения сформулировать очень трудно. Вспомним, например, три общих закона робототехники, предложенные в свое время А. Азимовым. Эти законы находятся в приоритетной связи. Первый из них самый приоритетный. Согласно ему робот никогда, ни при каких обстоятельствах не должен причинять вред человеку. Это закон всеобщего запрещения. И довольно легко себе представить, как можно организовать воздействие на подсистемы при опасности нарушения этого закона. Второй закон А. Азимова говорит о том, что робот всегда должен стремиться к достижению поставленной перед ним задачи, если это не противоречит первому закону. А третий закон робототехники указывает роботу на необходимость принимать все меры к самосохранению, если это не противоречит предшествующим двум законам. Но два последних закона уже не могут быть неспецифичными в отношении сигналов, передаваемых подсистемам робота. Требуется их спецификация по типам целей, которые ставятся перед роботом, и способам его самоохранительных действий.
Неспецифические сигналы централизованной части управляющей системы в наших многочисленных автоматных и неавтоматных моделях поведения были представлены различными воздействиями среды на подсистемы. Такие механизмы, как введение общей кассы или случайных парных взаимодействий, играют в этих моделях общерегулирующую роль. Напомним читателю, что, как мы говорили в § 4.4, целью коллектива может быть не только достижение целесообразного (или оптимального) поведения во внешней среде, но и поиск этих регулирующих воздействий, позволяющих подсистемам прийти к некоторому согласованному функционированию.
Для того чтобы еще раз подчеркнуть весьма важную для нас мысль о вреде «вытаскивания» специфических функций в централизованную часть системы управления, мы закончим этот параграф одной сценкой, которую можно было наблюдать на международной конференции по проблемам искусственного интеллекта и робототехники. Один из высокопоставленных представителей военно-морского флота США в ответ на жалобы докладчика о том, что весьма
176
трудно придумать небольшое число неспецифических законов целесообразного поведения для интегральных роботов, сказал, что он не видит в этом особой проблемы. И пояснил свою мысль следующим примером из жизни. Когда новички попадают на корабль, то первое время они никак не могут приспособиться к новой среде, совершают массу ошибок и вместо помощи часто наносят непоправимый вред. Команда вынуждена тратить силы на то, чтобы следить за новичками и оберегать их от беды. Однако всего этого можно избежать. Новичкам достаточно усвоить раз и навсегда, на весь их начальный период адаптации на корабле, только один неспецифический закон: «Если ты видишь движущийся предмет, то отдай честь, если же предмет неподвижен, то покрась его».
Это, конечно, анекдот, но мысль, высказанная в нем, точно отражает нашу проблему. Однако, к сожалению, сегодня мы слишком мало знаем о том, как надо строить эти регулирующие процедуры в системах децентрализованного управления,
Г л а в а 6 ДИАЛЕКТИКА ПРОСТОГО И СЛОЖНОГО
«Дороги, которые мы выбираем, следует отличать от дорог, которые выбирают нас».
Феликс Кривин
§ 6.1. Синтезогенез и интеграция усилий
«Все эти создания обладали тройственной симметрией и напоминали формой греческую букву гамма с тремя остроконечными плечиками, соединяющимися в центральном утолщении. В падающем свете они казались черными, как уголь, в отраженном — переливались синим и оливковым цветом, как брюшки некоторых земных насекомых. Наружные их стенки состояли из очень мелких пластин, напоминающих грани бриллианта, а внутри «мушки» содержали одну и ту же микроскопическую конструкцию. Ее элементы, в сотни раз меньшие, чем зернышки песка, образовывали что-то вроде автономной нервной системы, в которой удалось различить две частично независимые друг от друга цепи.
Меньшая часть, занимающая внутренность плеч, представляла собой микроскопическую схему, заведующую движением «насекомого», нечто вроде универсального аккумулятора и одновременно трансформатора энергии. В зависимости от способа, каким сжимали кристаллы, они создавали то электрическое, то магнитное поле, то переменные силовые поля, которые могли нагревать до относительно высокой температуры центральную часть; тогда накопленное тепло излучалось наружу однонаправленно. Вызванное этим движение воздуха, .как реактивная струя, делало возможным движение в любом направлении. Отдельный кристаллик не столько летал, сколько подпрыгивал, и не был, во всяком случае во время лабораторных экспериментов, способен точно управлять своим полетом. Несколько же кристалликов, соединяясь кончиками плеч друг с другом, образовывали систему с тем лучшими аэродинамическими показателями, чем больше их было.
173
Каждый кристаллик соединялся с тремя; кроме того, он мог соединяться концом плеча с центральной частью любого другого, что давало возможность образования многослойных комплексов. Соединения не обязательно требовали соприкосновения, кристалликам достаточно было сблизиться, чтобы возникшее магнитное поле удерживало все образование в равновесии. При определенном количестве насекомых система начинала проявлять многочисленные закономерности, могла в зависимости от того, как ее «дразнили» внешними импульсами, менять направление движения, форму, вид, частоту внутренних пульсаций; при определенных внешних условиях менялись знаки поля, и, вместо того, чтобы притягиваться, металлические кристаллики отталкивались, переходили в состояние «индивидуальной россыпи».
Эта длинная цитата из повести Станислава Лема «Непобедимый» приведена нами не случайно. На планете «Регис-III» люди столкнулись с необычным явлением. Из примитивных кристалликов, обладающих примитивным поведением, при определенных условиях возникал сверхорганизм— туча. И эта туча обладала почти неисчерпаемыми возможностями по адаптации своего поведения, ибо хранила в огромной памяти, складывающейся из памятей-песчинок отдельных кристаллов, необъятный запас знаний.
Однако столь ли уж необычен этот способ возникновения сложного из простого? После того, что читатель прочитал в предшествующих главах, подобный путь организации сложного поведения должен казаться ему весьма привычным; Наблюдения за биологическими организмами также не противоречат идее польского фантаста. Такое объединение более простых организмов в более сложный —один из путей эволюции в органическом мире. К. М. Завадский, много лет занимавшийся проблемами эволюции, назвал такой путь синтезогенезом. Переход от одноклеточных водорослей к многоклеточным был решающим шагом на пути прогресса органического мира;
сообщество рабочих пчел в улье или рабочих муравьев в муравейнике—примеры того же типа.
Но простое скопление однородных подсистем или организмов—это еще не новая система или организм. Множество рабочих пчел, встретившихся на цветущем лугу и относящихся к разным пчелиным
179
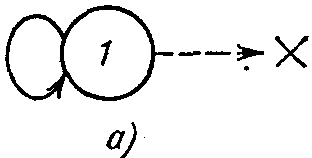
семьям,—это совсем не то, что множество рабочих пчел из одного улья. И совокупность пассажиров, оказавшихся одновременно в трамвае, резко отличается от множества покупателей и продавцов на колхозном рынке.
В чем же состоит это отличие? В самом общем виде можно сказать, что некоторая совокупность элементов является единой системой, если эти элементы обладают потенциальным свойством образовывать статические или динамические структуры, необходимые для «выживания» элементов и всей их совокупности, т. е. обладают свойством устанавливать взаимодействие друг с другом для достижения локальных и глобальной целей. Это, конечно, не определение, а скорее рассуждение о чрезвычайно сложном вопросе. Исчерпывающий ответ на него — предмет специального исследования, выходящего далеко за границы возможностей авторов. Но, как нам кажется, суть всех моделей коллективного поведения и взаимодействия в этом и состоит. Отметим еще, что когда речь идет о биологических совокупностях, то в реальных ситуациях эти потенциальные свойства проявляются лишь частично, а остальные — ждут своего часа. Хорошо- известны, например, опыты с некоторыми бактериями, которые всегда обитали в средах, где отсутствуют определенные виды углеводов. При искусственной пересадке их в среды, где эти непривычные углеводы были единственной доступной для бактерий пищей, они начинали вырабатывать фермент для их расщепления. Возможность этого была заложена в их генную структуру «на всякий случай» и реализовалась именно тогда, когда в этом возникла необходимость. Другой пример — огромные потенциальные возможности любого человека, подавляющее большинство которых никогда не проявляется у индивида, а возможно, и у человеческого сообщества.
Таким образом, синтезогенез—это путь увеличения числа потенциально возможных свойств, которые могут пригодиться системе при встрече с непривычными для нее ситуациями и средами.
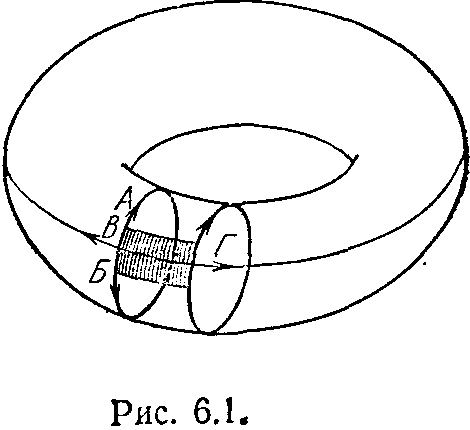
Рассмотрим простую модель, иллюстрирующую возможности синтезогенеза. На рис. 6.1 показан тороидальный мир — совокупность клеток, размещенных на внешней поверхности тора (обычная сушка
1
80
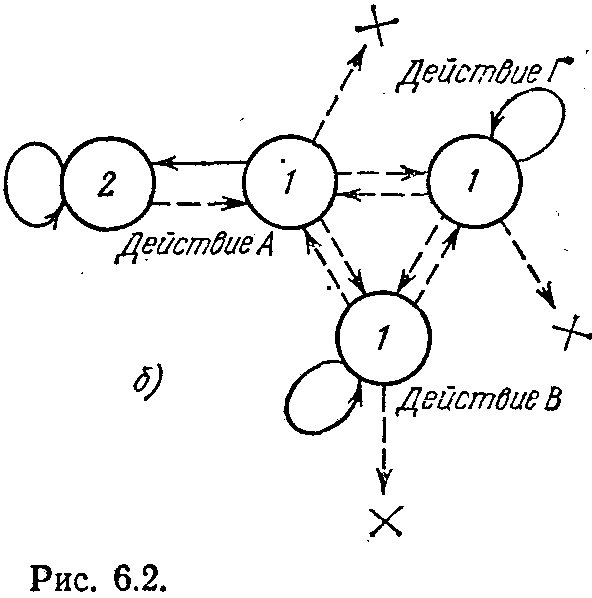
или баранка дают превосходное представление о тороидальной форме). Предположим, что в клетках этого мира может находиться пища, которой могут питаться «организмы», обитающие в них. В качестве таких «организмов» будем рассматривать автоматы с линейной тактикой. Простейшая форма подобного автомата — автомат с одним действием, показанный на рис. 6.2, а. В состоянии 1 при получении сигнала штраф автомат «умирает» (на рисунке это отмечено крестиком). Действие, которое может совершать автомат,— перемещение в некотором фиксированном направлении на одну клетку тора. Обозначим четыре возможных направления перемещения, показанные на рис. 6.1, через А, Б, В, Г. Тогда простейшие автоматы будут делиться на четыре типа — будем обозначать их теми же буквами. Допустим, что автоматы, находящиеся в одной клетке, могут объединяться. Если объединяются два автомата одного типа, то это приводит к увеличению длины лепестка (т. е. глубины памяти для этого действия). При объединении же автоматов различного типа новый автомат имеет уже не один лепесток, а два. На рис. 6.2,6 показан автомат, который возник в результате объединения четырех автоматов, два из которых относятся к
181
типу А, а оставшиеся два — к типу В и Г. Для удобства будем обозначать такой автомат как А2ГВ.
В отличие от классического автомата с линейной тактикой наш автомат не может накапливать наказания безгранично и «умирает», когда число подряд действующих штрафов (пунктирные стрелки) превышает число состояний, имеющееся у автомата (для автомата, показанного на рис. 6.2, б, оно равно четырем). Кроме того, смена лепестков происходит равновероятно.
Сигналы наказания и поощрения формируются средой следующим образом. Если автомат в данной клетке съедает пищу, то он получает сигнал поощрения, в противном случае—сигнал наказания. После того как автомат съест пищу (на что в модели требуется один такт) и уйдет из клетки, то пища может в ней одномоментно восстановиться или клетка останется пустой до того момента, когда по закону, характеризующему среду, пища снова восстановится.
Если в одну и ту же клетку попадает несколько автоматов, то они принудительно объединяются и образуют новый более сложный «организм».
Рассмотрим несколько ситуаций в эволюционном процессе на торе.
Н
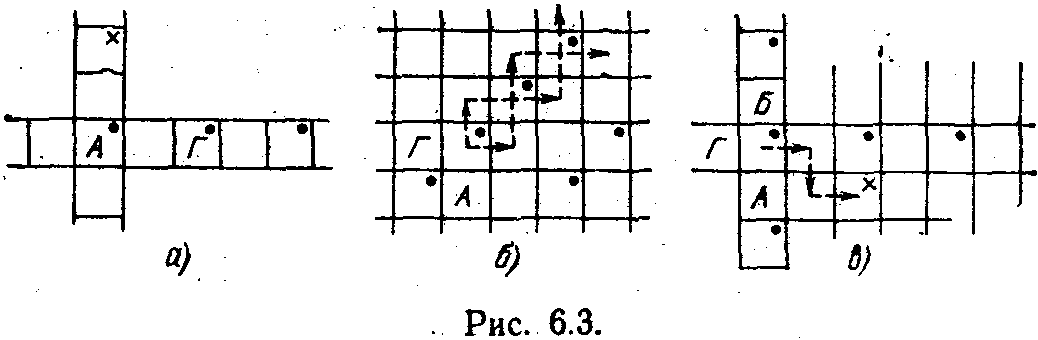
а рис. 6.3 показано несколько простейших ситуация на некотором участке тороидальной поверхности. Клетки, в которых имеется пища, отмечены точками. Предполагается, что пища, съеденная в клетках, полностью восстанавливается, как только автомат уйдет из нее. На рис. 6.3, а показаны два простейших автомата. Автомат Л съедает пищу в клетке, где он находится, и идет наверх. Но на этом кольце пищи нигде больше нет. В результате он погибает в клетке, помеченной крестиком. Иная судьба у автомата Г. Если пища имеется на всем кольце,
182
то этот автомат, двигаясь по замкнутому кольцу вправо, будет все время поддерживать свое существование. Он живет вечно, не беспокоясь ни о чем.
На рис. 6.2, б показана еще одна очень простая ситуация. Автоматы А и Г встречаются в клетке с пищей, объединяются и начинают движение. Каким оно будет? Это зависит от того, какое именно состояние окажется начальным. Ели это состояние, соответствующее состоянию 1 автомата А, то объединенный автомат сначала сделает шаг наверх. В этой клетке пищи нет и автомат получит наказание. Это заставит перейти его в состояние 1 бывшего автомата Г и сделать шаг на одну клетку вправо. Там пища есть. Съев ее, автомат сделает еще один шаг вправо. Получив наказание, он, как автомат А, сделает шаг вверх и получит пищу. Далее процесс будет повторяться циклически, если пища размещается на поверхности тора регулярным образом. Автомат будет двигаться по «диагональной линии» и жить вечное Если бы начальным состоянием объединенного автомата было состояние 1 автомата Г, то движение было бы аналогичным. Пунктирные стрелки показывают оба возможных пути автомата АГ.
Усложнение структуры далеко не всегда приводит к улучшению функционирования. Это положение иллюстрируется рис. 6.3, в. В клетке с пищей образуется автомат АБГ. Пусть начальным его состоянием является состояние 1 автомата Г. Сдвинувшись на одну клетку вправо и получив сигнал штраф, автомат переходит (путем равновероятного выбора) в состояние автомата Б. Он сдвигается вниз, но пищи там нет. Опять следует равновероятный переход, и автомат снова попадает в состояние автомата Г. Происходит сдвиг вправо. Но так как пищи в этой клетке нет, объединенный автомат погибает, исчерпав все свои ресурсы. Если бы объединения не произошло, то при том распределении пищи, которое показано на рис. 6.3, в, все три простейших автомата могли бы жить вечно.
Наши забавные автоматы на тороидальной поверхности можно исследовать с разных точек зрения. Но, к сожалению, это увело бы нас весьма далеко от основной канвы книги. Те, кому понравился этот мир, могут придумать много занимательных и интересных историй, полных драматизма и неожидан-
ных метаморфоз, которые могут развернуться на поверхности тора.
Для нас же важно отметить, что синтезогенез может приносить как пользу, так и вред, ибо иногда лучшее — враг хорошего.
Тем не менее путь синтеза, своеобразной полимеризации, часто встречается в эволюционирующих технических системах. Этот путь сыграл большую роль в создании мировой сети связи и транспортных сетей. При образовании комплексов резервированных устройств мы также сталкиваемся с явлением, подобным синтезогенезу.
Выскажем еще раз одну весьма важную мысль, связанную с синтезогенезом. В процессе такого объединения возникает особое явление, сходное (чисто внешне) с полимеризацией в химии. Элементы, вступая в объединение и не меняясь по своей структуре, как бы приобретают новые качественные возможности. И эти новые возможности зависят от механизма объединения. В гл. 4 мы уже столкнулись с этим явлением. Когда два автомата объединялись чисто механически (так, как объединяются автоматы в нашей модели эволюции на горе), число их состояний растет, как п2, если каждый из автоматов имел п состояний. Когда же они объединяются за счет случайного парного взаимодействия, то это дает им возможность функционировать как автоматам, обладающим памятью глубины 2n. В гл. 5 мы также столкнулись с явлением «полимеризации». Автомат всего с восемью состояниями, объединившись в шеренгу стрелков, как бы приобретал возможность работы с памятью всей совокупности автоматов, становился богаче по своим возможностям, не меняя своей структуры. Это явление кажется нам весьма любопытным.
Но, наряду с этим процессом в биологической и технической эволюции, идет и другой важный процесс, связанный с ростом неоднородности в организме с появлением специализированных подсистем.
§ 6.2. Сегрегациогенез и его последствия
Термин сегрегациогенез, как и термин синтезогенез, принадлежит К. М. Завадскому. Его смысл сводится к тому, что в процессе развития биологических
особей идет не только их усложнение путем объеди-нения более простых организмов в более сложные, но и процесс дифференциации функций, выполняемых отдельными подсистемами, и ведущий затем к изменению структуры этих подсистем для лучшего осуществления своих специфических функций. Прогресс требует отказа от универсальности, однотипности. Универсальный элемент делает все одинаково плохо. Если пища на торе в примере, рассмотренном в предыдущем параграфе, всегда расположена так, что для обхода клеток с пищей нужен ход шахматным конем, то имеет смысл, чтобы специфическая функция автомата позволяла бы ему прямо выполнять это движение в течение одного такта. Но если пища расположена иным образом, то подобное действие и не нужно.
Коллизия между универсальностью и специфичностью, между однородностью и разнородностью есть явление всеобщее, встречающееся всюду. Биоценозы и техноценозы также демонстрируют эту коллизию.
Количество рабочих пчел в улье может колебаться в довольно широких пределах, и они образуют подсистему, способную прожить самостоятельно, но самка пчелиного улья должна быть всегда одной единственной, и она быстро погибнет, если лишить ее рабочих пчел. Дифференциация здесь зашла уже весьма далеко, и отдельные подсистемы перестали уже быть способными к автономному функционированию вне той системы, в состав которой они входят.
Однако польза от появления подобных подсистем очевидна. Мы уже говорили в гл. 3 о пользе разнородности в коллективе автоматов. Ранги рефлексии, уровни пессимизма—оптимизма были первыми показателями различий, намечавшихся в подсистемах, которые позволяли неоднородному коллективу более успешно решать стоящую перед ним задачу, чем однородному коллективу. Правда, любой из автоматов такого коллектива мог бы функционировать и в одиночку. Но это просто означает, что специализация еще не дошла до того рубежа, за которым самостоятельное существование отдельной подсистемы становится невозможным. Однако специализация — непременный спутник прогресса, ибо только с ее помощью
185
можно уменьшать затраты времени на достижение тех или иных целей, стоящих перед организмом.
В качестве иллюстрации сказанного рассмотрим, например, эволюцию в области ЭВМ. На первом этапе каждая вычислительная машина представляла собой некоторое единое и неделимое целое. Ее процессор, память, устройства обмена с внешним миром и управляющая система находились в столь жестких связях, что не могли не только функционировать, но и анализироваться отдельно друг от друга. Все процессы в ЭВМ протекали строго последовательно под контролем центрального устройства управления. Такую ЭВМ мы можем уподобить некоторой «клетке» в мире вычислительной техники.
Как же происходила эволюция ЭВМ в последующие годы? Один путь был связан с усложнением структуры ЭВМ, введением в ее состав новых подсистем, обеспечивающих для нее возможность выполнения новых функций*) (например, графопостроителей, которые сделали возможным выводить из ЭВМ не только текстовую, но и графическую информацию, или появление в составе ЭВМ датчиков случайных чисел, позволяющих использовать при решении задач методы, опирающиеся на случайные распределения). Эти подсистемы, увеличивая сложность «клетки», не меняли принципиально условий ее существования. Но это усложнение вело к усложнению управляющей системы, на плечи которой падало все больше задач. И наступил момент, когда операционные системы ЭВМ (а именно они, как правило, выполняют роль центрального блока управления всеми процессами, протекающими в ЭВМ) стали самым узким местом. Появились грозные признаки того, что усложнение структуры ЭВМ приведет в тупик. Все чаще и чаще возникали ситуации, которые специалисты по операционным системам называют дедлоками (ловушками). Это такие состояния, когда требования различных процессов, протекающих в машине, предъявляют к операционной системе разноречивые требования, и она не знает, что ей делать.
*) Эти подсистемы могут быть реализованы и в виде программ.
186
Стало ясно, что при централизованном управлении дальнейшее усложнение структуры ЭВМ и улучшение ее функционирования уже невозможно.
П

ереход к комплексированию ЭВМ был тем следующим шагом в эволюции, который надо было неизбежно сделать. Синтезогенез сработал. Вместо «одноклеточного» вычислительного устройства появились «многоклеточные». Эти образования могли иметь различную структуру. На рис. 6.4 показаны некоторые типы структур комплексов ЭВМ. На рис. 6.4, а приведена структура с центральной ЭВМ 1, которая выполняет роль центрального управляющего устройства для ЭВМ 2, 3, 4, на рис. 6.4,6 мы видим смешанную структуру, а на рис. 6.4, в — децентрализованную, в которой все ЭВМ равноправны. Важно отметить, что даже в централизованной структуре возникает некоторая децентрализация. Центральная ЭВМ не все время ведет процессы в подчиненных ей машинах. Она лишь инициирует в них начало некоторых процессов, синхронизует протекающие процессы между собой и производит обмен информацией между процессами. А в остальном машины, входящие в систему, действуют самостоятельно. И это направление эволюции подтверждает правильность отказа от пути йогов, о котором говорилось в гл. 5.
187
Интересно отметить, что децентрализованная структура, показанная на рис. 6.4,в, демонстрирует возможность введения в структуру «организма» неспецифического централизованного управления. Показанный на этом рисунке пунктиром блок синхронизации К. может по специальной кольцевой шине передавать сигнал одновременно всем ЭВМ, образующим систему. Это может быть, например, сигнал прерывания всех вычислений для приема новой внешней информации, или для повторения вычислений, или для тестовой проверки. Но такой центральный управляющий блок может и отсутствовать. Тогда синхронизация работы децентрализованной системы будет осуществляться по типу кольца стрелков, о чем было рассказано в гл. 5.
Кроме этого основного пути эволюции ЭВМ приблизительно в те же годы развивался и еще один путь — создание ЭВМ на основе однородных клеточных структур, о которых мы также говорили в гл. 5. Этот путь был связан с идеей синтезогенеза в чистом виде. Предполагалось, что однородность и универсальность отдельных подсистем (автоматов, находящихся в клетках однородной структуры с потенциально однотипными связями между ними) позволят улучшить характеристики ЭВМ. Однако этого не произошло. Ибо сегрегациогенез оказался куда более эффективным в отношении этих характеристик.
И следующий шаг в эволюции ЭВМ — комплексирование не однотипных, а узкоспециализированных подсистем, причем для каждой из них четко определены те функции, которые она реализует. Сначала это привело к структурам того же типа, что и показанные на рис. 6.4. Отличие состояло лишь в том, что ЭВМ, входящие в систему, стали специализированными. Например, они могли быть специально созданы для обработки символьной информации, работы с матрицами, предварительной обработки и преобразования сигналов, поступающих от объекта управления, и т. п. Но при этом, как в автоматных моделях с рефлексией или уровнями пессимизма — оптимизма, все такие ЭВМ могли действовать и вне системы, автономно.
При дальнейшем сегрегациогенезе это свойство исчезло. Дифференциация коснулась даже той исходной ячейки — ЭВМ, которую мы уподобили клетке,
188
Е
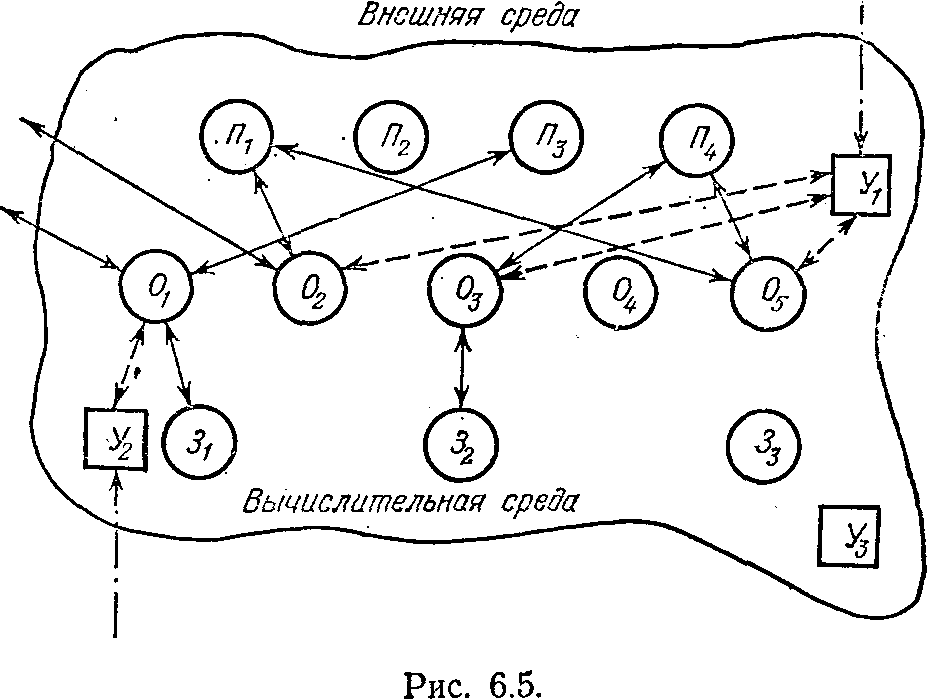
е составляющие как бы обрели самостоятельность, и возникла структура, показанная на рис. 6.5. Процессоры, блоки памяти, блоки обмена и управляющие блоки как бы плавают в некоторой вычислительной среде. Их объединение в структуру происходит динамически, управляющие блоки, получив задание, ищут исполнителей, свободных от работы, и организуют процесс. На рис. 6.5 показан такой момент, когда управляющий блок У1 объединил для решения задачи два процессора П1 и П4, один блок памяти 32 и три блока обмена O1, О3 и О5. Одновременно управляющий блок У2 организовал другой процесс, объединив для этого в структуру процессор Пз, запоминающее устройство 31 и обменное устройство O1. Задачи управляющие блоки получают из внешней среды. Из той же среды обменные устройства получают исходную информацию. Результаты решения также возвращаются во внешнюю среду. После окончания решения задачи структуры «рассыпаются».
В этой структуре сегрегациогенез зашел настолько далеко, что отдельные подсистемы автономно не могут существовать. Лишь объединившись в структуру, где обязательно наличие одного управляющего блока и хотя бы одного обменного устройства, связанного с процессорами или запоминающими устройствами, наш «организм» сможет функционировать. Способность образовывать структуры под задачи
189
демонстрирует его адаптационные возможности, а специализация отдельных подсистем позволяет реализовать связанные с ними функции параллельно и максимально быстро.
А
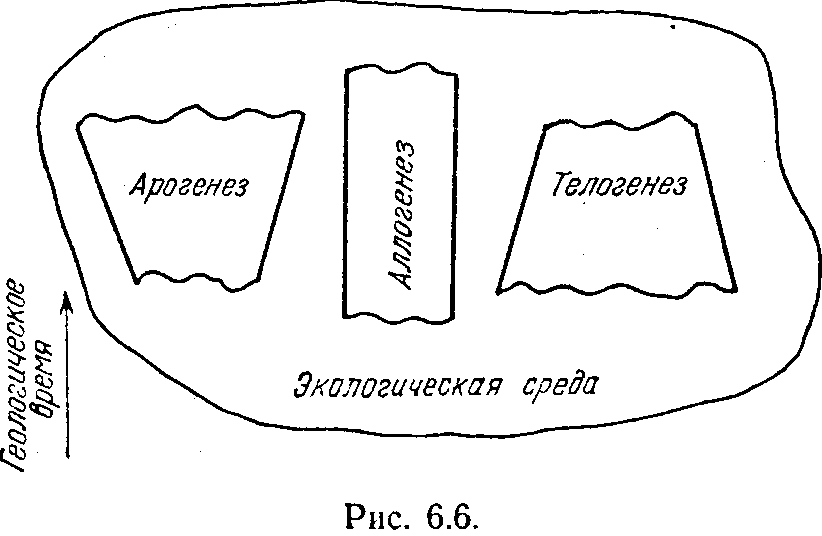
нализируя прогресс в эволюции, К. М. Завадский предложил следующую наглядную схему, показанную на рис. 6.6. Есть как бы три возможности в эволюционном развитии биологических организмов. При первом из них арогенезе идет расширение адаптационных возможностей организма. Он как бы расширяет набор сред, в которых он будет выживать и давать потомство. Этот процесс может идти либо за счет синтезогенеза (как в нашей модели эволюции на тороидальной поверхности), либо за счет сегрега-циогенеза (как, например, в вычислительной среде, дающей возможность решать любые задачи, для которых у системы хватает ресурсов). Заметим, что при наличии тех же ресурсов в рамках единой системы типа, показанной на рис. 6.4, а, не удалось бы, например, организовать одновременное протекание двух процессов, показанных на рис. 6.5.
Если арогенез есть расширение адаптационных возможностей системы, то аллогенез есть смена некоторых функций, реализуемых организмом, на новые, экологически равноценные. Другими словами, при аллогенезе происходит как бы смена одной экологической ниши на другую, более выгодную для выживаемости организма. Такое явление можно наблюдать не только в биологии, но и в технике. В эпоху клавишных вычислителей, предшествующую появле-
190
нию ЭВМ, они использовались в основном в научных расчетных бюро. ЭВМ вытеснили их оттуда, но они нашли свою экологическую нишу в бюро технико-экономических расчетов, которым невыгодно пользоваться услугами ЭВМ. Самолеты в свое время вытеснили дирижабли, но похоже, что горячие поклонники дирижаблей нашли для них новую подходящую нишу в современном техноценозе, и в ближайшее время мы, возможно, вновь увидим в небе их неповторимые силуэты.
Наконец, телогенез — это как бы обратная сторона арогенеза. При телогенезе происходит очень глубокая адаптация к заданному состоянию экологической среды, которая достигается глубокой специализацией организма. Примеры телогенеза в технических системах очевидны. Практически все узкоспециализированные системы могут рассматриваться с этой точки зрения. Первобытное рубило, пригодное для всех случаев жизни, постепенно породило огромное количество рубящих инструментов, многие из которых пригодны для выполнения очень конкретных работ, но не могут использоваться для чего-либо иного (например, колун, если только не использовать его обуха для забивания чего-либо).
Арогенез, аллогенез и телогенез — это не альтернативные пути эволюции. Они действуют согласованно и одновременно. Доминирование любого из них может оказаться в развитии некоторого организма временным и преходящим. Но все эти пути направлены на единственную цель — улучшение адаптации .организма к данной среде и, как следствие этого, .увеличение его выживаемости в ней. И сказанное .вполне можно перенести на технические системы.. .