
Новых пород свиней на полигибридной основе
| Вид материала | Реферат |
- Вопросы, 46.61kb.
- Определение свойств горных пород и оценка сопротивляемости горных пород разрушению, 79.2kb.
- Выращивание свиней по канадской технологии, 101.01kb.
- Задачи селекции : повышение урожайности сортов культурных растений, увеличение продуктивности, 105.58kb.
- - наука о методах создания и улучшения пород животных, сортов растений, штаммов микроорганизмов, 68.13kb.
- С. Чернышевка, ул, 380.66kb.
- Организация воспроизводства стада свиней, 561.55kb.
- Увага! Африканська чума свиней!, 29.28kb.
- Продуктивные качества свиней крупной белой породы с использованием генотипов пород, 719.89kb.
- Зачет по теме: «Селекция», 41.19kb.
Условные обозначения: 1. КБ – крупная белая порода; Л - ландрас; ПБ – полтавско-белорусский мясной тип; ПХ – полтавско-харьковский мясной тип. 2. Равно числу скрещиваний в процессе создания породной комбинации типа.
Пояснения к таблице 1. При определении числа пород в группах встречав-шиеся в разных сочетаниях одни и те же породы объединяли, а их доли крови суммировали. 2. Для получения суммарной доли крови пород степени их кров-ности суммировали в соответствии с направлением их продуктивности. 3. Обусловленное числом скрещиваний количество дроблений степени кровности указывает также на число расщеплений хромосом, представляющее интерес с позиций обмена факторами между хроматидами гомологичных хромосом, а, следовательно, повышения частоты свободной комбинации и взаимодействия генов при образовании гамет.
Определение числа пород и кратности дробления степеней кровности можно показать на примере породной комбинации, помещённой в группе 3 – КБ х ПБ и представляющей собой следующее сочетание пород:
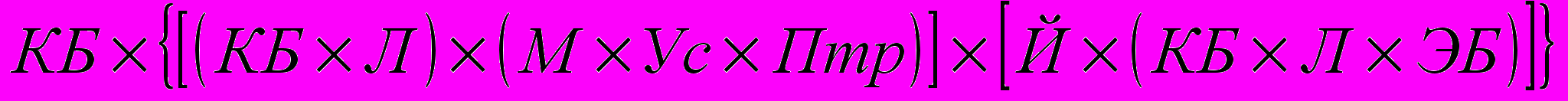
Таким образом, в указанном сочетании было использовано 7 пород, принявших участие в 9 скрещиваниях, т.е. подвергших-ся 9 делениям степеней кровности.
Еще сложнее генетическая модель группы 5 – (КБ х ПБ) х ПБ, которую можно записать так:
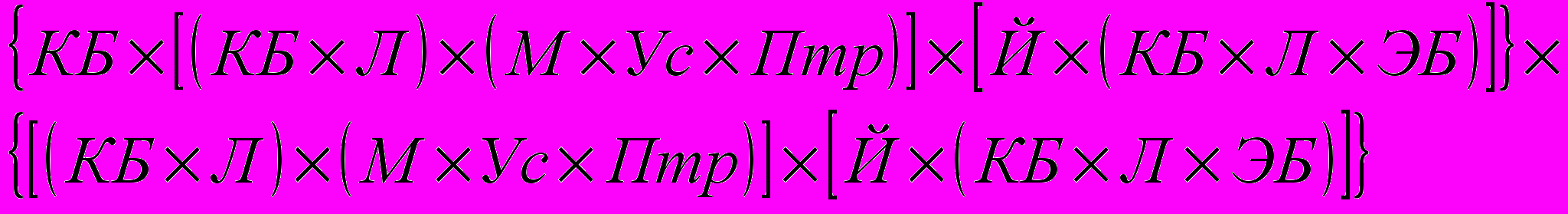
По степеням кровности участвовавших в скрещивании пород это составит:
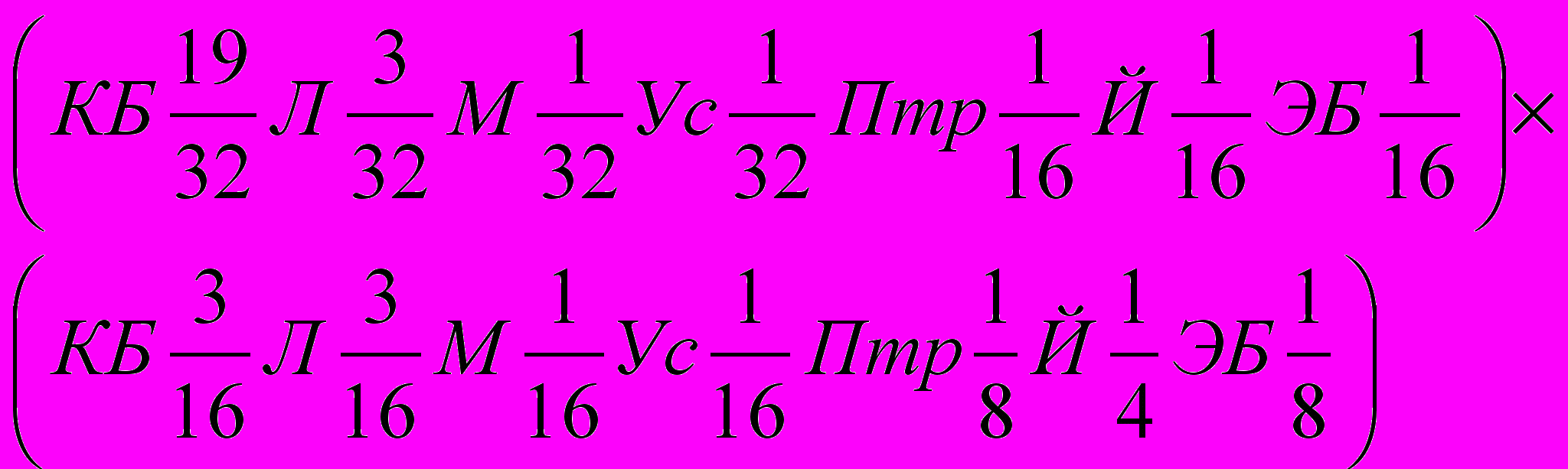 ,
,или
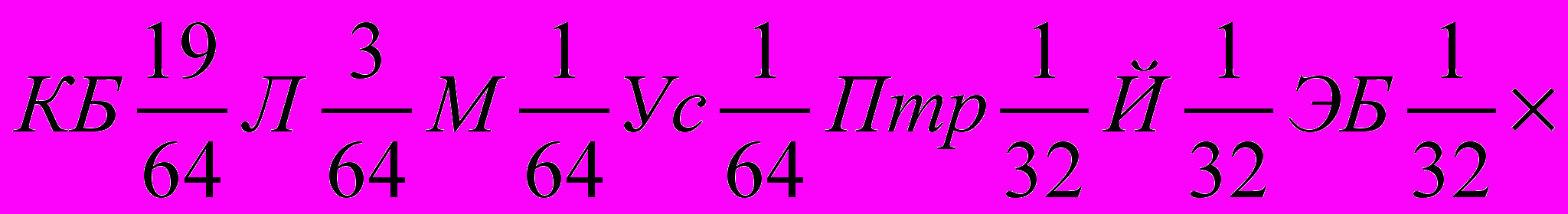
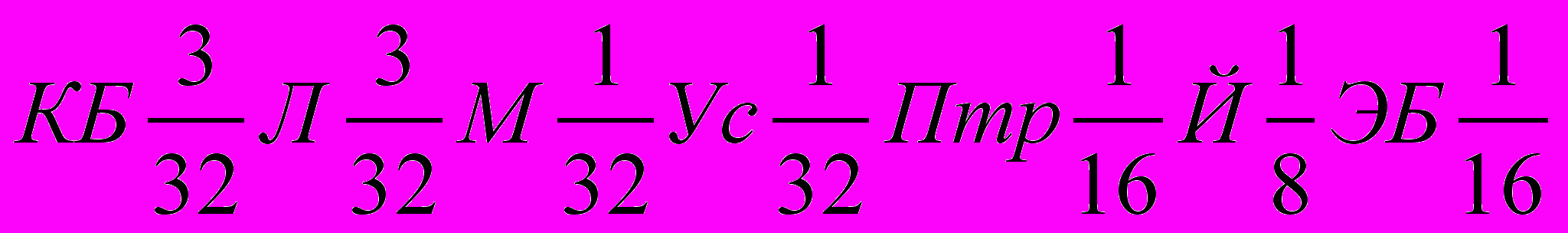 ,
,что при суммировании долей крови повторяющихся пород будет:
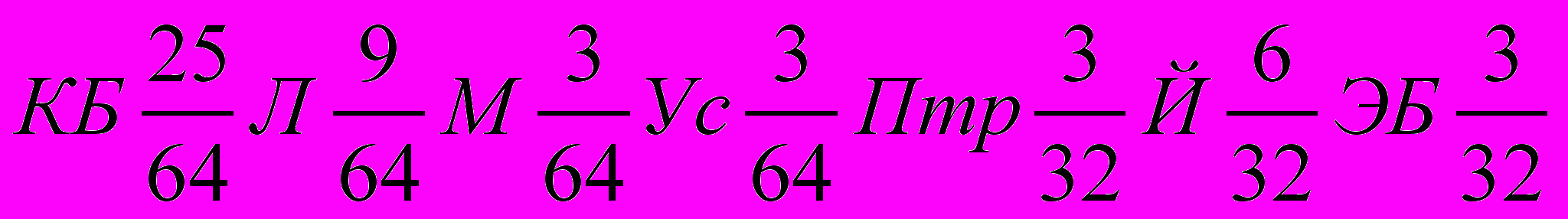 , а
, а в долях единицы –
КБ0,390625Л0,140625М0,046875Ус0,46875Птр0,09375Й0,1875
ЭБ0,09375
В данном сочетании насчитывается 7 пород, скрещивание которых дало 18 дроблений долей крови (долей генов).
Генетический вклад пород в генофонд породных сочета-ний (табл. 1) по долям крови и степень их изменчивости показа-ны в таблице 2.
- Кровность пород и степень ее изменчивости, %
| Группа | Условное обозначение пород | Коэф- фици-ент измен-чивос-ти | |||||||
| КБ | Л | М | Ус | Птр | Уэ | Й | ЭБ | ||
| 1 | 100,0 | - | - | - | - | - | - | - | - |
| 2 | 50,0 | 50,0 | - | - | - | - | - | - | 25,0 |
| 3 | 59,4 | 9,4 | 3,1 | 3,1 | 6,2 | - | 12,5 | 6,2 | 12,0 |
| 4 | 75,0 | 25,0 | - | - | - | - | - | - | 25,0 |
| 5 | 39,1 | 14,1 | 4,7 | 4,7 | 9,4 | - | 18,7 | 9,4 | 9,6 |
| 6 | 79,7 | 4,7 | 1,6 | 1,6 | 3,1 | - | 6,2 | 3,1 | 14,2 |
| 7 | 20,3 | 20,3 | 6,2 | 6,2 | 12,5 | 6,2 | 18,7 | 9,4 | 6,9 |
Наиболее продуктивными оказались многопородные (7 – 8 – породные) комбинации в группах 3, 5, 6, 7 с числом дроблений степеней кровности, находившимся в пределах 9-20 (табл.1). Самая высокая мясная продуктивность отмечалась в наиболее выровненных по долям крови комбинациях 5 и 7 групп, у которых степень кровности породы с самым высоким долевым участием (в данном случае крупной белой породы) была менее 40%, а коэффициент изменчивости доли крови по всем породам был самым низким и составлял 9,6 и 6,9% соответственно (табл.2). У них отмечалось 18-20 дроблений степеней кровности, а суммарная доля крови пород мясо-сального направления продуктивности была на уровне 26-43, а мясного направления – 56-73%.
В многопородных комбинациях доля крови крупной белой породы в наибольшей степени повышалась в том случае, когда она использовалась в качестве отцовской формы на заключи-тельном этапе скрещивания (группа 6).
У животных этой группы мясная продуктивность состави-ла 54,2% и находилась примерно на уровне двухпородных ком-бинаций свиней крупной белой и ландрас пород (группы 2 и 4) с самым низким числом дроблений (1-2) долей крови и самым высоким коэффициентом их изменчивости, равным 25%.
Рассматриваемую схему воспроизводительного скрещива-ния, предусматривающую спаривание животных не отдельных пород, а сложных породных комбинаций, можно, вероятно, назвать способом «модульного создания высокопродуктивных генотипов». Преимущества данной схемы вполне очевидны. Они заключаются, во-первых, в использовании при создании проектного генотипа генов многих пород в нужном соотноше-нии, во-вторых, в сокращении сроков выведения породы за счёт одновременного (параллельного) создания и скрещивания авто-номных полигибридных модулей и ускорения, таким образом, формирования единой генетической основы новой популяции, в-третьих, в получении однородных по долям крови (генов) высокопродуктивных групп животных и целых популяций.
Метод гомогенизации наследственности или модульного создания высокопродуктивных генотипов свиней следует рас-сматривать как важное средство типизации животных в поли-гибридных скрещиваниях при выведении пород (а также произ-водстве товарных гибридов) и эффективен лишь в сочетании с целенаправленным отбором и подбором в процессе селекции. Генетическая консолидация скороспелой мясной породы наряду с гомогенизацией наследственности достигалась также путём:
- повышения концентрации однотипных генов с помощью целенаправленного отбора и гомогенного подбора в про-цессе преимущественной (тандемной) селекции зональ-ных и заводских типов по одному - двум наиболее важ-ным хозяйственно полезным признакам;
- создания однотипных животных по направлениям про-дуктивности и степени развития признаков в ходе после-дующей селекции по независимым уровням на целевой стандарт.
Наследственность породы формируется в процессе селек-ции по фенотипическому сходству. В отличие от инбридинга, приводящего к усилению концентрации идентичных генов неза-висимо от их эффекта, при подборе по принципу «подобное с подобным» взаимодействуют гены, вызывающие сходный эф-фект, независимо от того, являются они аллелями, или нет. Такая система опосредованного отбора и подбора не остается индифферентной к механизму гомозиготизации и при адди-тивных эффектах приводит в процессе длительной селекции к постепенному изменению признаков.
Смешение крови животных разных пород в многопо-родных скрещиваниях становится причиной уменьшения степе-ни кровности каждой из них в генофонде потомства, снижающей вероятность доминирования наследственности любой из пород, с одной стороны, и расширяющей разнообразие генных комби-наций, - с другой. С увеличением числа скрещиваемых пород в одно спаривание, как это происходит в сочетаниях многопо-родных гибридов, повышается кратность расщепления закоди-рованной в хромосомах наследственности животных разных пород, разложения ее на мелкие составные части и возможность формирования на этой основе новой наследственности.
В ходе последующей селекции создаются предпосылки для рекомбинации сходных по своему действию генов, повышения на новой основе суммарной степени кровности однотипных животных и увеличения, таким образом, генного вклада живот-ных желательных генотипов в генофонд потомков, а, следова-тельно, и улучшения селекционных признаков.
Усиление признаков бывает значительнее при гомогенном подборе по степени выраженности этих признаков, что обуслов-лено сходством химизма наследственности, протекающих в организме однотипных родителей биохимических процессов, обеспечивающих специфику и результаты аллеломорфных взаи-модействий, взаимного обогащения и постепенного формиро-вания на этой основе новой наследственности. Установлена прямая зависимость наследования признаков от степени кров-ности сходных по направлению и уровню продуктивности участвующих в скрещивании пород: чем выше суммарная доля генов участвующих в скрещивании сходных пород, тем выше аддитивный эффект наследования признаков, присущих данным породам.
3. Моделирование процесса роста и развития
В связи с изучением процесса роста и развития как важного селекционного признака и разработкой на этой основе теории высокой скорости роста (15,28) появляется возможность регулирования этого процесса в целях повышения продук-тивности животных, базирующегося на следующих принципах:
- связи великорослости как конечного результата роста и скороспелости;
- зависимости живой массы свиней в процессе роста от интенсивности роста на отдельных стадиях онтогенеза.
Первый принцип основывается на единстве противоречий двух сторон единого процесса роста и развития, которое можно сформулировать так: дифференциация клеток и тканей организ-ма является следствием ускорения и причиной замедления их роста. Применительно к признакам скороспелости и велико-рослости эту связь можно выразить следующим образом: чем выше интенсивность роста свиней на ранних стадиях онтоге-неза и короче фаза активного роста, тем выше скороспелость и меньше живая масса взрослых животных (великорослость) как конечный результат роста.
Второй принцип опирается на важные закономерности индивидуального роста, относящиеся к увеличению живой мас-сы животных в связи с изменением их прироста на отдельных стадиях онтогенеза и выражающихся в следующем:
- количественные изменения процесса роста на одной стадии онтогенеза вызывают адекватные изменения на другой;
- каждому уровню роста на одной стадии соот-ветствует свой оптимальный уровень на другой.
Указанными закономерностями обусловлены разные типы роста у свиней разных пород, встречающиеся также, правда, в менее выраженной форме, в пределах внутрипородной измен-чивости, что свидетельствует, во-первых, о генетической обус-ловленности типов роста и возможности использования их в селекции и, во-вторых, о возможности моделирования их как сложных селекционных признаков, включающих в себя ско-рость, интенсивность, продолжительность, великорослость и другие характеристики процесса роста (15).
В работе по созданию скороспелой мясной породы форми-рование желательного типа роста осуществлялось по модели, разработанной путем комбинирования источников ускорения роста у свиней крупной белой породы и ландрас (33). Первая из них характеризуется быстрым ростом и умеренной скороспе-лостью. Животные этой породы отличаются высокой скоростью роста до 6 – 7 – месячного возраста, большой продолжительнос-тью наиболее активного роста (в период от 6 до 12 – месячного возраста) и большой великорослостью. Вторая отличается быс-трым ростом, высокой скороспелостью и умеренной высокорос-лостью. Животные характеризуются высокой скоростью роста, скороспелостью великорослости и достаточно большой продол-жительностью активного роста на ранней стадии онтогенеза.
Модель желательного типа проектировалась по принципу совмещения типов роста указанных пород и сочетала в себе великорослость одной породы с высокой скороспелостью другой. Разработанная формула для определения желательного типа роста показывает, как будет изменяться в онтогенезе живая масса одной породы, если она будет нарастать по схеме вели-корослости другой:
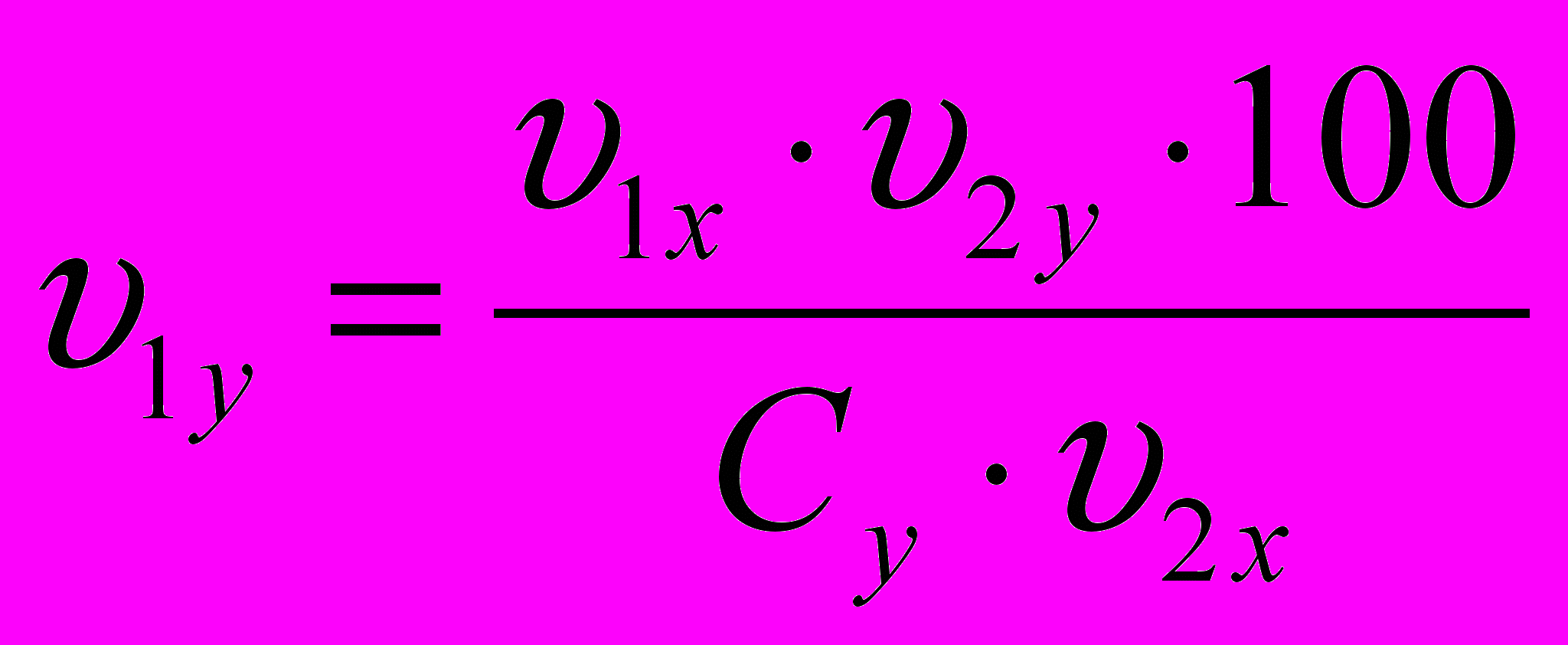 ,
,где:
 и
и 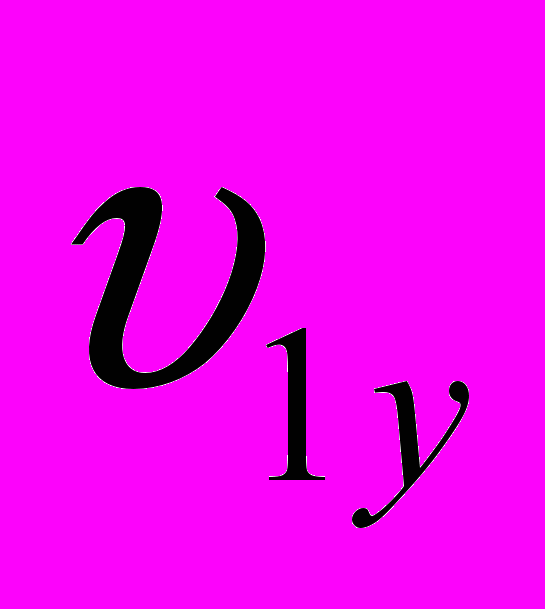 - живая масса животных x и y пород в онтогенезе;
- живая масса животных x и y пород в онтогенезе;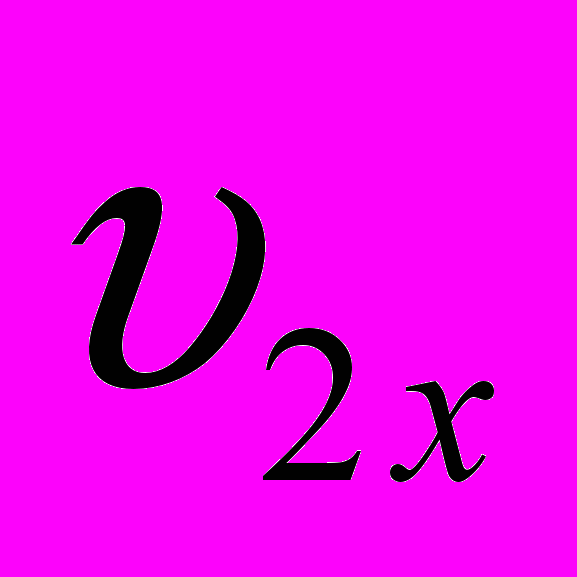 и
и 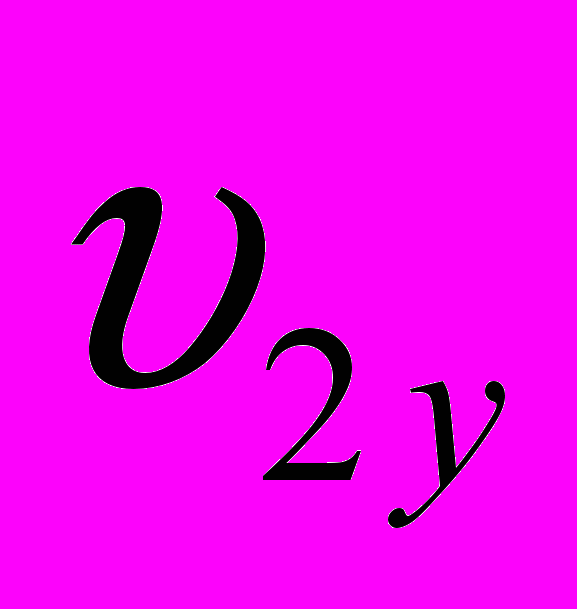 - конечная живая масса (великорослость) животных x и y пород;
- конечная живая масса (великорослость) животных x и y пород;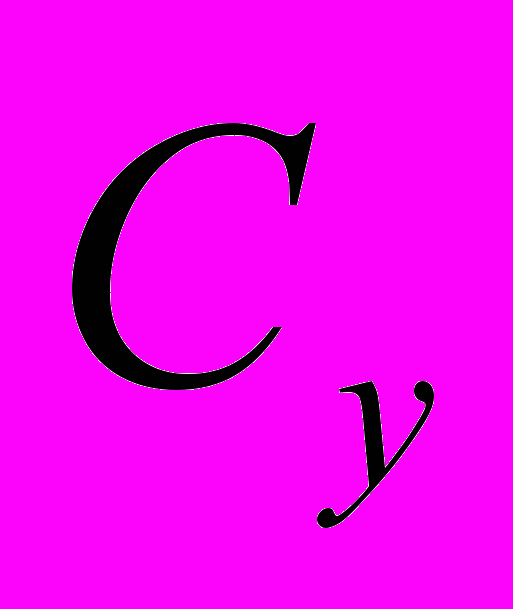 - показатель скороспелости великорослости (доля живой массы в онтогенезе от живой массы в момент завершения роста) породы у, принятой за 100%.
- показатель скороспелости великорослости (доля живой массы в онтогенезе от живой массы в момент завершения роста) породы у, принятой за 100%. Предположим, надо узнать, как будет изменяться в онто-генезе живая масса животных породы у, если она будет нарас-тать по схеме увеличения скороспелости великорослости поро-ды х.
Пример. Определить живую массу хрячков породы у в возрасте 6 месяцев, если их скороспелость будет равна скоро-спелости великорослости хрячков породы х, живая масса кото-рых в возрасте 6 месяцев составляет 88 кг и конечная живая масса (великорослость) 313 , а конечная живая масса породы у будет 336 кг.
Тогда

Если повышение скорости роста животных с проек-тируемым ростом предусматривается не только путём повы-шения интенсивности роста (т.е. увеличения скороспелости великорослости), но одновременного увеличения великорослос-ти (конечной живой массы), тогда конечную живую массу поро-ды у (
) в уравнении следует заменить на конечную живую массу животных проектируемого типа (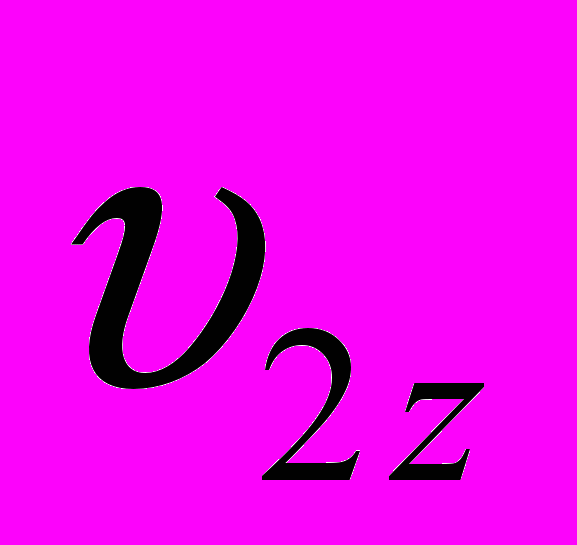 ).
).Тогда формула примет такой вид:
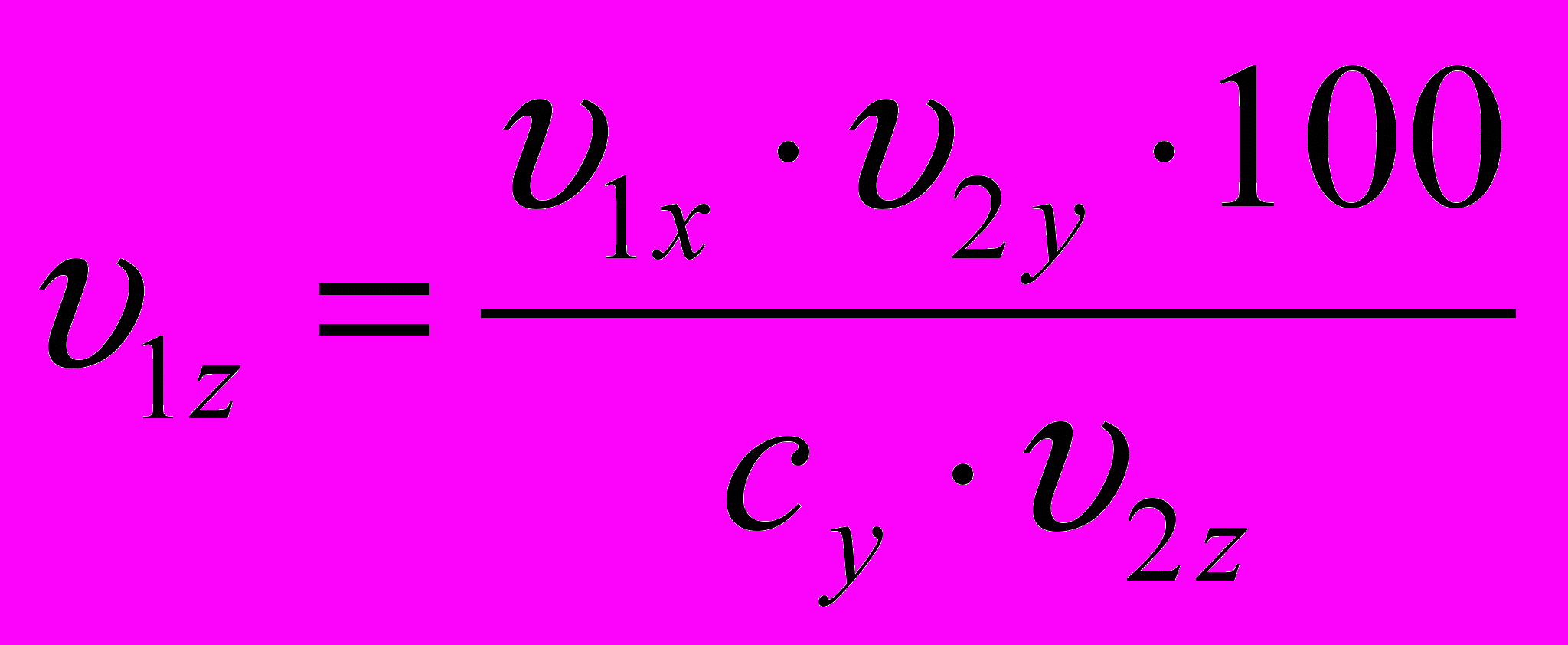 ,
,Где
- конечная живая масса (великорослость) проек-тируемой породы.Таким образом, модель типа роста животных скороспелой мясной породы вобрала в себя особенности роста свиней великорослой крупной белой породы и скороспелой породы ландрас. Её принципиальное отличие от исходных типов роста заключается в смещении интенсивности роста на более ранний период онтогенеза в сравнении с крупной белой породой и увеличение продолжительности наиболее активного роста на более поздние сроки, чем у свиней породы ландрас. Такое изменение характера протекания процесса роста способствует значительному повышению абсолютной скорости роста у живот-ных проектируемого типа и даёт возможность получать при ин-тенсивном откорме желательную реализационную массу свиней 120 кг примерно в возрасте 6 месяцев, а 100 кг – в 5,5 месяцев при среднесуточном приросте 800г и более.
Желательный тип роста по разработанной модели форми-ровали двумя путями.
Первый путь - это набор в полигибридных скрещиваниях пород, сходных по типам роста, позволивший сформировать и закрепить в генетическом отношении смоделированный тип роста. Из 13 исходных пород, принимавших участие в создании скороспелой мясной породы, только 2 породы было мясо-сального (универсального) направления продуктивности, а остальные – мясного направления. Соотношение долей крови в породе было примерно: 23% крупной белой породы и 77% ландрас и других, сходных с ней пород.
Второй путь – включение смоделированного типа роста в целевой стандарт породы, селекционировавшейся на этот стан-дарт по фенотипическому сходству. Стандартом предусмат-ривалось, во-первых, повышение скорости роста примерно до 800г в суки, во-вторых, проводить селекцию не до 100кг, как это принято в отечественной практике, а до 120 кг, увеличивая продолжительность откорма в зоне наиболее активного роста на 20-25 суток, что служит одним из главных источников уско-рения роста. Смещение продолжительности интенсивного от-корма в зоне наиболее активного роста на более поздний период под генетическим контролем одновременно мясной продук-тивности, позволяет повышать среднесуточный прирост до 800-900г в сутки.
4. Фенотипическая оценка молодняка по собственной продуктивности
Действенным фактором повышения скорости роста, конверсии корма и мясной продуктивности свиней при крупномасштабной селекции служит массовая оценка животных по собственной продуктивности на основе скорости роста и толщины шпика без проведения трудоёмкой производственной операции по учёту расхода корма на прирост живой массы. Способность животных конвертировать корма в продукцию, пожалуй, единственный в своём роде признак, который можно не измерять, но о котором с большой определенностью можно судить по скорости роста свиней.
Для этого достаточно знать важнейший принцип: только те животные расходуют мало корма на продукцию, которые быстро растут.
Метод основан на определении высокой корреляции скорости роста и конверсии корма, приближающейся к, несуществующей в биологии, функциональной зависимости (14), получившей научное доказательство по соотношению под-держивающего и продуктивного корма в структуре его общих затрат на продукцию. Подтверждение приоритета выражено гра-фическим построением указанных затрат в форме асимпто-тических кривых, устремлённых справа вверх налево (15).
Оценочная шкала животных по величине прироста и толщине шпика строится по принципу учёта соотношения классов оценки по обоим указанным признакам. Соблюдение этого принципа предполагает повышение класса оценки животных в связи с повышением скорости роста и уменьшением толщины шпика, а также исключает возможность завышения класса оценки по толщине шпика при снижении скорости роста животных. Построенная по такому принципу шкала имеет форму корреляционной решётки, отражающей отрицательную связь (15).
Сопоставимое соотношение селекционной ценности клас-сов позволяет повысить надёжность оценки животных по уров-ню развития обоих селекционных признаков. За величину клас-сового промежутка по среднесуточному приросту принято 100г, а по средней толщине шпика на спине – 5мм. Изменение каж-дого из этих признаков на один класс соответствует по своей селекционной ценности изменению относительного содержания мяса и сала в теле свиней на 2-4%.
Экспериментальная проверка эффективности применения данного метода в четырёх поколениях дала следующие резуль-таты: возраст животных по достижении живой массы 100кг у свиней разных линий сократился на 20-30 суток (8,8-10%), среднесуточный прирост за период выращивания с момента отъёма повысился на 72-84г (14,7-17,1%), толщина шпика над 6-7-м грудными позвонками уменьшилась на 2-5 мм (9,1-11,8%).*
В процессе создания породы возраст по достижении живой массы 120кг в F4 в сравнении с исходными формами сократился с 209 до 188 суток, среднесуточный прирост увеличился с 767 до 847г, расход корма на 1 кг прироста живой массы снизился с 3,79 до 3,38 корм. ед., а доля мяса в туше повысилась с 56,9 до 58,0% (30).
Значение этого метода оценки для крупномасштабной селекции состоит в том, что он, благодаря своей простоте и доступности даёт возможность проводить проверку всего селекционируемого поголовья и повышать результативность отбора. Но великая роль его заключается в предоставлении возможности оценивать и отбирать животных не в искусственно созданной среде, а в условиях естественной конкуренции животных с учётом влияния генетических, паратипических факторов, взаимодействия генотипа и среды, выявлять особей, способных лучше поедать корма и конвертировать их в продукцию. Поэтому оценка молодняка по собственной продуктивности, а свиноматок и хряков – по результатам выращивания их потомства, явилась ключом к повышению эффективности селекции и способствовала сокращению сроков выведения скороспелой мясной породы примерно в 1,5 раза в сравнении с традиционными методами (27).
* Кабанов В.Д., Бадаева В.А., Ж. «Животноводство», №10, 1977г.
5. Формирование разветвленной генеалогической структуры породы
В условиях крупномасштабной селекции, проводимой на обширных территориях разных зон нашей страны с разнообраз-ными природно-климатическими, хозяйственными условиями, охватом большого числа хозяйств и поголовья, вынуждены при-бегать к формированию сложной генеалогической структуры по-роды, включающей в себя не только линии хряков и семейства свиноматок, но также зональные и заводские типы. Важной ха-рактерной особенностью скороспелой мясной породы служит наличие в ней, помимо линий и семейств, укрупненных зональ-ных и заводских типов, что при генетическом их сходстве при-дает породе большее генофенотипическое разнообразие, повы-шенную жизненность и продуктивность.
Генетическая формула породы с учетом степени кровности составляющих ее заводских типов и исходных пород выглядят так (31):
| Липецкий | 1) 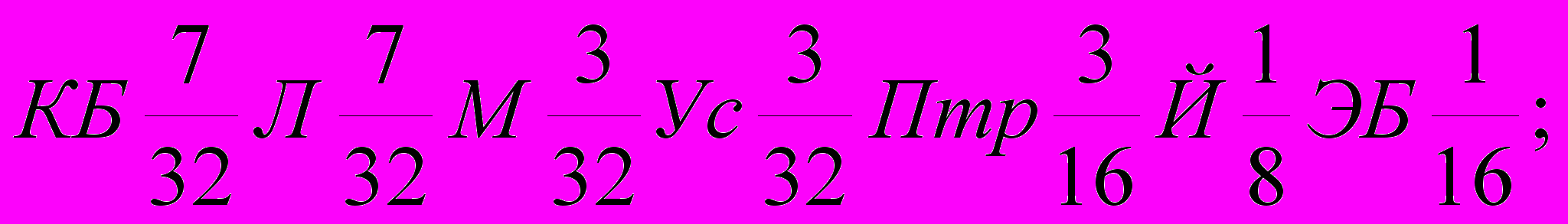 |
| 2) 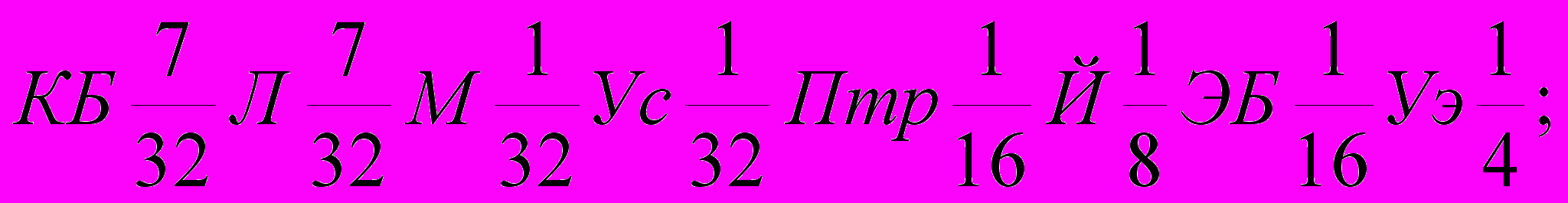 | |
| Степной | 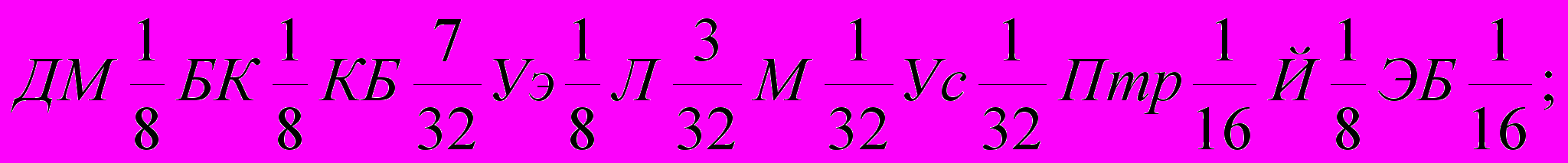 |
| Красно-дарский | 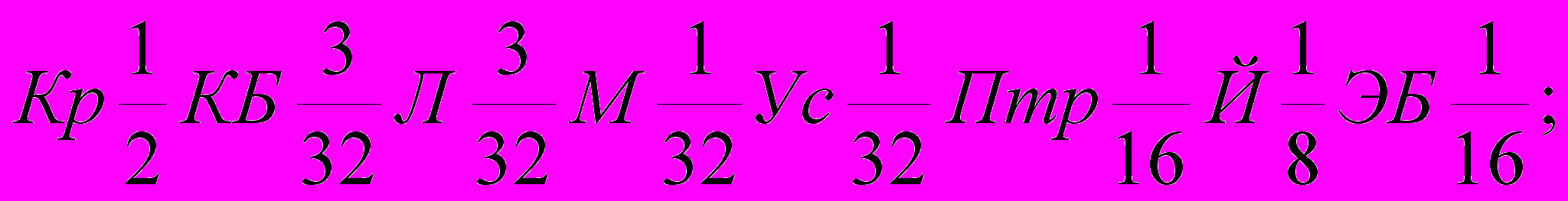 |
| Ленин-градский | 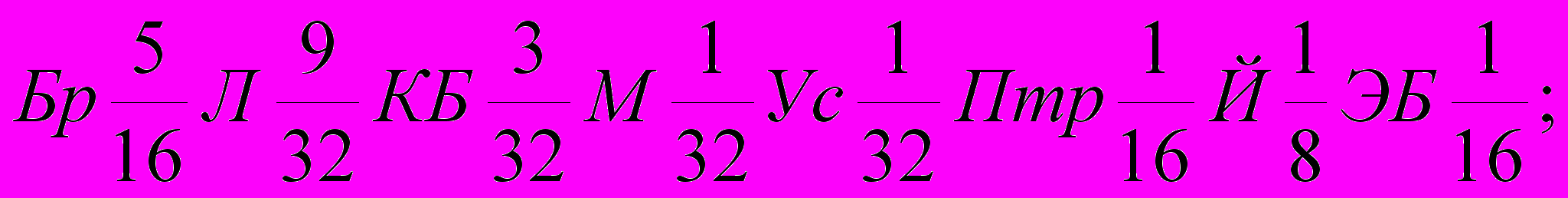 |
| Сибир-ский | 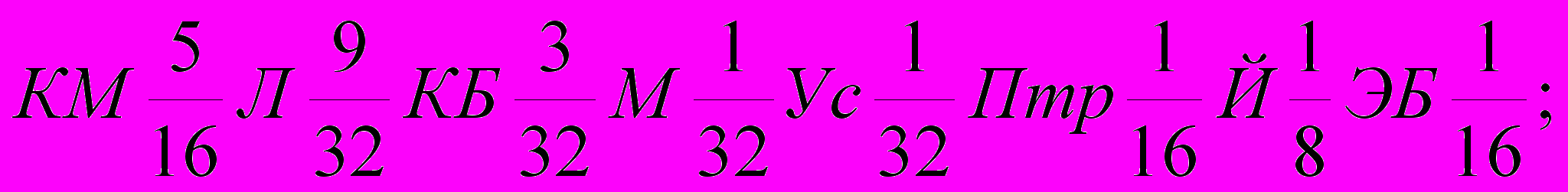 |