
Цитофизиологические особенности крови промысловых рыб Волго-Каспийского канала
| Вид материала | Автореферат |
- Рыбохозяйственные и экологические аспекты эффективности искусственного воспроизводства, 422.61kb.
- Комментарий начальника Регионального управления фскн россии А. Долбни о целях проведения, 12.87kb.
- Характеристики отрядов, подотрядов и семейств рыб, их многообразие в зависимости, 587.95kb.
- Итоги и перспективы изучения паразитов и болезней морских промысловых гидробионтов, 211.89kb.
- Урок по теме: «Костные рыбы», 73.15kb.
- Лекция №15. Тема: Товароведная характеристика и таможенная экспертиза промысловых рыб, 130.05kb.
- Урок по теме: многообразие рыб. Класс костные рыбы. Цель, 59.64kb.
- Рабочая учебная программа по дисциплине Гематология Для специальности, 174.73kb.
- План лекций для студентов педиатрического факультета на весенний семестр 2011-2012, 50.89kb.
- Основные группы рыб, их роль в природе и практическое значение, 29.02kb.
3.1. Типы патологических изменений форменных элементов периферической крови промысловых рыб Волго-Каспийского канала
При анализе мазков крови среди нормальных клеточных элементов, были установлены разнообразные типы отклонений их структуры. Присутствие их в составе периферической крови с определённой статистической частотой рассматриваются как свидетельство проявления хронического токсикоза организма рыб В-К канала. Характерными патологическими изменениями структуры эритроцитов крови, обнаруженными при анализе мазков крови промысловых рыб, были следующие: пойкилоцитоз (появление в крови эритроцитов различной формы); анизоцитоз (разноразмерность); нарушение осморезистентности мембраны клетки; псевдоагглютинация; цитолиз (разрушение эритроцитов); ядра-тени; вакуолизация.
Среди нарушений структуры ядер эритроцитов установлены следующие: хроматинолиз (уменьшение видимого количества хроматина, ядро окрашено в светлые тона, контуры его сохраняются); протуберанцы кариоплазмы во внутриклеточное пространство; гиперхроматоз стенки ядра эритроцитов (ядра имеют форму кольца, контур четко окрашен, центр ядра светлый, за счет концентрации хроматина по периметру ядра); кариолизис; кариорексис; инвагинация ядра; деформация ядра; пристеночные ядра; эксцентричные ядра (ядро смещено от центра); шистоцитоз; амитоз. Отмеченные нарушения, относимые ранее к артефактам, в условиях В-К канала в настоящее время представляют собой реальные этапы гематологических процессов. Так, появление фестончатых краев эритроцитов, ядерных теней, безъядерных эритроцитов, по-видимому, есть последовательные звенья одной цепи. Аналогичное мнение высказано ранее Житеневой Л.Д., 1999.
3.2. Оценка физиологического состояния особей судака обыкновенного (Stizostedion lucioperca) по состоянию показателей крови и инвазированности паразитами
Исследование инвазированности судака Волго-Каспийского бассейна показало, что паразитические сообщества были представлены 16 видами гельминтов, относящихся к 7 классам. Частота встречаемости особей, зараженных гельминтами за период исследования, в среднем составляла 98,0 % весной и 96,0 % осенью, т.е. практически вся рыба была заражена паразитами. Отмечено незначительное снижение уровня заражения рыб паразитами к осени. Паразитирование гельминтов в организме судака может отражаться на изменении параметров периферической крови. Обнаружена корреляционная зависимость между паразитированием в организме судака гельминтов и изменением параметров периферической крови рыб. Паразитирование отдельных гельминтов вызывало снижение количества эритроцитов в объеме крови (R=-0,57 при р<0,05), увеличение содержания гемоглобина в крови (R=+0,60, р<0,05), повышение содержания Нф (R=+0,53 при р<0,05) и Пэф (R=+0,67 при р<0,05), увеличение абсолютного количества лейкоцитов литре крови (R=+0,51, р<0,05), что является ответной реакцией организма, направленной на нейтрализацию неблагоприятного эндогенного воздействия.
Показатели периферической крови судака приведены в таблице 3.2.1. Осенью 2006 г. отмечено недостаточное содержание гемоглобина и общего количества эритроцитов в литре крови судака. При этом наблюдались относительно низкие показатели СОЭ. В 2007 году было отмечено повышенное содержание гемоглобина у большинства исследованных рыб в летний (48%) и осенний (48%) периоды исследования. Количество эритроцитов было на достаточном уровне у 56% исследованных рыб летом и 52% рыб осенью. Концентрация гемоглобина в крови особей судака в 2008 году была достоверно ниже его уровня в 2007 году (при р<0,05). Данные, приведённые в табл. 3.2.1, свидетельствуют о некотором снижении содержания гемоглобина и количества эритроцитов в литре крови рыб летом и осенью 2008 года в сравнении с аналогичными периодами 2007 года.
Таблица 3.2.1
Показатели крови, отражающие физиологическое состояние
особей судака В-К канала
| Показатели крови: | Условная норма | 2006 | 2007 | 2008 | Среднее | Общее среднее M±m | ||||||
| Осень M±m | Лето M±m | Осень M±m | Лето M±m | Осень M±m | Лето M±m | Осень M±m | ||||||
| Hb, г/л | 70,0-72,0 | 61,9±2,2 | 76,2±2,8 | 75,0±2,2 | 63,8±1,9 | 66,9±1,2 | 70,0±2,3 | 67,9±2,2 | 69,0±2,3* | |||
| СОЭ, мм | | 1,8±0,2 | 1,6±0,2 | 1,5±0,1 | 1,0±0,1 | 1,5±0,1 | 1,3 ± 0,1 | 1,6±0,2 | 1,4±0,2 | |||
| СГЭ, пг | 22,4-42,2 | 46,3±2,6 | 38,0±1,5 | 39,5±1,6 | 39,0±1,2 | 38,1±1,2 | 38,5±1,3 | 41,3±1,8 | 39,9±1,6* | |||
| Колич. эритроцитов *1012 в л | Лето: 1,67-1,72 Осень 1,70-1,85 | 1,30±0,10 | 1,88±0,08 | 1,84±0,08 | 1,54±0,06 | 1,64±0,05 | 1,71±0,07 | 1,59±0,08 | 1,65±0,07* | |||
| Колич. лейкоцитов *109 в л | 35,0-95,0 | - | 89,5±12,3 | 90,0±10,9 | 149,2±16,9 | 69,2±7,4 | 119,3±14,6 | 79,6±9,1 | 99,5±11,9 | |||
| ПЭФ, % | 3,0 | 0,4±0,2 | 0,1±0,09 | 0,04 ± 0,03 | 0,0 | 0,2±0,08 | 0,07±0,04 | 0,2±0,05 | 0,13±0,05 | |||
| ЭФ, % | | 0,5±0,20 | 0,09±0,07 | 0,0 | 0,36±0,15 | 0,08±0,06 | 0,23±0,11 | 0,19±0,09 | 0,21±0,09 | |||
| НФ, % | 0,1-2,0 | 4,0±1,30 | 9,8±2,50 | 11,9±4,4 | 2,70±1,07 | 1,74±0,36 | 6,25±1,79 | 5,88±2,02 | 6,06±1,90 | |||
| ЛФ, % | 86,4 | 93,0±1,30 | 86,0±3,10 | 84,7±5,0 | 96,3±1,1* | 97,5±0,4 | 91,1±2,1 | 91,7±2,2 | 91,4±2,2* | |||
| Молодые лейкоциты % | - | 0,0 | 3,1±0,7 | 0,0 | 0,02±0,02 | 0,10±0,04 | 1,56±0,36 | 0,03±0,01 | 0,83±0,19 | |||
| Мн, % | 2,1 | 0,1±0,06 | 0,5±0,48 | 3,4±0,7 | 0,68±0,20 | 0,56±0,13 | 0,6±0,34 | 1,35±0,3 | 0,98±0,32 | |||
Повышение количества Нb и Эр может зависеть от воздействия многих токсических факторов, к этому может приводить также нехватка кислорода в воде. Насыщение эритроцита гемоглобином (СГЭ, пг) во все периоды исследования соответствовало норме.
«Белая» кровь исследуемых особей судака носила ярко выраженный лимфоидный характер. Гранулярные формы лейкоцитов были представлены псевдоэозинофилами (Пэф), эозинофилами (Эф) и нейтрофилами (Нф). Агранулярные формы: моноцитами (Мн), лимфоцитами (Лф). Особенности в соотношении различных форм лейкоцитов в периферической крови судака осенью 2006 г. были представлены относительно невысоким содержанием фагоцитов (моноциты, палочкоядерные, сегментоядерные нейтрофилы), что может объясняться физиологическими особенностями рыб. Абсолютное количество лейкоцитов в крови рыб на протяжении 2007 г. было в пределах нормы. Анализ иммунограммы показал, что весной 2007 г. у рыб наблюдалась выраженная моноцитопения. Низкий процент моноцитов в лейкограмме может быть связан с тем, что основная масса моноцитов сконцентрировалась в очагах воспаления, трансформируясь в макрофаги, что часто наблюдается при длительном воздействии неблагоприятных факторов среды (Житенева Л. Д. и другие, 2004). Количество нейтрофилов было повышенным, но они были на стадии молодой клетки имеющей слабую функциональную активность. Осенью 2007 г. наблюдался моноцитоз и нейтрофилия, проявляющиеся на фоне снижения содержания остальных клеток гранулярного ряда. Это может свидетельствовать о неблагоприятном состоянии организма рыб и иммунологической недостаточности. Изменения в соотношении различных форм лейкоцитов в 2008 г. были представлены значительным повышением количества лимфоцитов, на фоне снижения эозинофилов и моноцитов, хотя количество нейтрофилов колебалось в пределах нормы, также как и в 2007 г. отмечали их слабую фагоцитарную активность. Деструктивные изменения нейтрофилов (вакуолизация цитоплазмы) охватывали не более 0,1 % клеток на мазке крови у 3,0 % исследованных особей. Зрелые лимфоциты были представлены, в основном, клетками узкоплазменной формы. У отдельных рыб в 2008 г. (20,0 % весной, 48,0 % осенью) отмечали явление кариолиза в лимфоцитах в яркой форме (рис. 3.2.1).
-
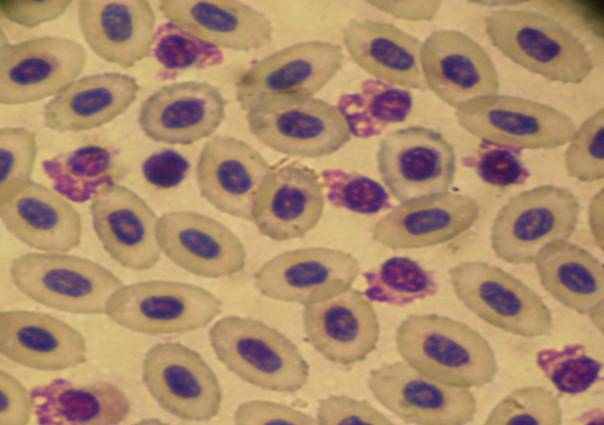
Рисунок 3.2.1. Кариолиз лимфоцитов судака обыкновенного В-К канала (осень 2008г.).
ув. × 1000
Подобные деструктивные изменения ядерных структур могут быть спровоцированы наличием токсических агентов в организме, а также при заболеваниях (вирусные, паразитарные). Исследованные особи судака были неоднородны по характеру гематологических показателей, что может быть обусловлено индивидуальной резистентностью их организма.
Гематологические показатели находятся в тесной связи с возрастом и стадиями зрелости гонад рыб, отражая физиологическое состояние их организма. По мере созревания гонад наблюдается изменение ряда гематологических показателей, отмечено повышение гемоглобина, СГЭ, количество лейкоцитов в пересчете на литр крови, это было отмечено ранее и другими авторами (Житенева Л.Д., 1986; Головина Н.А., 1997). После нереста (VI-II СЗГ) отмечено снижение гемоглобина, СГЭ и СОЭ. У самок после нереста возросло количество Нф, что, вероятно, объясняется резорбцией остаточных ооцитов и пустых фолликулов. Повысилось количество моноцитов, что возможно связано с участием этих клеток в процессе овуляции, они выделяют ферменты, обеспечивающие освобождение созревших яйцеклеток фолликула (Житенева Л.Д., 1987; Иванов А.А., 2003), и, в среднем, у самок отмечено повышенное количество моноцитов (0,64%) по сравнению с самцами (0,18%). Установлено, что у самок, находящихся на II СЗГ и, в среднем, количество гемоглобина было выше. У самцов количество эритроцитов и лейкоцитов в пересчете на литр крови рыб ниже, чем у самок. Полученные результаты свидетельствуют, что организм рыб обладает большими компенсаторными возможностями. Физиологическое состояние организма рыб, его ослабление проявляется в зависимости от зрелости гонад. Значительные отклонения большинства показателей крови от условной нормы, можно рассматривать не только как естественные изменения. Значительное количество патологических изменений показателей крови связано с проявлением хронического токсикоза рыб.
Таблица 3.2.2
Типы цитопатологических нарушений в эритроцитах судака В-К канала
| Основные типы нарушений | Количество особей (%) с нарушениями структуры клеток от общего количества обследованных рыб | ||||||
| 2006 | 2007 | 2008 | Среднее | ||||
| Осень | Лето | Осень | Лето | Осень | Лето | Осень | |
| Пойкилоцитоз | 32,0* | 78,0* | 46,0* | 88,0** | 72,0* | 83 | 50 |
| Анизоцитоз | 8,0* | 74,0* | 25,0* | 92,0** | 84,0** | 83 | 39 |
| Агглютинация | 12,0* | 17,0* | 8,0 | 68,0* | 8,0* | 42 | 23 |
| Изменение внешней плазматической мембраны | 76,0** | 96,0** | 83,0* | 72,0* | 52,0* | 84 | 70 |
| Смещение ядер к периферии | 0,0 | 100,0 | 100,0 | 60,0* | 28,0* | 80 | 64 |
| Пикноз | 0,0 | 43,0* | 0,0 | 68,0* | 52,0* | 55 | 26 |
| Деформация ядер | 32,0* | 56,0* | 71,0* | 0,0 | 0,0 | 28 | 34 |
| Кариорексис | 20,0* | 56,0* | 21,0* | 76,0* | 56,0* | 66 | 32 |
| Инвагинация ядра | 0,0 | 56,0* | 96,0** | 76,0* | 48,0* | 66 | 72 |
| Разрушение клетки | 0,0 | 70,0* | 92,0** | 76,0* | 36,0* | 73 | 64 |
| Ядра-тени | 0,0 | 65,0* | 42,0* | 100,0 | 52,0* | 82 | 47 |
| Хроматинолиз | 0,0 | 56,0** | 4,0 | 16,0* | 8,0* | 36 | 6 |
| Ядра-кольца | 0,0 | 0,0 | 0,0 | 56,0* | 28,0* | - | - |
| Протуберанцы | - | - | - | 16,0* | 16,0* | - | - |
| Вакуолизация цитоплазмы | 0,0 | 22,0* | 37,5* | 48,0* | 4,0 | 35 | 21 |
| Кариолиз | 0,0 | 17,0* | 4,0 | 44,0* | 12,0* | 30 | 8 |
| Шистоциты | 4,0 | 9,0* | 25,0* | 0,0 | 0,0 | 9 | 14 |
| Амитоз | 32,0* | 13,0* | 8,0 | 44,0* | 12,0* | 28 | 17 |
| Амитоз – деление только ядра | 0,0 | 13,0* | 21,0* | 8,0* | 12,0* | 10 | 16 |
| Количество исследованных рыб | 25 | 23 | 24 | 25 | 25 | 24 | 25 |
Примечание: * - р<0,05; ** -р<0,01 (р – достоверность доли, уровень значимости)
В периферической крови особей судака были обнаружены изменения в структуре эритроцитов, отличающиеся степенью проявления патоморфологических нарушений (табл. 3.2.2). К их числу относится: пойкилоцитоз эритроцитов, который отмечен во все сезоны исследования, в основном, их полиморфностью (максимально до 15,0 % клеток на мазке). Отмечены клетки грушевидной, серповидной, ромбовидной формы. Патологическая форма эритроцитов с отростками (характерна для угнетения эритропоэза) встречалась единично, у отдельных особей, независимо от сезона исследования. Снижение эластичности клеточной мембраны эритроцитов косвенно указывает на изменение её осмотических свойств. У одной особи (2008 г.) пойкилоцитоз охватывал до 72,0 % эритроцитов. У этой особи судака отмечено максимальное количество трематод Rossicotrema donicum (90 экз.), поражающих покровы тела. У всех исследованных рыб нарушения эритроцитарной мембраны проявлялись гемагглютинирующим эффектом без явных признаков деструкции. Этот показатель значительно снижался (до 8,0 %) осенью. Подобные явления, как правило, являются результатом действия неблагоприятных факторов как вне, так и внутри организма (интоксикация различной этиологии).
В летний период исследования отмечено увеличение доли рыб с эритроцитами, имеющими морфологические нарушения. Так, доля (%) рыб с эритроцитами меньшего диаметра в сравнении с нормоцитами, по отношению к общему количеству обследованных особей в 2007 и 2008 гг. возрастала в весенне-летний период относительно осени. Наличие микроцитов в периферическом русле не превышало 5,2 % от общего пула эритроцитов у отдельных особей и отражало относительно нормальный процесс гемопоэза судака. К числу патоморфологических изменений на мазках крови относятся эритроциты в стадии разрушения.
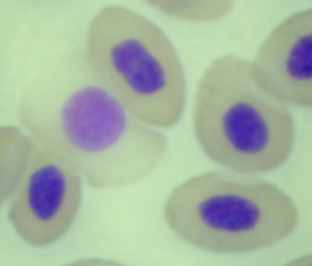 | 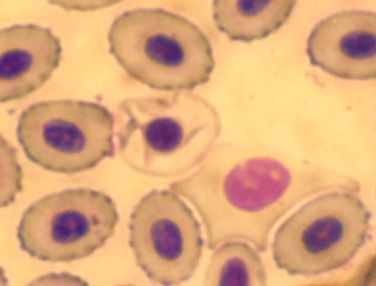 | | |
| Рисунок 3.2.2. Разрушение эритроцитов (судак, осень 2008), ув.×1000 | | ||
 | 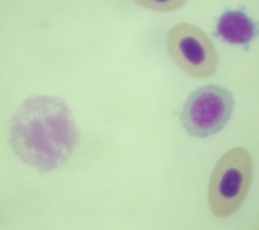 | ||
| Рисунок 3.2.3. Тени эритроцитов (судак, осень 2008), ув. х1000 | |||
У отдельных особей в 2007 г., отмечен высокий уровень цитолиза клеток крови (57% от общего числа исследованных клеток указанных экземпляров рыб) (рис. 3.2.2). В остальные периоды исследования встречаемость данного типа отклонений на мазках крови была не высокой, хотя и данное нарушение отмечали более чем у половины исследованных рыб. В 2007 и 2008 гг. в периферической крови рыб регистрировали разрушившиеся эритроциты (полный лизис). В летний период 2008 г. у всех исследованных особей рыб на мазках крови, регистрировали ядра-тени (рис. 3.2.3).
Деструктивные изменения эритроцитов были представлены деформацией, пикнозом ядра. Явление кариорексиса, хроматинолиза, гиперхроматоза стенки ядра (ядра-кольца), кариолиза с явлениями близкими фрагментозу (ареол хроматина вокруг ядра, «протуберанцы» кариоплазмы во внутриклеточное пространство) чаще отмечали в летний период (рис. 3.2.4.).
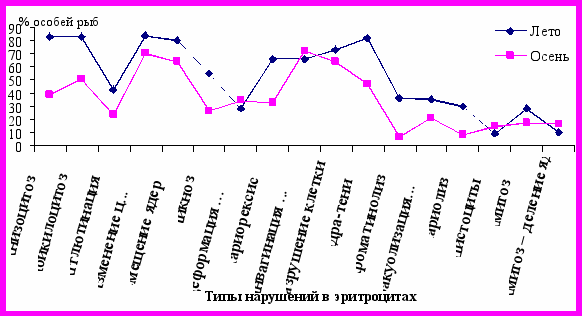
Рисунок 3.2.4. Доля особей судака (%) с нарушениями
в эритроцитах крови
Подобные патологические изменения ядра встречаются при различных повреждениях клетки. Изменение коллоидно-осмотического состояния ядра и цитоплазмы происходит вследствие торможения транспорта веществ через внешнюю плазматическую мембрану клетки (Шиффман Ф. Дж., 2001). На мазках крови были отмечены эритроциты в стадии амитоза. Уменьшение частоты встречаемости и интенсивности проявления цитопатологических изменений к осени (рис. 3.2.4.) может свидетельствовать: о некоторой стабилизации токсикологической и гидрохимической обстановки водной среды в осенний период, о более благополучном физиологическом состоянии рыб в результате летнего нагульного периода. Этому способствовало и небольшое снижение количества полостных паразитов рыб осенью.
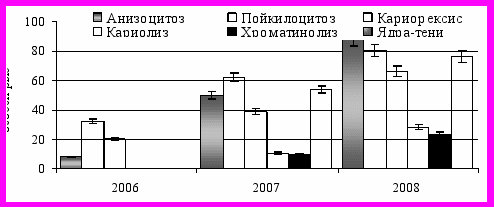
Рисунок 3.2.5.
Доля особей судака с цитопатологическими отклонениями (средний %)
в эритроцитах крови
В период с 2007 по 2008 гг. возросло количество рыб с нарушениями в структуре ядра эритроцитов (рис. 3.2 5), кариорексис, хроматинолиз, кариолиз. С 2006 по 2008 гг. возрастает количество (% особей рыб) со структурными нарушениями в эритроцитах: пойкилоцитоз, разноразмеренность. Такие нарушения эритроцитарной мембраны возникают при контакте с токсическими веществами, обладающими повреждающим действием.
Согласно данным токсикологической характеристики воды Волго-Каспийского канала, в период с 2006 по 2009 гг. было отмечено увеличение содержания в воде некоторых токсических компонентов: марганец, нефть и др. (Ежегодник. «Качество поверхностных вод», 2007; 2008; 2009).
Большинство показателей цитопатологических изменений в количественном отношении не выходили за пределы «условной нормы». Однако, большое разнообразие форм патологических изменений, связанных с проявлением хронического токсикоза в виде нарушений структуры ядра эритроцитов и высокое их общее (суммарное) количество, создаёт общую неблагоприятную картину физиологического состояния рыб.
3.3. Анализ физиологического состояния организма леща европейского (Abramis brama) Волго-Каспийского канала по гематологическим показателям и инвазированности паразитами
Исследование инвазированности особей леща Волго-Каспийского бассейна по сезонам года с 2006 г. по 2008 г., показало, что паразитические сообщества были представлены 16 видами гельминтов, относящихся к 6 классам, степень инвазии рыб во всех случаях соответствовала бессимптомному паразитоносительству. Ни в одном из исследованных случаев инвазия не сопровождалась наличием необратимых видимых патологических изменений. Интенсивность поражения рыб не достигала летального порога, однако почти все особи леща (весной 96 %) были заражены паразитами. Исследованием установлено существование корреляционной зависимости между паразитированием в организме леща гельминтов и изменением некоторых параметров периферической крови рыб (также как и для судака). Следовательно, паразитирование гельминтов в организме леща отражается непосредственно на изменении параметров периферической крови рыб.
Анализ состояния периферической крови исследованных особей леща осенью 2006 года обнаружил пониженное (недостаточное) содержание гемоглобина и количества эритроцитов в одном литре периферической крови особей леща (табл. 3.3.1.). У этих же рыб был отмечен относительно низкий уровень СОЭ. В 2007 году, как и в 2006, было отмечено пониженное содержание гемоглобина. Насыщение эритроцита гемоглобином (СГЭ) было достаточно низким. При этом наблюдался низкий уровень СОЭ у этих же рыб. Количество эритроцитов в литре крови находилось в пределах нормы. В 2008 году отмечено недостаточное содержание гемоглобина и уровень СОЭ. Количество эритроцитов в литре крови весной было незначительно выше условной нормы, а осенью - ниже. Преобладающими клетками в составе белой крови рыб являются лимфоциты, которые составляли у отдельных особей до 100 % всех лейкоцитов, но средний уровень их был в пределах нормы (Справочник…, 1986, Методические указания…, 1974, 1999; Эколого-гематологические характеристики, 1997). Общее количество лейкоцитов в литре крови леща в течение исследования было в пределах нормы. Однако, в лейкограмме выявлены существенные различия в составе популяций клеток. В период с 2006 по 2008 гг. отмечено понижение содержания псевдобазофилов в крови рыб, количество псевдоэозинофилов увеличилось с 2006 по 2008 гг., но всегда находилось в пределах нормы. На фоне снижения нейтрофилов из года в год и относительно условной нормы, происходило увеличение доли клеток лимфоидного ряда как относительно нормы, так и с 2006 по 2008 год.
Таблица 3.3.1.
Показатели крови, отражающие физиологическое состояние особей леща
| Показатели крови: | Условная норма | 2006 | 2007 | 2008 | Среднее по сезонам | Общее среднее M±m | |||
| Осень M±m | Весна M±m | Осень M±m | Весна M±m | Осень M±m | Весна M±m | Осень M±m | |||
| Hb, г/л | 92,0-101,0 | 75,9±3,8 | 79,7±2,2 | 88,1±4,3 | 82,8±3,0 | 82,8±4,6 | 81,2±2,6 | 82,3±4,2 | 81,8±3,4* |
| СОЭ, мм | 4,7 | 2,5±0,3 | 2,7±0,4 | 2,1±0,2 | 3,6±0,3 | 0,5±0,07 | 3,2±0,4 | 1,7±0,2 | 2,4±0,3 |
| Колич. эритроцитов *1012 в л | 1,36 | 1,11±0,11 | 1,35±0,04 | 1,38±0,08 | 1,51±0,1 | 1,1±0,09 | 1,43±0,07 | 1,20±0,09 | 1,31±0,08 |
| СГЭ, пг | 70,2 | 70,4±9,0 | 52,9±1,4 | 58,8±3,8 | 54,9±3,4 | 73,7±5,1 | 53,9±2,4 | 67,7±5,9 | 60,8±4,2 |
| Колич. лейкоцитов *109 в л | 45,0-120,0 | - | 65,5±7,7 | 80,3±9,9 | 72,7±7,4 | 77,4±11,0 | 69,1±7,5 | 78,8±6,9 | 74,0±7,3 |
| ПЭФ, % | 4,9 | 1,73±0,40 | 1,71±0,32 | 2,0±0,57 | 0,05±0,03 | 2,52±1,86 | 0,88±0,17 | 2,08±0,9 | 1,48±0,56 |
| ПБФ, % | | 10,38±1,2 | 2,62±0,69 | 0,9±0,27 | 0,6±0,14 | 0,12±0,06 | 1,61±0,41 | 3,8±0,51 | 2,70±0,46 |
| НФ, % | 5,5-14,4 | 4,96±0,69 | 7,5±1,25 | 3,0±0,4 | 3,0±0,44 | 2,8±0,56 | 5,25±0,84 | 3,6±0,5 | 4,41±0,70 |
| ЛФ, % | 81,0-89,0 | 82,3±1,67 | 86,59±1,6 | 93,7±0,64 | 95,6±0,5* | 93,7±1,93 | 91,1±1,07* | 89,9±1,4 | 90,5±1,2** |
| Молодые лейкоциты,% | - | 0,0 | 0,66±0,25 | 0,3±0,11 | 0,18±0,14 | 0,28±0,12 | 0,42±0,19 | 0,19±0,08 | 0,3±0,1 |
| Мн, % | 2,3-8,4 | 0,0 | 0,91±0,23 | 0,2±0,08 | 0,6±0,27 | 0,5±0,12 | 0,75±0,25 | 0,23±0,07 | 0,5±0,2 |
Содержание моноцитов (%) всегда находилось ниже условной нормы. Отмеченный лимфоцитоз – увеличение содержания лимфоцитов относительно всего пула клеток, на фоне монопении, а также снижение количества нейтрофилов и их малой функциональной активности, свидетельствует о неблагополучном состоянии организма, иммунологической недостаточности, что может быть спровоцировано хроническим воздействием токсических веществ или наличием инфекционных заболеваний неизвестной этиологии. Описанное физиологическое состояние особей леща нельзя оценивать как угнетенное. По-видимому, наблюдаемая картина крови с её отклонениями есть результат адаптации организма рыб к существованию в условиях хронического токсикоза и интенсивной антропогенной нагрузки.
Анализ патоморфологических изменений в структуре эритроцитов леща
- Патоморфологические изменения клеток красной крови леща соответствуют аналогичным нарушениям, обнаруженным в крови особей судака (табл. 3.3.2). Пойкилоцитоз эритроцитов был выражен, как и у судака, полиморфностью клеток. Эритроциты с отростками (характерные для угнетения эритропоэза), встречались единично, у отдельных особей. Осенью 2007 г. у 96% особей в крови рыб регистрировали эритроциты различной формы, но данное изменение формы вполне обратимо и может быть возвращено в нормальное состояние при изменившихся условиях.
- На мазках крови исследованных особей рыб отмечены случаи амитоза эритроцитов, находящихся на различных стадиях деления. Отдельно выделяли эритроциты с ядрами в стадии деления. Снижение частоты встречаемости данного признака отмечено от весны к осени, что может быть обусловлено физиологическим состоянием организма рыб. Доказано, что при токсическом воздействии происходит первая ответная реакция организма в виде увеличения концентрации гемоглобина в крови рыб. Повышение частоты встречаемости делящихся клеток путем амитоза приводит к увеличению их количества и, как следствие, к увеличению общего количества гемоглобина. Такая реакция эритроцитов может иметь компенсаторный характер и служить элементом адаптации организма рыб при токсикозах. Амитотическое деление эритроцитов может быть результатом воздействия повреждающих токсических веществ на ядерные структуры клетки. Разноразмеренность клеток красной крови отмечали у исследованных особей во все периоды исследования. Однако частота встречаемости данного показателя у отдельных рыб не превышала 4,3 % от общего пула эритроцитов, и отражало нормальный процесс гемопоэза. В крови рыб отмечали эритроциты, находящиеся на стадии разрушения.
- Таблица 3.3.2.
Типы цитопатологических нарушений в эритроцитах особей леща
| Основные типы нарушений | Количество особей (%) с нарушениями структуры клеток от общего количества обследованных рыб | ||||||
| 2006 | 2007 | 2008 | Среднее | ||||
| Осень | Лето | Осень | Лето | Осень | Лето | Осень | |
| Пойкилоцитоз | 68,0* | 65,0* | 96,0** | 100,0 | 88,0* | 83 | 84 |
| Анизоцитоз | 4,0 | 5,0 | 52,0* | 95,5** | 52,0* | 50 | 36 |
| Агглютинация | 40,0* | 15,0* | 4,4 | 41,0* | 4,0 | 28 | 16 |
| Изменение внешней плазматической мембраны | 12,0* | 30,0* | 52,0* | 73,0* | 88,0* | 52 | 51 |
| Смещение ядер к периферии | 0,0 | 100,0 | 61,0* | 95,5** | 80,0* | 98 | 75 |
| Пикноз | 60,0* | 5,0 | 48,0* | 95,5** | 44,0* | 50 | 51 |
| Деформация ядер | 36,0* | 90,0** | 48,0* | 86,0* | 76,0* | 88 | 53 |
| Кариорексис | 12,0* | 10,0* | 39,0* | 91,0** | 28,0* | 50 | 26 |
| Инвагинация ядра | 0,0 | 50,0* | 39,0* | 77,0* | 40,0* | 63 | 40 |
| Разрушение клетки | 0,0 | 15,0* | 96,0** | 95,5** | 72,0* | 55 | 84 |
| Ядра-тени | 0,0 | 0 | 44,0* | 100 | 88,0* | 50 | 66 |
| Хроматинолиз | 0,0 | 20,0* | 30,0* | 91,0** | 40,0* | 55 | 35 |
| Ядра-кольца | 0,0 | 0,0 | 0,0 | 45,0* | 80,0* | - | - |
| Протуберанцы | - | - | - | 36,0* | 24,0* | - | - |
| Вакуолизация цитоплазмы | 0,0 | 35,0* | 17,0* | 14,0* | 0,0 | 24 | 9 |
| Кариолиз | 0,0 | 5,0 | 26,0* | 68,0* | 32,0* | 37 | 29 |
| Шистоциты | 0,0 | 10,0* | 4,0 | 27,0* | 8,0* | 19 | 6 |
| Амитоз | 40,0* | 60,0* | 30,0* | 55,0* | 28,0* | 58 | 33 |
| Амитоз – деление только ядра | 0,0 | 70,0** | 40,0* | 77,0* | 40,0* | 74 | 40 |
| Количество исследованных рыб | 25 | 20 | 23 | 22 | 25 | 21 | 24 |
- У всех исследованных рыб в 2008 г. встречались эритроциты в предгемолитическом состоянии (95% весной и 72% осенью). Полностью разрушенные эритроциты, ядра-тени, отмечались у всех исследованных рыб весной и у 88% особей осенью 2008 г. Частота их встречаемости (% от общего количества исследованных эритроцитов), в среднем, составляла 1,1 % весной и 0,9 % осенью, и не превышала летального порога.
При исследовании крови леща (как и судака) регистрировали увеличение количества (%) особей с цитопатологическими нарушениями в весенний период (по сравнению с осенью). Снижение частоты встречаемости и интенсивности проявления цитопатологии к осени может свидетельствовать: о некоторой стабилизации токсикологических процессов водной среды в осенний период, о более благополучном физиологическом состоянии рыб после летнего нагульного периода. Это указывает на влияние сезонного фактора, на физиологическое состояние рыб. С 2006 по 2008 гг. возросло количество особей леща с патологическими нарушениями эритроцитов крови – пойкилоцитоз, анизоцитоз. Возросло с 2006 по 2008 г. количество структурных нарушений в ядре эритроцитов крови – хроматинолиз, кариолиз, кариорексис (рис. 3.3.1.).

Рисунок 3.3.1.
Уровень (средний %) цитопатологических отклонений в эритроцитах крови леща
Возросла доля (процентное содержание рыб в выборке) с нарушениями в структуре ядра эритроцитов периферической крови леща (так же как и судака). Доля (%) особей леща с присутствием явления кариорексиса в эритроцитах в 2006, 2007, 2008 гг. составила 12%, 24% и 59% соответственно, количество особей судака с нарушением в структуре ядра (кариорексис) в 2006 г. составило 20%, в 2007г. – 38%, в 2008 г. – 66%. Для периферической крови судака и леща с 2006 по 2008 гг. отмечено достоверное возрастание количества (%) особей с другими структурными нарушениями ядра эритроцитов (кариолиз, хроматинолиз)