
Физиологическая роль сосудистых рефлексогенных зон в интегративной регуляции функций дыхания и кровообращения 03. 00. 13 Физиология
| Вид материала | Автореферат |
СодержаниеСодержание работы Методы исследований. Результаты исследований и их обсуждение. Таблица 1. Общая характеристика, объем и методы исследования |
- Тесты рубежного контроля по разделам «физиология кровообращения и дыхания», 149.56kb.
- Физиологическая роль рецептора irr (insulin receptor-related receptor) в регуляции, 25.58kb.
- Бета-адренореактивные структуры в регуляции адаптационных возможностей системы кровообращения, 709.33kb.
- Игры, которые лечат, 31.3kb.
- Роль нейронов медиобазального гипоталамуса, синтезирующих дофамин, в центральной регуляции, 261.75kb.
- Пояснительная записка Нормальная физиология наука и учебная дисциплина о жизненных, 578.98kb.
- Урок 29 Дата проведения Тема: «Механизм вдоха и выдоха. Регуляция дыхания. Охрана воздушной, 58.3kb.
- Роль -эндорфина в нейроэндокринной регуляции функций иммунной системы 14. 00. 36 Аллергология, 657.6kb.
- Тема: Значение дыхания. Органы дыхательной системы: дыхательные пути, голосообразование, 62.1kb.
- Урок №38 «Значение дыхания. Органы дыхательной системы;», 101.56kb.
СОДЕРЖАНИЕ РАБОТЫ
Предмет и объект исследований. Опыты проводились на 229 взрослых кошках обоего пола массой 1,9 – 4,5 кг под внутривенным уретановым наркозом 1 г/кг массы животного при исходном артериальном давлении 110 – 130 мм рт. ст. У кошек в отличие от собак и кроликов редуцированы анастомозы между позвоночными и затылочными артериями (L.R. Muller, 1924; Л.А. Андреев, 1937; J.P. Whinsant et al., 1956; De la Torre et al., 1959, А.Д. Ноздрачев, 1973, и др.), что способствует гуморальной изоляции сосудов вертебрального бассейна. Особое внимание уделялось наркотизации животного с целью более полной стандартизации условий эксперимента. Вмешательства проводились во втором (легкий наркоз) и третьем (глубокий) периоде третьей стадии собственно наркоза. Уретановый наркоз в отличие от других видов меньше угнетает рефлекторные реакции дыхания и кровообращения. Он слабо влияет на процессы возбуждения и торможения в центральной нервной системе (М.Д. Машковский, 1993), синаптическую межнейронную передачу (И. Тасаки, 1957), периферические синапсы и не влияет на передачу возбуждения в вегетативных ганглиях (S.M. Hagiwara, N. Saito, 1959).
См. схему опыта на рис. 1.

Рис. 1. Общая принципиальная схема опытов
1, 2, В – система для перфузии СРЗ позвоночной артерии, изменения и регистрации давления в ней; 3 – специальная лигатура для зажатия артерии (позвоночной или сонной); 4, А – регистрация внешнего дыхания; 5 – ультратермостат; 1, 6 – система шприц-манометр, сосуд Мариотта; 7 – бедренная вена; 8 – ободочная вена; 9, Д – фото-аудиография объемной скорости перфузии; Б – отметчик времени; 10 – регистрация системного артериального давления.
Диаметр правой позвоночной артерии в среднем в 1,5 – 2 раза меньше левой (J.S. Stopford, 1915, 1916; H. Duret, 1973; Н.В. Верещагин, 1980), кроме того, она имеет меньше анастомозов с корешково-спинальными сосудами. По этим причинам в условиях острого опыта ее гемодинамическая изоляция оказывается предпочтительнее, чем левой.
Методы исследований. Для гемодинамической изоляции (на основе методики В.Н. Черниговского) в устье ПА, расположенном на уровне VI шейного позвонка (А.И. Акаевский, М.И. Лебедев, 1971; А.Д. Ноздрачев, 1973), у ее входа в канал поперечных отростков шейных позвонков ввязывалась приводящая канюля. Медиальнее от нее перевязывалась плечеголовная, латеральнее – подключичная артерии, щито-шейный и реберно-шейный стволы. Это исключало связь позвоночной артерии с веточками глубокой шейной и передней спинномозговой артериями. В области атлантоокципитального сочленения позвоночника, на уровне II шейного позвонка также проводилась перевязка позвоночной артерии. Лигирование щито-шейного ствола, а также позвоночной артерии на уровне II шейного позвонка устраняет связь с анастомозирующими веточками позвоночной артерии на всех ее экстракраниальных уровнях, кроме того, исключается связь экстракраниального отдела позвоночной артерии с коллатералями передней спинальной артерии, берущей начало от интракраниальной части позвоночной артерии. Для наложения лигатуры на позвоночную артерию на уровне II шейного позвонка использовалась модифицированная нами игла Дешана (рац. предл. №1012, 10.10.2000). Допустимо, что даже если небольшая часть жидкости через мелкие коллатеральные веточки может медленно проникать за пределы позвоночной артерии при пороговых изменениях давления в ней, то изменение давления в этих областях будет подпороговым, и афферентация с рецепторов ЗПА будет доминировать. Для выключения коллатералей и анастомозов артериол и капилляров корешково-спинальных сосудов шейного утолщения позвоночника дополнительно проводилась их эмболизация 10% взвесью ликоподия в вазелиновом масле по общепринятой методике, применяемой для выявления рефлекторных реакций (К. Гейманс, Д. Кордье, 1940; Д.И. Смирнов, 1954; Г.Н. Котова, 1957; С.В. Куприянов, 2007, и др.). Эффективность такой гемодинамической изоляции ранее подтверждена ангиографически (Ю.Г. Александров, 1989). В контрольных опытах для визуальной проверки полноты изоляции в ЗПА нагнеталась тушь.
Для изменения давления (повышения, снижения) в ЗПА в одних случаях приводящая канюля, совмещенная через тройник с манометром, соединялась с сосудом Мариотта (H.E. Hering, 1924; B. Oberg, 1977; В.М. Хаютин, Г.П. Конради, 1986; Б.И. Ткаченко с соавт., 1994; С.В. Куприянов, В.С. Куприянов, Л.М. Семенова, 2006). При других наблюдениях для этих целей использовалась система «шприц-манометр». Барорецепторы ЗПА стимулировались изменениями уровня расположения сосуда Мариотта или нагнетанием и декомпрессией шприцем за 10 с 0,5 мл раствора Рингера-Локка t 37С. Это вызывало статическое повышение или снижение давления в ЗПА. Отсутствие расхода раствора и примеси крови в системах, связанных с позвоночной артерией, указывали на полноту гуморальной изоляции. Повышение давления вышеуказанными способами осуществлялось на 30 – 50 мм рт. ст., что соответствует адекватной (физиологической) стимуляции барорецепторов СРЗ, возникающей, например, при физической нагрузке. Кроме того, в отдельных сериях снижение давления в ЗПА вызывалось окклюзией в ее начальном отделе. Метод окклюзии сосудов, питающих СРЗ, широко используется в экспериментальной физиологии (И. Навалихин, 1870; С. Heymans, J. Boukaert, 1933; В.Н. Черниговский, 1960; В.М. Хаютин, 1964; А.Д. Ноздрачев, 1973; Б.И. Ткаченко с соавт., 1994; С.В. Куприянов, Н.А. Агаджанян, 2005, и др.). Так, в начале окклюзии одной общей сонной артерии давление в ее каротидной зоне снижается на 50% от исходного уровня (D. Chungeharoen et al, 1952), что соответствует порогу возбудимости барорецепторов и возникновению прессорного рефлекса (В.М. Хаютин, 1964; C.Z. Pelletier et al, 1972; H.R. Kirchheim, 1976; Н.Н. Алипов с соавт., 2007). Поскольку величина давления в общих сонных и позвоночных артериях, их функция кровоснабжения головного мозга идентичны, то можно предположить, что порог возбудимости их барорецепторов примерно одинаков. Окклюзия одной позвоночной артерии была кратковременной (5 – 15 с) и вызывалась специальным зажимом нашей конструкции (рац. предл. №1005, 16.09.99). Такой зажим устраняет натяжение артерий при их зажатии и возникновение болевых реакций.
Для хемостимуляции исследуемых СРЗ использовались растворы D,L (2 оксипропионовой, С3Н6О3) молочной кислоты (0,5 – 3,0 ммоль/л), гидрокарбоната натрия (NaHCO3, 310 – 700 ммоль/л), три (гидроксиметил)-аминометана (трисамина, трисбуфера, 0,3 М). Указанные минимальные концентрации молочной кислоты (имевшие рН 7,34 – 7,06) соответствуют нормальному и повышенному содержанию лактата (естественного метаболита) в плазме крови. В отдельной серии экспериментов в качестве раздражителя использовалась венозная кровь, вводимая в ЗПА и каротидный синус методом аутогемоперфузии. Для изучения рефлекторного компонента терапевтического влияния инфузии озона использовался физиологический раствор с содержанием данного вещества от 400 до 1600 мкг/л. Барботирование физиологического раствора озоном осуществлялось по общепризнанной методике (О.В. Масленников, К.Н. Конторщикова, 2003; Е.Н. Сизова с соавт., 2003; V. Dvorak, 2005) с использованием аппаратов марки «Озон 3» производства МП «Озон» (Москва) или «Медозонс-БМ» (Н. Новгород). Определение концентрации озона в 200 400 мл физиологического раствора (Рингера-Локка) проводилось по времени барботирования. 10 минутная экспозиция создает концентрацию озона в растворе от 400 до 1600 мкг/л. В условиях клиники такой раствор используется для внутривенных введений (С.П. Алехина с соавт., 1996).
Во всех случаях ЗПА и каротидный синус подвергались предварительной гемодинамической изоляции. Перфузия гуморально изолированной позвоночной артерии (чаще правой) и каротидного синуса растворами хемоактивных веществ осуществлялась свободным током под постоянным давлением. Растворы вводились через канюлю, расположенную либо в месте отхождения позвоночной артерии от плечеголовного ствола, при этом на экстракраниальном уровне раствор свободно вытекал из позвоночной артерии в раневую область. Либо растворы вводились проксимальнее места бифуркации общей сонной артерии и покидали каротидный синус через надрез в начальном отделе наружной сонной артерии. Таким образом, перфузия исследуемых СРЗ не сопровождалась изменением давления в них и не приводила к активации их барорецепторов. Контролем в данном случае являлось введение в зоны позвоночных и сонных артерий физиологического раствора.
Так же, как и в случае активации барорецепторов, сила раздражения хеморецепторов находилась в физиологических пределах. Например, указанные концентрации молочной кислоты соответствуют нормальному и повышенному содержанию лактата в венозной крови и плазме, наблюдаемому в естественных условиях при умеренной, например, физической нагрузке (Е.А. Строев, В.Г. Макарова, 1986; Т.Т. Березов, Б.Ф. Коровкин, 2004; Е.С. Северин с соавт., 2005). 0,3 М раствор трисамина (3,66%) являлся стандартным, используемым в клинике для устранения ацидоза и повышения щелочного резерва крови (Р.А. Альтшулер, В.М. Балагин, Н.Б. Назарова, 1967; М.Д. Машковский, 1993).
Регистрация внешнего дыхания осуществлялась методом пнеймографии при трахеостомии с использованием капсулы Марея в нашей модификации с применением гофрированного сельфона, заменяющего резиновую мембрану. Такой сельфон не меняет своих эластических свойств при растяжении или спадании и позволяет проводить линейную регистрацию внешнего дыхания (рац. предл. №1004, 16. 09. 99). Системное артериальное давление регистрировалось ртутным или электроманометром (МЭП-И-01) с бедренной артерии.
Изучение роли барорецепторов ЗПА в регуляции тонической активности вен осуществлялось в условиях перфузии раствором Рингера-Локка (t 37 С) при p-const. (10 15 мм рт. ст.) бедренной или левой ободочной вены с захватом ее большой аркады со средней ободочной веной (далее в тексте – ободочная вена). Регистрация скорости и объема перфузата осуществлялась прибором нашей конструкции (рац. предл. №1006, 16.09.99, С.В. Куприянов), который обеспечивает одновременную графическую запись, а также аудио- и фотонаблюдение за характером перфузии. Перфузия под постоянным давлением физиологически оправдана для сосудов низкого давления. Учет изменения объема перфузата по частоте его капель в единицу времени позволяет точно судить об изменениях просвета исследуемых вен. Этот метод нашел широкое применение в опытах по изучению рефлекторных и гуморальных изменений тонуса лимфатических и емкостных сосудов в работах многих авторов (Д.И. Смирнов, 1954; Г.Н. Котова, 1957; В.В. Петровский, 1960; В.С. Куприянов, 1966; Д.П. Дворецкий, 1967; П. Джонсон, 1982; В.А. Левтов, А.Т. Матчанов, Н.А. Анисимова, 1991). Известно, что в венах отсутствуют пульсовые колебания крови, порождаемые vis a tergo вследствие работы сердца (Б.Ф. Вериго, 1905; Е.Б. Бабский с соавт., 1972; Р. Шмидт, Г. Тевс, 1986; К.В. Судаков, 2000). По мнению Г.П. Конради (1973), перфузия при р const. лишена важного недостатка, присущего резистографии – «… принудительного, насильственного проталкивания неизменного объема жидкости через сосуд, что может оказывать избыточное механическое давление на него».
В контрольных опытах все вышеописанные воздействия на хемо- и барорецепторы зон позвоночных и сонных артерий проводились после их новокаиновой блокады. Также в сериях экспериментов по изучению барорефлексов с этих СРЗ на вены скелетных мышц и спланхнического бассейна в качестве контроля проводилась фармакологическая и/или хирургическая денервация соответствующих эффекторных сосудистых областей.
Для стандартизации условий экспериментов ответные реакции регистрировались в начале каждого опыта, когда отрицательные влияния условий острого опыта имеют минимальную выраженность. Стимуляция хемо- и барорецепторов осуществлялась до момента проявления максимальной выраженности рефлексов КРС, после чего вмешательство прекращалось.
Обработка полученных данных проводилась в среде электронных таблиц “Excel” фирмы “Microsoft”, статистическая обработка результатов – с использованием пакета «Statistica». Оценку достоверности результатов проводили на компьютере IBM PC Pentium III по t-критерию Стьюдента или критерию знаков (Л.С. Каминский, 1974). Корреляционный анализ выраженности барорефлексов осуществлен методом наименьших квадратов (Д. Худсон, 1970), корреляционный анализ хеморефлексов – методом Пирсона.
Результаты исследований и их обсуждение. Общий объем исследований приведен в табл. 1.
Адекватная (соответствующая физиологическим величинам) стимуляция барорецепторов гуморально изолированной ЗПА повышением давления на 30 –50 мм рт. ст. вызывала достоверное снижение тонуса бедренной вены (34 наблюдения), что выражалось в увеличении объема перфузата в единицу времени. Одновременно происходило рефлекторное угнетение внешнего дыхания и снижение уровня САД.
Таблица 1. Общая характеристика, объем и методы исследования
| Серии опытов | Регистрируемые параметры, использованные методики | Число наблюдений (животных), n |
| Исследование барорецептивной активности зоны позвоночных артерий | 1) САД – окклюзионный способ; 2) внешнее дыхание – трахеостомический метод; 3) тонус периферических вен – перфузия (р-const.); 4) тонус вен внутренних органов – перфузия (р-const.). | 120 (32) |
| Проведение корреляционного анализа выраженности барорефлексов с зон позвоночных и сонных артерий | Метод наименьших квадратов | 21 (5) |
| Исследование хеморефлексов с зоны позвоночных артерий, сравнение их выраженности с хеморефлексами каротидных синусов | 1) САД – окклюзионный способ; 2) внешнее дыхание – трахеостомическая пнеймография. | 449 (102) |
| Корреляционный анализ выраженности хеморефлексов на внешнее дыхание и уровень системного артериального давления с зоны позвоночных артерий и каротидного синуса | Метод Пирсона (определение среднего квадратичного отклонения) | 50 (10) |
| Анализ межсистемных взаимодействий при реализации баро- и хеморефлексов с исследуемых сосудистых рефлексогенных зон | Коэффициент Хильдебранта (оценка межсистемных отношений) | 160 (23) |
| Контрольные наблюдения во всех сериях | Фармакологическая и/или хирургическая денервация | 127 (57) |
| Общее количество наблюдений (животных), n | 927 (229) |
Угнетение вентиляции легких происходило преимущественно за счет уменьшения амплитуды дыхательных движений. Реже уменьшение глубины дыхания сопровождалось снижением его частоты. В подавляющем большинстве случаев частота сердечных сокращений (ЧСС) оставалась неизменной, в отдельных случаях происходило ее уменьшение. О рефлекторной природе наблюдаемых реакций свидетельствует следующее. 1. Введение в позвоночную артерию 0,5 мл 2% раствора новокаина спустя 20 мин. устраняло все указанные реакции. 2. Во всех наблюдениях после вмешательства при возвращении уровня давления в ЗПА к исходной величине происходило синхронное возвращение регистрируемых показателей тонической активности вен задних конечностей, дыхания и САД к исходным или близким к ним параметрам. Это указывает на проявление периода последействия рефлекса. 3. Выраженность указанных рефлекторных реакций находилась в относительно прямой зависимости от силы раздражителя, а при равной величине стимула – от продолжительности раздражения барорецепторов. Это, по нашему мнению, является результатом проявления закона силовых отношений и суммации афферентации в центральной нервной системе, что также характерно для рефлекторных ответов.
Качественно однонаправленные реакции тонической активности вен, внешнего дыхания и уровня САД, возникающие в ответ на прессию в гемодинамически изолированной ЗПА, были получены в серии экспериментов при перфузии (p – const.) ободочной вены. Эти реакции также носили рефлекторную природу.
Для объективной оценки роли бароафферентации от ЗПА в регуляции тонической активности периферических и органных вен, а также уровня артериального давления и деятельности дыхательной системы были проведены эксперименты на 46 животных со снижением давления в изолированной позвоночной артерии.
Одновременно возникало сужение исследуемых вен, стимуляция внешнего дыхания и повышение САД. После прекращения вмешательств наступало восстановление исходного тонуса венозных сосудов, вентиляции легких и стабилизация общего кровяного давления. В ряде контрольных экспериментов проводилась первичная фармакологическая и хирургическая денервация бедренной вены, при которой описанные прессорные и депрессорные рефлексы на этот сосуд выпадали. При этом реакции с барорецепторов ЗПА на внешнее дыхание и САД сохранялись, а исчезали только после новокаиновой блокады области ПА. Контрольные вмешательства доказывают рефлекторное происхождение этих реакций.
Разнонаправленная стимуляция барорецепторов ЗПА (де- и прессорная) вызывает противоположные рефлекторные реакции исследуемых функций. Например, локальное снижение давления в позвоночной артерии приводит к повышению САД, тонуса ободочной вены и стимуляции дыхательной системы. Подобные рефлексы, но только на внешнее дыхание и общее кровяное давление (без сочетанных с ними реакций емкостных сосудов), были продемонстрированы автором ранее и изложены в диссертации на соискание степени кандидата медицинских наук (С.В. Куприянов, 2000).
Какова возможная целесообразность полученной в настоящей работе однонаправленности рефлексов на емкостный и резистивный бассейны? В чем значение одновременных с ними реакции внешнего дыхания?
Обобщение приведенных здесь данных с более ранними результатами исследований нашей лаборатории по изучению барорецептивной активности показало следующее. Во-первых, вены скелетных мышц и внутренних органов не являются пассивными трубками, проводящими кровь к сердцу, они способны к самостоятельной тонической деятельности. Во-вторых, одним из механизмов регуляции тонуса этих сосудов является бароафферентация от СРЗ позвоночных артерий. В третьих, прессорная или депрессорная активация барорецепторов ЗПА вызывает однонаправленные изменения тонуса резистивных и емкостных сосудов. Физиологическое значение такого однонаправленного изменения их тонуса заключается, очевидно, в адекватной компенсации исходных изменений интенсивности кровообращения на уровне тканей. Например, депрессорное раздражение барорецепторов СРЗ наблюдается при исходном снижении САД в целостном организме. При этом различные периферические ткани и органы оказываются в состоянии гипоксии и гиперкапнии. Компенсация такого состояния возможна рефлекторным повышением уровня САД и уменьшением емкости венозного бассейна. Последнее необходимо для перемещения депонированной крови через сердце в магистральные артерии и может быть достигнуто повышением тонуса вен скелетных мышц и внутренних органов. Все эти рефлексы артерий и вен зарегистрированы в настоящей работе. Одновременно происходит рефлекторная гипервентиляция, обеспечивающая оптимальную, большую оксигенацию крови. Противоположные, но также однонаправленные изменения тонуса резистивного и емкостного русла наблюдаются при исходном повышении давления в большом круге и, очевидно, направлены на рефлекторное компенсаторное угнетение интенсивности кровотока на уровне тканей. Приспособительное рефлекторное снижение давления в большом круге кровообращения уменьшает поступление крови в венозный бассейн, а расширение вен предотвращает избыточное перемещение крови из вен в сердце. При этом уменьшается и амплитуда дыхательных движений. Все эти совместные реакции, протекая однонаправленно, способствуют сохранению нормального кровотока и оптимального газообмена в тканях организма. Обоснованным будет предположение, что и другие СРЗ, например, синокаротидная или аортальная, способны однонаправленно изменять тонус резистивного и емкостного руслов большого круга кровообращения.
Таким образом, барорецепторы ЗПА обеспечивают сочетанную однонаправленную регуляцию внешнего дыхания, САД и тонической активности емкостных сосудов различных отделов большого круга кровообращения. Такая одновременная реакция различных систем организма, направленная на формирование общего конечного полезного результата, характерна для функциональных систем (П.К. Анохин, 1975; К.В. Судаков, 1987, 1997, 2000). Возникает вопрос о существовании взаимозависимости между выраженностью этих реакций. Для ответа был проведен корреляционный анализ выраженности изменений амплитуды дыхательных движений с выраженностью изменений САД, возникающих при барорефлексах с позвоночной и синокаротидной зон. Выявление корреляции этих показателей представляет интерес для физиологии и патофизиологии, поскольку является критерием, указывающим на объединение реакций дыхательной и сердечно-сосудистой систем в единую функциональную систему. Деятельность этой функциональной системы проявляется не только в норме, но и при патологии.
Для корреляционного анализа были использованы данные, изложенные в диссертации автора на соискание степени кандидата медицинских наук, дополненные результатами опытов, описанных в настоящей работе выше. Проанализировано 21 наблюдение, сделанное при поочередном полном зажатии на 10 с правых позвоночной и сонной артерий. Определялось отношение изменений реакции внешнего дыхания (амплитуды волн пнеймограммы), выраженных в процентах к исходному уровню (А/А0, %), к относительному изменению САД, выраженному в мм рт. ст. (р, мм рт. ст.). Статистическая обработка выраженности реакций внешнего дыхания и САД проводилась по методу наименьших квадратов (Д. Худсон, 1970). Этот метод позволяет при разбросе экспериментальных точек, характеризующих в наших наблюдениях выраженность реакций внешнего дыхания и САД (Xi; Yi), в предположении существования линейной зависимости между ними (Y=bX), опытным путем вычислить значение b, определить доверительный интервал, провести единственную прямую, наиболее близко расположенную ко всем этим точкам, и оценить степень достоверности рассматриваемого предположения. Значение b определяется из условия df/db=0, где функция
n
f = Σ (Yi – bXi)2.
i=1
Следовательно,
n n
b = Σ Xi·Yi/ Σ Xi2.
i=1 i=1
Анализ показал наличие корреляции между изменениями амплитуды дыхательных движений и уровнем САД, носящей в определенном пределе линейную зависимость. При барорефлексах с обеих зон более выраженные изменения одного из рассматриваемых параметров соотносились с большей выраженностью изменений другого, и наоборот.
Выявленная линейная зависимость между выраженностью рефлекторных изменений внешнего дыхания и САД, реализующаяся с барорецепторов позвоночной и синокаротидной зон, доказывает функционально-системную взаимосвязь дыхания и САД. Другими словами, бароафферентация от исследованных СРЗ является фактором формирования функциональной КРС.
Хеморецепторы СРЗ, как и их барорецепторы, участвуют в срочной (нервной) регуляции дыхания и кровообращения, обеспечивая сохранение постоянства напряжения О2 и СО2 в крови и тканях организма и их рН, а также уровня САД (В.Ф. Маркелова, А.М. Уголев, 1960; Г.Г. Исаев, 1983; M. D. de Burgh, 1985; В.М. Хаютин, Г.П. Конради, 1986; Н.А. Агаджанян с соавт., 1987; Р.Ш. ГабдрахмАнов с соавт., 1990; L. Sinoway, S. Prophet, 1990; R.C. Fitzerald, 2000; Ю.Ю.Орлова с соавт., 2005; С.Г. Кривощеков, 2006, и мн. др.). Поэтому в следующих сериях экспериментов изучалась хеморецептивная активность зоны позвоночных артерий.
Была проведена перфузия изолированной ЗПА растворами молочной кислоты (0,5 – 3,0 ммоль/л). При этом зарегистрировано достоверное увеличение уровня САД и стимуляция внешнего дыхания. Полученные данные качественно соответствуют подобным результатам других авторов, изучавших хеморецептивную активность классических рефлексогенных зон (В.Ф. Маркелова, А.М. Уголев, 1960; Г.Г. Исаев, 1983; W.F. Nolan et al, 1985; M. D. de Burgh, 1985; В.М. Хаютин, Г.П. Конради, 1986; Р.Ш. Габдрахманов с соавт., 1990; L. Sinoway, S. Prophet, 1990; R.C. Fitzerald, 2000; С.Г. Кривощеков с соавт., 2006, и мн. др.). При активации хеморецепторов изолированной ЗПА растворами молочной кислоты различных концентраций наблюдается следующая выраженность реакций дыхательной и гемодинамической систем: повышение уровня САД – 18,85+9,61 мм рт. ст.; увеличение ЧСС – на 4,62+3,34% от исходного (до вмешательства) показателя; увеличение амплитуды внешнего дыхания – на 156,09+74,77% от исходной величины; повышение частоты дыхательных движений – на 26,53+25,38% от исходной. Следует отметить, что направленность рефлексов на дыхательную и сердечно-сосудистую системы не зависела от концентрации молочной кислоты в перфузируемом растворе. Количественный анализ реакций показывает, что выраженность рефлексов находится в прямо пропорциональной зависимости от концентрации раствора. При содержании молочной кислоты равной 3,0 ммоль/л изменения во внешнем дыхании и САД были больше, чем те же показатели – при использовании раствора в 0,5 ммоль/л.
Большой разброс показателей изменения ЧСС и частоты дыхания (ЧД), очевидно, свидетельствует о нестабильности реакций частотных характеристик, наблюдающихся при реализации хеморефлексов в конкретных условиях наших экспериментов. В данном случае более реактивными и стабильными при воспроизведении оказываются рефлексы, направленные на увеличение уровня САД и амплитуды дыхательных движений. Рефлекторное увеличение объема вентиляции легких происходило в основном за счет увеличения амплитуды дыхательных движений, в меньшей степени (реже) – их учащения. Изменения в деятельности сердечно-сосудистой системы также происходили в основном за счет повышения уровня САД и в меньшей степени за счет увеличения ЧСС. Подобная большая реактивность амплитуды внешнего дыхания и уровня САД в сравнении с частотой дыхания и сердечных сокращений наблюдалась во всех сериях наших экспериментов как по изучению баро-, так и хеморецептивной активности СРЗ. Очевидно, что при данных условиях рефлекторные реакции изменения частоты дыхания и работы сердца не являются основным способом достижения приспособительного результата со стороны дыхательной и сердечно-сосудистой систем.
Качественно такие же рефлексы, но менее выраженные, были зарегистрированы при введении в гемодинамически изолированные позвоночные и сонные артерии венозной крови, взятой у того же животного. Было произведено 43 наблюдения на 9 животных (табл. 3). В 36 случаях (р<0,01) регистрировалось повышение уровня САД. Одновременно в 33 наблюдениях (р<0,01) происходила стимуляция внешнего дыхания, которая во всех случаях была результатом увеличения его амплитуды. Из них в 15 наблюдениях также увеличивалась и частота дыхательных движений. В 12 случаях из общего количества опытов повышение активности сердечно-сосудистой системы происходило не только за счет поднятия уровня общего кровяного давления, но и за счет увеличения ЧСС, при этом в редких наблюдениях происходило и увеличение пульсового давления.
Новокаиновая блокада области позвоночной артерии устраняла все указанные реакции, что убедительно доказывает их рефлекторный характер.
Введение венозной крови в область позвоночной артерии моделирует развитие физиологического (в пределах нормы) ацидоза в целостном организме. В этом случае общий приспособительный результат достигается системами дыхания и гемодинамики прежде всего за счет значительного увеличения на 88,06+29,44% амплитуды внешнего дыхания, в меньшей степени – за счет повышения на 9,67+3,17 мм рт. ст. уровня САД, а также в некоторых случаях увеличением частоты сердечных сокращений и дыхания. Однако, изменение двух последних параметров происходит в малом количестве наблюдений и не может быть подвергнуто достоверной статистической обработке. Как сказано выше, факт нестабильности реализации рефлексов на частоту дыхания и сердечной деятельности говорит о том, что этот вид кардиореспираторных реакций в данных условиях не является основным способом достижения общего конечного результата. Очевидно, что при умеренной ацидотической активации повышению частотных характеристик организм предпочитает реакции увеличения глубины дыхания и уровня САД. Возможное объяснение этого факта будет дано ниже.
Стимуляция активности дыхательной и сердечно-сосудистой систем наблюдалась и при введении в СРЗ позвоночных и сонных артерий раствора гидрокарбоната натрия в концентрациях 310 – 700 ммоль/л. В этой серии осуществлено 207 наблюдений на ЗПА и 7 вмешательств на зоне каротид. Рефлекторные реакции стимуляции внешнего дыхания происходили в основном за счет увеличения его амплитуды (175 случаев). Иногда (90 случаев из 207) эта реакция сопровождалась также увеличением частоты дыхания, которая, однако, чаще оставалась без изменений (113 из 207). Одновременно с усилением дыхания достоверно и функционально однонаправленно происходила интенсификация гемодинамики, чаще выражавшаяся в повышении уровня САД (189 случаев). В 66 наблюдениях также увеличивалась ЧСС. В подавляющем количестве опытов (126 наблюдений) этот показатель работы сердца не изменялся.
Описанные рефлексы оказываются качественно однонаправленными с реакциями, возникающими при активации рецепторов ЗПА растворами молочной кислоты. На первый взгляд, эти данные следует признать как минимум любопытными, ведь гидрокарбонаты – фактически являются «химическими антагонистами» молочной кислоты. Логично ожидать, что их влияния на хеморецепторы СРЗ должны быть разнонаправленными. Объяснение этому «феномену» дано ниже.
В следующей серии экспериментов производилась активация хеморецепторов ЗПА раствором трисамина (трисбуфера) – вещества, обладающего основной буферной активностью, т.е. поддерживающего слабощелочную рН среды. Было произведено 40 вмешательств на 10 животных. При сдвиге рН среды в гуморально изолированной рефлексогенной ЗПА в щелочную сторону реализуются реакции одновременного сочетанного угнетения внешнего дыхания и снижения уровня САД. Угнетение внешнего дыхания происходит преимущественно за счет уменьшения амплитуды дыхательных движений (33 случая из 40 наблюдений), при этом глубина его уменьшается более чем на 1/3 (в среднем на 38,52+10,95%; р<0,01) от исходного уровня. В некоторых случаях (8 наблюдений) происходит также уменьшение частоты дыхания, однако этот параметр работы дыхательной системы чаще (27 наблюдений из 40) остается неизменным. При этом уровень САД снижается (39 случаев из 40 наблюдений) в среднем на 15,27+5,62 мм рт. ст. (р<0,01). В 2 случаях также происходило уменьшение ЧСС, однако в подавляющем большинстве наблюдений (37 случаев) этот параметр оставался стабильным. При одном вмешательстве произошло его незначительное увеличение. В 6 опытах было зарегистрировано увеличение глубины дыхания и в 5 случаях увеличение его частоты (малое количество подобных наблюдений не нарушает достоверности).
Все описанные реакции носят рефлекторный характер, выявляемый контрольными вмешательствами вышеописанными методами. Также за рефлекторное происхождение этих реакций систем дыхания и кровообращения свидетельствует ряд косвенных признаков: во-первых, короткая продолжительность латентного периода всех указанных реакций, которая составляла 0,5-2,5 с; во-вторых, наличие, кроме латентного, также периодов развития максимальной выраженности ответа и последействия, характерных для рефлекторных ответов; в третьих, жесткая зависимость продолжительности реакций от времени действия раздражителя, после прекращения перфузии ЗПА раствором трисамина внешнее дыхание и САД стремятся вернуться к исходным или близким к ним величинам; в четвертых, следует учитывать, что одним из признаков хеморефлексов является удлиненность периода последействия, что также наблюдалось в наших экспериментах.
Угнетение внешнего дыхания, осуществленное только за счет снижения амплитуды дыхательных движений, происходило, как было сказано выше, более чем на 1/3 исходной величины. Снижение системного артериального давления – в среднем на 15,27+5,62 мм рт. ст. В наших экспериментах афферентация поступала только от рецептивного поля одной артерии. Логично предположить, что в целостном организме выраженность данного рефлекса будет больше, так как импульсация будет поступать и от второй артерии этой СРЗ, а также от других зон, деятельность которых аллиирована. На основании вышесказанного можно сделать вывод, который не выдвигается на защиту в рамках настоящей диссертации. При реализации афферентации от различных СРЗ приоритетное значение имеет та, которая сигнализирует ЦНС об отклонении гомеостатического показателя от нормы. При этом дыхательный и сердечно-сосудистый центры запускают реализацию приспособительных рефлексов даже в том случае, если от других СРЗ поступает информация о неизменности гомеостаза. Очевидно, в этом случае выраженность рефлекса будет меньше, чем при суммации однотипной афферентации от нескольких рефлексогенных зон.
Какова целесообразность описанных хеморефлексов? Общепризнано, что конечной целью функционально сочетанного взаимодействия систем дыхания и гемодинамики, т.е. результатом кардиореспираторных реакций, является регуляция газообмена в тканях и их КОС (D.M. Grenfield, 1964; H.P. Koepchen, 1983; Ф.Г. Ситдиков, С.И. Русинова, 1992; Н.А. Агаджанян с соавт., 2003; В.М. Покровский с соавт., 2003; Ж.А. Донина с соавт., 2006; И.В. Мухина с соавт., 2007; В.Ф. Пятин с соавт., 2007; А.И. Елфимов с соавт., 2007, и мн. др.). В наших экспериментах прессорная реакция, сопровождающаяся стимуляцией внешнего дыхания и наблюдающаяся в ответ на введение в исследуемые зоны молочной кислоты, очевидно направлена на усиление кровотока в тканях, компенсацию развивающего в них ацидоза, увеличение доставки О2 и удаление СО2. Противоположные реакции угнетения внешнего дыхания и снижения уровня САД наблюдаются при развитии алкалоза. Это состояние в наших экспериментах моделировалось введением в зоны позвоночных артерий или каротидного синуса раствора тисамина (трисбуфера). Афферентация, возникающая в этом случае от хеморецепторов, реализует рефлексы, направленные на снижение интенсивности кровотока в тканях. При этом изменения в деятельности дыхательной и сердечно-сосудистой систем направлены на достижение общих результатов: компенсаторное развитие на уровне тканей гипоксии, гиперкапнии и накопление в них кислых продуктов. Логично считать, что конечным результатом кардиореспираторных реакций в этом случае будет компенсация исходного защелачивания тканей.
Как объяснить однонаправленность реакций дыхательной и сердечно-сосудистой систем при активации хеморецепторов исследуемых зон как молочной кислотой, так и раствором стандартного гидрокарбоната (NaHCO3)? Следует учитывать, что кровь, как и межклеточная жидкость, является буферным раствором. Упрощенная формула гидрокарбонатного буфера может быть представлена следующим образом:
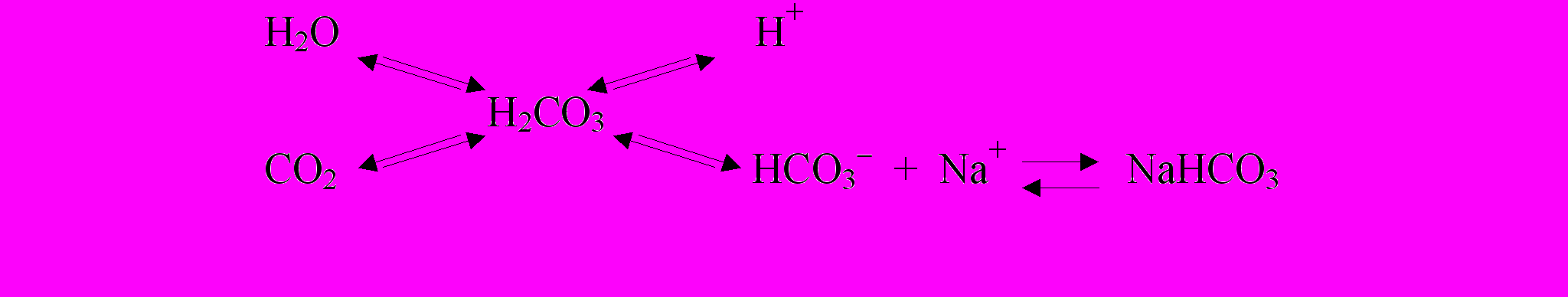
Становится понятным, что в отсутствии эффективных механизмов удаления H2O и CO2, введение в эту формулу любого одного из составляющих приведет к увеличению содержания всех других элементов буфера. В частности, при экзогенном увеличении концентрации NaHCO3 в изолированной СРЗ увеличивается и CO2. Отсутствие эффективных механизмов его удаления способствует постоянному высвобождению все новых (пусть даже и в малом количестве) порций протонов, которые вступают во взаимодействие с хеморецепторами ПА. Очевидно, СРЗ регистрируют изменения рН крови именно по содержанию в среде протонов и концентрации CO2, так как существование рецепторов на основания (в данном случае HCO3ˉ) многими авторами отрицается (И.С. Бреслав, В.Д. Глебовский, 1981; Б.И. Ткаченко с соавт., 1992). Следовательно, на введение стандартного основания в среду, всегда богатую протонами, организм способен реагировать, как на ацидотическое воздействие.
Трисамин не является естественным буфером организма. Он (трисбуфер), участвуя в реакциях обмена, непосредственно связывает протоны, включая их в свою молекулу (М.Д. Машковский, 1984). Поэтому при введении трисамина в организме происходит не только повышение рН, но и фактическое уменьшение концентрации протонов, основного закисляющего вещества в организме.
Наиболее частыми причинами развития метаболического ацидоза в целостном организме являются нарушения выведения воды почками или углекислого газа через легкие. Хорошо известно, что при попытке компенсации ацидоза введением солевых растворов, после краткосрочного уменьшения кислотности крови, зачастую развивается еще большее, по сравнению с исходным состоянием, снижение рН. Из вышесказанного следует: в клинике компенсация развития ацидоза должна проводиться только препаратами, подобными трисамину/трисбуферу, которые непосредственно связывают ионы водорода и в дальнейшем препятствуют их высвобождению. Использование с этой целью солевых растворов гидрокарбонатов Na, K, Ca, фосфатных солей следует признать не только неэффективным, но даже опасным. В условиях выраженного (клинически значимого) ацидоза их введение в конечном итоге будет способствовать увеличению концентрации Н+ и может восприниматься организмом как дополнительный, чрезмерно избыточный закисляющий фактор.
Широкое использование в практической медицине озона определяет целесообразность более детального изучения его влияния на организм человека. В клинике используются такие его свойства, как бактерицидное (фунгицидное и вирицидное), обезболивающее, дезинтоксикационное, активирующее кислородозависимые процессы, оптимизирующее про- и антиоксидантные системы, гемостатическое и антикоагулянтное, иммуномодулирующее. Однако механизмы, посредством которых реализуются эти эффекты, изучены на настоящий момент недостаточно и объясняются исключительно его прямым действием на ткани или чужеродные агенты (E.-B. Haddad et al., 1996; V. Bocci, 1997; А.М. Иваненко, А.В. Орлов, 1999; Е.Н. Сизова с соавт., 2003; И.Д. Муратов, 2005; V. Dvorak, 2005, и мн. др.). В доступной литературе не удалось обнаружить сведений о влиянии озона на центральные механизмы регуляции гомеостаза и, в частности, на активность таких жизненно важных систем, как дыхательная и сердечно-сосудистая. Отсутствуют сведения об участии в обеспечении подобных реакций СРЗ, в том числе ЗПА.
На 18 животных проведено 68 наблюдений при перфузии ЗПА озонированным физиологическим раствором и 10 вмешательств на каротидах. В 31 случае реализовывались сочетанные реакции снижения уровня САД и угнетения внешнего дыхания. В 8 – реакции дыхательной и сердечно-сосудистой систем были разнонаправлены. Во всех случаях депрессорная реакция и уменьшение амплитуды дыхания всегда проявлялись в начале опыта. При повторных введениях озона в одну и ту же СРЗ позвоночных артерий (еще 20 вмешательств) реакции этих двух систем организма становились нестабильными с преобладанием прессии и стимуляции внешнего дыхания. Спустя 20 мин. после введения в зону позвоночных артерий 0,5 мл 2% ого раствора новокаина все описанные реакции не воспроизводились (9 наблюдений), что указывает на их рефлекторную природу.
Во всех сериях экспериментов, как по изучению баро-, так и хемоактивности СРЗ, выраженность рефлексов оказывалась больше при активации хеморецепторов зоны каротидных синусов. В данной серии введение озонированного раствора в нее вызывало падение уровня САД на 20,83+2,17 мм рт. ст. и уменьшение амплитуды дыхательных движений на 92,0+9,6% от исходной величины. Частота дыхания и частота сердечной деятельности становились реже только в некоторых случаях. В 2/3 наблюдений при первичных введениях озонированного физиологического раствора частота внешнего дыхания и в 83,33% наблюдений частота работы сердца оставались неизменными. В остальных случаях происходило уменьшение обеих частотных характеристик. Не было зарегистрировано ни одного случая увеличения частотных характеристик работы дыхательной и сердечно-сосудистой систем.
При перфузии озоном ЗПА выраженность указанных кардиореспираторных реакций уменьшалась на 10,1+1,12 мм рт. ст. и 50,46+6,98% соответственно. Частота дыхания зачастую (в 40% случаев) оставалась неизменной. Также в 70% наблюдений неизменной была частота работы сердца. В остальных случаях первичных перфузий она (частота) снижалась. При многократных введениях озонированного раствора в одну и ту же позвоночную артерию или каротидную зону направленность и выраженность реакций становились неоднозначными с большим разбросом их численных выражений.
Таким образом, при первичной активации хеморецепторов зон позвоночных и сонных артерий раствором озона реализуются рефлексы угнетения внешнего дыхания и подавления гемодинамики, которые выражаются преимущественно уменьшением амплитуды дыхательных движений и снижением уровня САД. Урежение работы сердца и легких в данном случае не является основным рефлекторным механизмом достижения приспособительного результата. В опытах на одном животном при многократном введении озона в одну и ту же зону позвоночных артерий или каротидного синуса данная зависимость нарушается, направленность и выраженность рефлексов приобретают значительный разброс с преобладанием прессорного ответа.
Каков механизм и какова физиологическая целесообразность выявленных рефлексов на САД и дыхание при активации хеморецепторов зон позвоночных и сонных артерий озоном Известно, что молекула озона в растворе способна диссоциировать на свободный и атомарный кислород. Свободный О2 воздействует на хеморецепторы СРЗ, вызывая рефлекторное снижение САД и угнетение внешнего дыхания, т.е. реакции, противоположные действию на них СО2. Депрессорное влияние О2 на САД и дыхание с хеморецепторов классических СРЗ общеизвестно. Целесообразность такой депрессии связана с необходимостью уменьшить поступления О2 в ткани (Ф. Гродинз, 1966; А. Гайтон, 1969; Т.Т. Березов, Б.Ф. Коровкин, 2004), т.к. избыточное напряжение О2 в клетках отрицательно влияет на обмен веществ (Чандан К. Сен, Осмо Хеныинен, 1995; Minuz Pietro et al., 2006). Описанные нами рефлексы дыхательной и сердечно-сосудистой систем приводят к снижению поступления излишнего количества О2 в ткани. При повторных на одном животном перфузиях исследуемых сосудистых зон озоном депрессорный рефлекс сменялся прессорной реакцией САД. Эта прессорная реакция устранялась введением новокаина в зону ПА. Как можно объяснить эту смену ответов Следует учитывать, что нарастающее при длительном использовании озона увеличение концентрации освобождающегося атомарного кислорода должно оказывать альтеративное влияние на хеморецепторы. Кроме того, недиссоциированный озон не является естественным стимулом для хеморецепторов организма. Он лишен специфической точки воздействия на организм. По данным литературы, повышенная концентрация атомарного О2 и недиссоциировавшего озона в жидких средах организма ведет к проявлению их агрессивных токсических свойств, которые нарушают целостность мембраны клеток организма, в первую очередь, исследуемых нами хеморецепторов, вследствие повышенного окисления их фосфолипидов и липопротеидов, повреждения полипептидных цепей и протеидов. Озон в больших концентрациях и при длительной его экспозиции активирует перекисное окисление липидов мембраны эритроцитов. Е.Н. Сизова (2003) наблюдала сокращение гладких мышц изолированной полоски коронарных сосудов, мышц матки, бронхов животных при прямом продолжительном действии на них растворов с повышенным содержанием озона. В ситуации длительного повторного введения озона в зону позвоночных артерий (или каротидных синусов) с хеморецепторов реализуется альтеративное неспецифическое влияние. Реакция организма на фактически болевой раздражитель всегда проявляется однотипно, в частности, повышением САД и стимуляцией внешнего дыхания.
Полученные результаты соответствуют данным подобных исследований ряда других авторов. Например, И.В. Мухиной с соавт. (2007) показана дозозависимость влияния парентерального введения озонированных физиологических растворов на динамику поведения и структуру головного мозга крыс. Так умеренное (1,7 мкг/л) использование данных растворов приводило к увеличению двигательной и ориентировочно-исследовательской деятельности. При многократном и длительном применении озона в высоких концентрациях (2,5 мкг/л) развивались его токсическое влияние, угнетение активности животных, нарушение структуры неокортекса и отек головного мозга. Таким образом, характер действия озона (как и любого лекарственного вещества, применяемого с лечебной целью) связан с его концентрацией в организме. Любое биологически активное соединение в зависимости от его дозы может оказывать лечебное и токсическое действие. Действие озона носит как местный гуморальный характер, хорошо описанный в литературе, так и общий рефлекторный, продемонстрированный в настоящей работе и оказывающий опосредованное влияние через СРЗ и ЦНС.
Обобщение данных всех вышеописанных серий экспериментов, а также результатов, изложенных в диссертации автора на соискание степени кандидата наук, позволило установить целесообразность сочетанного проявления баро- и хеморефлексов, возникающих с СРЗ позвоночных артерий и каротидного синуса. В целостном организме баро- и хеморецепторы СРЗ функционируют в тесном единстве по принципу обратного взаимовлияния (В.Ф. Молчатская, 1990; D. Heisted Donald et al., 1995). Сочетанные рефлексы с барорецепторов на внешнее дыхание и САД сопряжены с последующими изменениями газового состава крови, которые влияют на активность хеморецепторов. И наоборот, исходное изменение химизма крови вызывает такие хеморефлексы, которые приводят к активации барорецепторов СРЗ (Е.И. Малыгина, 1961; D.В. Kаtzin, Е.Н. Rubinstein, 1974; Г.А. Вакслейгер, 1978; Р.Ш. Габдрахманов с соавт., 1990). О важности изучения роли совместной механо- и хеморецептивной активности в регуляции деятельности дыхательной и сердечно-сосудистой систем говорится в работах многих авторов (В.М. Хаютин, 1964; B. Folkow, E. Neil, 1971; Г.П. Конради, 1973; S. Hilton, 1982; В.М. Хаютин, Г.П. Конради, 1986; R.C. Fitzgerald, 2000; L. Glass, 2001; С.Г. Кривощеков с соавт., 2005, 2006).
При сопоставлении данных всех серий экспериментов выявляются следующие зависимости. Во-первых, порог раздражимости хеморецепторов СРЗ меньше, чем у барорецепторов. Во вторых, средняя выраженность хеморефлексов больше, чем барорефлексов. Следовательно, не только порог возбудимости хеморецепторов СРЗ ниже, чем у барорецепторов, но и функциональная активность первых выше. Во всех других сериях наших экспериментов наблюдалась подобная же зависимость, которая характеризуется большей хеморецептивной активностью исследованных СРЗ в сравнении с активностью их барорецепторов. В третьих, при реализации барорефлексов в наших экспериментах выраженность изменения внешнего дыхания и уровня САД была приблизительно в три раза меньше, чем их выраженность при реализации этими системами организма хеморефлексов (рис. 2). Наличие такой достаточно жесткой корреляции косвенно подтверждает их функционально системное взаимодействие, показывая объединение дыхательной и сердечно-сосудистой систем в единую функциональную систему (кардиореспираторную). В четвертых, кардиореспираторные реакции в определенных условиях реализуется в основном за счет изменений амплитуды внешнего дыхания и уровня САД. При этом частотные характеристики (внешнего дыхания и работы сердца) оказываются менее реактивными или же остаются неизменными. Под этими определенными условиями, по нашему мнению, следует понимать силу баро- и хемораздражителей, которая в экспериментах находилась в пределах физиологических границ, характерных для существования организма при умеренной (не чрезмерной) нагрузке.
По своей хемо- и барорецептивной активности ЗПА, по аналогии с другими СРЗ, участвует в рефлекторном контроле уровня САД и интенсивности внешнего дыхания. Этот контроль осуществляется с одновременным участием сердечно-сосудистого и дыхательного центров. При развитии, например, гипоксии вначале повышается функциональная активность хеморецепторов СРЗ, которая направлена на увеличение вентиляции легких и оксигенацию крови. Этому также способствует одновременное повышение САД, увеличивающее доставку крови в легкие. Затем, когда газообмен в тканях организма нормализуется, повышается активность барорецепторов, которая обеспечивает снижение возрастающего уровня САД до нормы. Такое взаимодействие дыхательного и сердечно-сосудистого центров, имеющее рефлекторную природу, обеспечивает сохранение гомеостаза газообмена и обмена веществ в тканях организма. Очевидно, что данные рефлекторные реакции носят системно функциональный характер. По мнению К.В. Судакова (2000), одним из основных способов доказательства функционально-системного объединения реакций является демонстрация их корреляции. Нами проведен корреляционный анализ выраженности барорефлексов на внешнее дыхание и уровень САД с ЗПА и каротидного синуса. Для этого проведена статистическая обработка результатов 50 наблюдений, взятых из разных серий вышеописанных опытов. Выборка осуществлена случайным образом.
Прежде всего, был разработан метод расчета выраженности изменений внешнего дыхания при его регистрации в остром эксперименте способом трахеостомической пнеймографии. Основная сложность численного выражения изменения минутного объема дыхания (МОД) при такой его регистрации заключается в невозможности в разных опытах (особенно на различных животных) стандартизировать на кимограмме амплитуду волн пнеймограмм, выражающую величину изменения дыхательного объема (ДО). Было предложено определять МОД, ДО и изменения частоты дыхания (ЧД) в процентах к исходному уровню. При этом амплитуду дыхательных движений и частоту дыхания непосредственно до момента вмешательства считать за 100%. В этом случае общеизвестная формула МОД=ДО*ЧД принимает вид:
 , где
, гдеМОД% - изменение интенсивности внешнего дыхания, выраженное в процентах к исходному уровню;
ДО% - изменение дыхательного объема, в процентах к исходному уровню;
ЧД% - изменение частоты дыхания, в процентах.
В литературе не было обнаружено описания подобного метода расчета выраженности изменения внешнего дыхания при его трахеостомической пнеймографии, и на него получено свидетельство о рационализаторском предложении (№ 1138 от 31.03.08). Подробное математическое обоснование данного метода дается в соответствующем разделе диссертации автора. Реакции гемодинамической составляющей кардиореспираторной системы, регистрируемые в наших экспериментах кровавым (прямым) способом, определялись в абсолютных единицах (мм рт. ст.) по выраженности изменения уровня системного артериального давления (р).
Корреляция определялась методом Пирсона. Этот метод позволяет в предположении существования зависимости между величинами x и y выявить истинность, степень и направление такой зависимости. Было показано, что между анализируемыми параметрами существует квадратичная зависимость (y = ax2 + bx + c), характеризуемая в этом конкретном случае достоверностью аппроксимации R2 = 0,56. Тот же массив данных был исследован на существование других зависимостей: линейной, экспоненциальной, степенной, логарифмической. Величины достоверности аппроксимации (R2) в этих случаях составили – 0,38; 0,28; 0,39 и 0,33 соответственно. Следовательно, в ряду выбранных зависимостей наиболее достоверной является именно квадратичная. Значение коэффициента корреляции q при этом составило +0,65, что соответствует прямой средней (ближе к сильной) связи между анализируемыми величинами. Например, большая выраженность рефлекторных ответов дыхательной системы соотносилась с большей выраженностью рефлексов на общее кровяное давление. Тем самым подтверждается существование механизма координации ответных реакций дыхательного и сердечно-сосудистого центров под влиянием афферентации от изученных сосудистых рефлексогенных зон. То есть данные рефлекторные реакции носят системно функциональный характер.
Этот вывод подтверждается и другим методом, основанным на расчете коэффициента Хильдебранта. Как известно, этот коэффициент, характеризующий межсистемные отношения в деятельности гемодинамики и респирации, в норме у человека равен 2,8 – 4,9. В наших экспериментах значение этого показателя в покое, т.е. до активации рецепторов зон позвоночных и сонных артерий, составило 3,25 – 5,6 (δ = 0,14). Очевидно, что подобное отличие указанных величин связано с видовыми особенностями. При реализации рефлексов от изученных СРЗ, несмотря на фактические отклонения частотных характеристик в работе дыхательной системы и сердца, не происходит рассогласования в осуществлении кардиореспираторного взаимодействия. Данный показатель (коэффициент) практически не изменялся, оставаясь равным своей первоначальной (до вмешательства) величине.
Баро- и хеморефлексы, полученные в наших экспериментах с позвоночных и сонных артерий, были качественно однонаправленными (рис. 2), т.е. в деятельности этих двух зон проявляется синергизм. Однако, выраженность последних оказывалась больше, что свидетельствует о меньшем пороге рецепторов каротидного синуса и их большей функциональной активности. Кроме того, в исследованных условиях формируется тип кардиореспираторного ответа с доминированием дыхательной составляющей. Например, средняя выраженность реакций внешнего дыхания при его, например, стимуляции составила 187,42+82,17%, т.е. увеличивалась почти в 3 раза по отношению к исходному (до вмешательства) уровню (рис. 2). При этом повышение САД составляло 14,42+6,29 мм рт. ст., что соответствует менее 10% от исходного уровня систолического давления.
Таким образом, на основании проведенных комплексных физиологических исследований, качественного анализа результатов экспериментов, корреляционного анализа (с использованием различных математических методов) выявлено участие баро- и хеморецепторов СРЗ ПА и каротидного синуса в формировании сопряженных на систему дыхания и собственных на гемодинамику рефлексов, являющихся результатом интегративной регуляции деятельности КРС. Достижение этой функциональной системой общего приспособительного результата (регуляции гомеостаза газообмена и кислотности на уровне тканей) в заданных условиях достигается преимущественно за счет реакций внешнего дыхания и в меньшей степени за счет кардиального или васкулярного компонентов, а также за счет большей реактивности САД в сравнении с работой сердца.
ЗАКЛЮЧЕНИЕ
Настоящей работой показано, что в позвоночных артериях находится самостоятельная рефлексогенная зона, функциональный аналог классической зоны каротидного синуса. Их совместная баро- и хеморецептивная активность направлена на регуляцию деятельности дыхательной и сердечно-сосудистой систем, которые в условиях исходного сдвига рН крови и/или кровяного давления объединяются в единую кардиореспираторную функциональную систему. Мотивирование этой функциональной системы осуществляется афферентацией от исследованных СРЗ, а деятельность проявляется описанными баро- и хеморефлексами. Собственные и сопряженные как хемо-, так и барорефлексы различных СРЗ, в частности ЗПА, направлены на достижение общего конечного полезного результата – коррекции КОС тканей и газообмена в них. Полученные лабораторные данные нашли применение в клинике, предложены новые методики оперативной диагностики развития состояний ацидоза или алкалоза. Разработана система экспресс оценки КОС рожениц и новорожденных. Эта методика не выдвигается на защиту в рамках настоящей диссертации и упоминается здесь для демонстрации практического использования результатов наших исследований системообразующего значения баро- и хемоафферентации от СРЗ. Использование в гинекологической практике этих коэффициентов обусловлено простотой их определения и возможностью оперативно, а главное, объективно оценить состояние новорожденных и матерей. Обоснованным будет предположение, что методика оценки КОС, на которую получено авторское свидетельство (авт. св во 4638208/30-14 от 13.01.89), может быть использована и в других областях практической медицины.