
Isbn 978-5-7262-1377 нейроинформатика 2011
| Вид материала | Документы |
СодержаниеКлючевые слова Методы исследования Частотно-временная структура пик-волнового разряда Структура предшественников судорожной активности Гипотетический нейросетевой механизм инициации |
- Isbn 978-5-7262-1377 нейроинформатика 2011, 107.92kb.
- Isbn 978-5-7262-1377 нейроинформатика 2011, 136.96kb.
- Isbn 978-5-7262-1377 нейроинформатика 2011, 143.59kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 127.94kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 25.66kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 105.62kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 142.85kb.
- Isbn 978-5-7262-1376 нейроинформатика 2011, 103.58kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 79.42kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 136.25kb.
ISBN 978-5-7262-1377-4. НЕЙРОИНФОРМАТИКА – 2011. Часть 3
Е.Ю. Ситникова, А.А. Короновский, А.Е. Храмов
Институт высшей нервной деятельности и нейрофизиологии РАН, Москва
Саратовский госуниверситет им. Н.Г.Чернышевского
jenia-s@mail.ru
Электрографические корреляты
абсанс-эпилепсии и состояния
“судорожной готовности”
(по данным вейвлетного анализа)
Проведено исследование ритмической активности головного мозга при развитии абсанс-эпилепсии. Обнаружено синхронное появление дельта и тета компонентов непосредственно перед началом эпилептических разрядов (т.н. состояние «судорожной готовности»). Предполагается, что в данном состоянии происходит нарушение баланса в работе главных осцилляторных систем головного мозга (кортико-таламо-кортикальной и септо-гиппокампальной нервных сетей), что способствует возникновению пик-волновых разрядов.
^ Ключевые слова: генетическая модель абсанс-эпилепсии, крысы линии WAG/Rij, вейвлетный анализ, дельта активность, тета-компонент
Введение
Электроэнцефалографическое исследование традиционно используется в клинике неврологических заболеваний. Результаты электроэнцефалографии (ЭЭГ) являются главным источником информации для диагностики эпилептических расстройств в условиях стационара или амбулатории. Особого интереса заслуживают новые методы и подходы к анализу ЭЭГ, которые направлены на выявление патологических процессов в центральной нервной системе, дающие прогностическую информацию о течении болезни. В рамках этой проблемы мы провели исследование характера изменений ритмической активности головного мозга при развитии эпилепсии, а именно, абсанс-эпилепсии.
Клинически абсанс-эпилепсия проявляется как кратковременная потеря сознания (от нескольких секунд до нескольких минут), не сопровождающаяся явными расстройствами движений (за исключением подергиваний мышц лица и глаз). В нашей работе были использованы крысы линии WAG/Rij, имеющие генетическую предрасположенность к абсанс-эпилепсии [1]. У человека и исследованных нами животных приступы абсанс-эпилепсии проявляются внезапно при отсутствии внешних признаков, по которым можно было бы предсказать их появление. Это значительно затрудняет контроль состояния больных и отягчает течение болезни.
Диагностическим признаком абсанс-эпилепсии служат генерализованные пик-волновые разряды на ЭЭГ. Считается, что пик-волновые разряды появляются вследствие эпилептической трансформации особой ритмической активности, возникающей в норме во время медленноволнового сна – сонных веретён [2]. Однако, сонные веретена сами по себе не могут провоцировать пик-волновые разряды. Показано, что пик-волновым разрядам предшествуют 5-9 Гц ритмическая активность (исследования на крысах линии GEARS [3]). Интересно, что на ЭЭГ здоровых животных обнаружены 5-9 Гц осцилляции, идентичные предшественникам эпилептических разрядов у крыс линии GEARS. Таким образом, пик-волновые разряды могут развиваться на фоне нормальных физиологических ритмов. Нейрофизиологические механизмы, лежащие в основе инициации эпизодов абсанс-эпилепсии до сих пор остаются неизвестными. Это является центральной проблемой нашего исследования.
^ Методы исследования
I. Экспериментальная работа была выполнена на крысах с генетической предрасположенностью к абсанс-эпилепсии (линия WAG/Rij). Многоканальную регистрацию ЭЭГ у крыс проводили в свободном поведении с использованием вживленных электродов. Запись ЭЭГ осуществляли при стандартном световом режиме непрерывно в течение длительного времени (от 5 до 24 часов). В эксперименте были использованы самцы крыс линии WAG/Rij в возрасте от 5 до 12 месяцев (всего 24 крыс).
II. Распознавание эпизодов эпилептической активности на ЭЭГ проводили в соответствии с рекомендациями Международной Федерации электроэнцефалографии и клинической нейрофизиологии, IFSECN [4]. Пик-волновые разряды состояли из последовательно сменяющих друг друга высокоамплитудного пика и медленной волны. В данной работе мы руководствовались критериями пик-волновых разрядов, разработанных с учетом видовых особенностей крыс WAG/Rij [5, 6]. Амплитуда пик-волновых разрядов более чем в 3 раза превышала амплитуду фоновой ЭЭГ, длительность разрядов составляла не менее 1 секунды. Пик-волновые разряды имели резко очерченную асимметричную форму с преобладанием негативных компонентов: пика и волны (рис. 1).
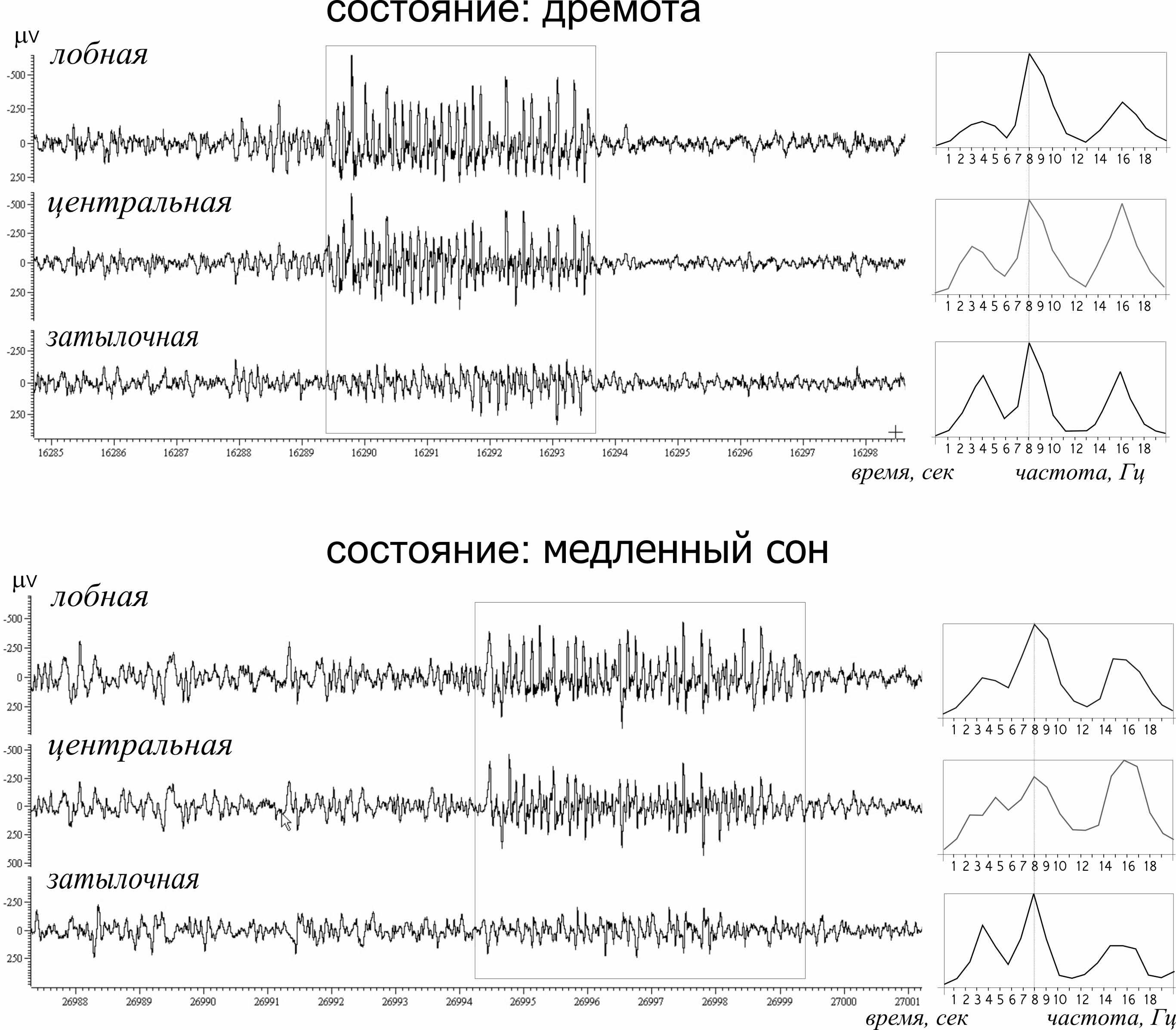
Рис. 1. Пик-волновые разряды на электроэнцефалограмме крысы линии WAG/Rij (самец, возраст 8 месяцев). Регистрирующие электроды находились в трёх областях коры больших полушарий (лобной, центральной и затылочной). Эпилептические разряды помещены в рамку, справа показаны соответствующие спектры мощности (метод Фурье)
III. Частотно-временной анализ ЭЭГ проводили с использованием традиционного для энцефалографии метода быстрого преобразования Фурье, а также непрерывного вейвлетного преобразования с использованием комплексный вейвлет Морле в качестве базисной функции [7]. Непрерывное вейвлетное преобразование представляет собой свёртку исследуемого сигнала x(t) и базисной функции
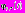 :
: 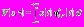 , где * означает комплексное сопряжение. Базисная функция может быть получена из материнского вейвлета с помощью следующего преобразования:
, где * означает комплексное сопряжение. Базисная функция может быть получена из материнского вейвлета с помощью следующего преобразования: 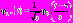 , где s – временной масштаб, определяющий растяжение или сжатие материнской функции, τ – временной сдвиг вейвлетного преобразования и
, где s – временной масштаб, определяющий растяжение или сжатие материнской функции, τ – временной сдвиг вейвлетного преобразования и 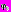 – прототип вейвлетной функции, называемый также материнским вейвлетом.
– прототип вейвлетной функции, называемый также материнским вейвлетом. Результаты предварительных исследований свидетельствуют о том, что комплексный вейвлет Морле является наиболее подходящим материнским вейвлетом для частотно-временного анализа сигналов физиологической природы [7, 8]. Комплексный вейвлет Морле описывается формулой:
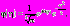 , где параметр
, где параметр 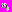 определяет форму и ширину вейвлетной функции в фурье-пространстве, а, следовательно, качество разрешения различных частотных компонент сигнала. Параметр определяет отношение между временными масштабами преобразования s и частотами f исходного сигнала. Выбор
определяет форму и ширину вейвлетной функции в фурье-пространстве, а, следовательно, качество разрешения различных частотных компонент сигнала. Параметр определяет отношение между временными масштабами преобразования s и частотами f исходного сигнала. Выбор 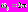 обеспечивает оптимальное частотно-временное разрешение, при этом соотношение между масштабами вейвлетного преобразования и частотами Фурье-спектра сводится к простой формуле s = 1/f.
обеспечивает оптимальное частотно-временное разрешение, при этом соотношение между масштабами вейвлетного преобразования и частотами Фурье-спектра сводится к простой формуле s = 1/f. Вейвлетный спектр – это величина
 представляющая собой мгновенное значение энергии преобразования, т.е. величину энергии, приходящейся в момент времени τ на масштаб s.
представляющая собой мгновенное значение энергии преобразования, т.е. величину энергии, приходящейся в момент времени τ на масштаб s.^ Частотно-временная структура пик-волнового разряда
Спектральный анализ пик-волновых разрядов с использованием быстрого преобразования Фурье показал, что пик мощности разрядов приходится на частоту ~10 Гц. В спектре были найдены дополнительные пики на частоте ~ 20 Гц (рис.1) и иногда ~ 30 Гц, т.е. высокоамплитудные гармоники, кратные основной частоте следования пиков в составе пик-волновых разрядов. Из теории сигналов известно, что высокочастотные гармоники появляются на спектре мощности сигнала, имеющего регулярные резкие скачки амплитуды (на ЭЭГ это – периодически возникающие острые волны или пики). Появление гармоник на спектре пик-волновых разрядов было впервые описано в работе [9] на крысах WAG/Rij, но этот феномен не получил объяснения и ему не придали особого значения.
Непрерывный вейвлетный анализ пик-волновых разрядов, проведенный с помощью комплексного вейвлета Морле, показал, что, во-первых, основная частота следования пиков в течении пик-волнового разряда плавно снижалась от максимального значения в начале разряда 11,1±1,5 Гц (здесь и далее среднее±станд.откл.) до 7,4±0,8 Гц – в конце. Таким образом, в течение пик-волновых разрядов происходило снижение средней частоты в среднем на 2,8±0,8 Гц. Этот феномен был впервые описан И. Мидзяновской и соавторами [10], а затем исследован более детально с применением вейвлетного анализа на базе модифицированного вейвлета Морле [11]. Наши данные полностью подтверждают существование характерной динамики частоты внутри эпилептического разряда, которая была описана в ранних работах [10, 11].
Во-вторых, мы обнаружили, что появление эпилептических (пик-волновых) разрядов на ЭЭГ сопровождалось резким увеличением мощности в широком диапазоне частот 10–100 Гц. При этом каждый отдельный пик, входивший в состав пик-волнового комплекса, отображался на вейвлетном спектре в виде локального всплеска мощности (рис. 2).
Эти высокочастотные спектральные компоненты, порожденные регулярной последовательностью пиков на ЭЭГ, послужили важным диагностическим признаком, который лег в основу алгоритма автоматического распознавания пик-волновых разрядов [8]. Данный метод автоматической диагностики эпилептических разрядов на ЭЭГ был основан на расчете суммарной энергии вейвлетного спектра в диапазоне частот 30–50 Гц. Момент, когда это значение превышало пороговый уровень и не снижалось в течение 1 сек, фиксировали как начало эпилептической активности. Момент, когда уровень энергии оказывался ниже порогового, – как конец разряда. Метод позволил распознавать пик-волновые разряды на ЭЭГ с точностью 98–100 %.
^ Структура предшественников судорожной активности
В этой части работы приведены результаты анализа ЭЭГ, зарегистрированной в лобной коре. Данная область коры оказалась наиболее интересной в плане развития эпилепсии, поскольку пик-волновые разряды в этой области имеют максимальную амплитуду. Предметом исследования послужили 3-секундные интервалы непосредственно до начала пик-волновых разрядов (т.н. «предшественники разрядов»). Последние имели сложную частотно-временную структуру, которую можно охарактеризовать как совокупность нескольких частотных компонент в диапазоне от 2 до 12 Гц.
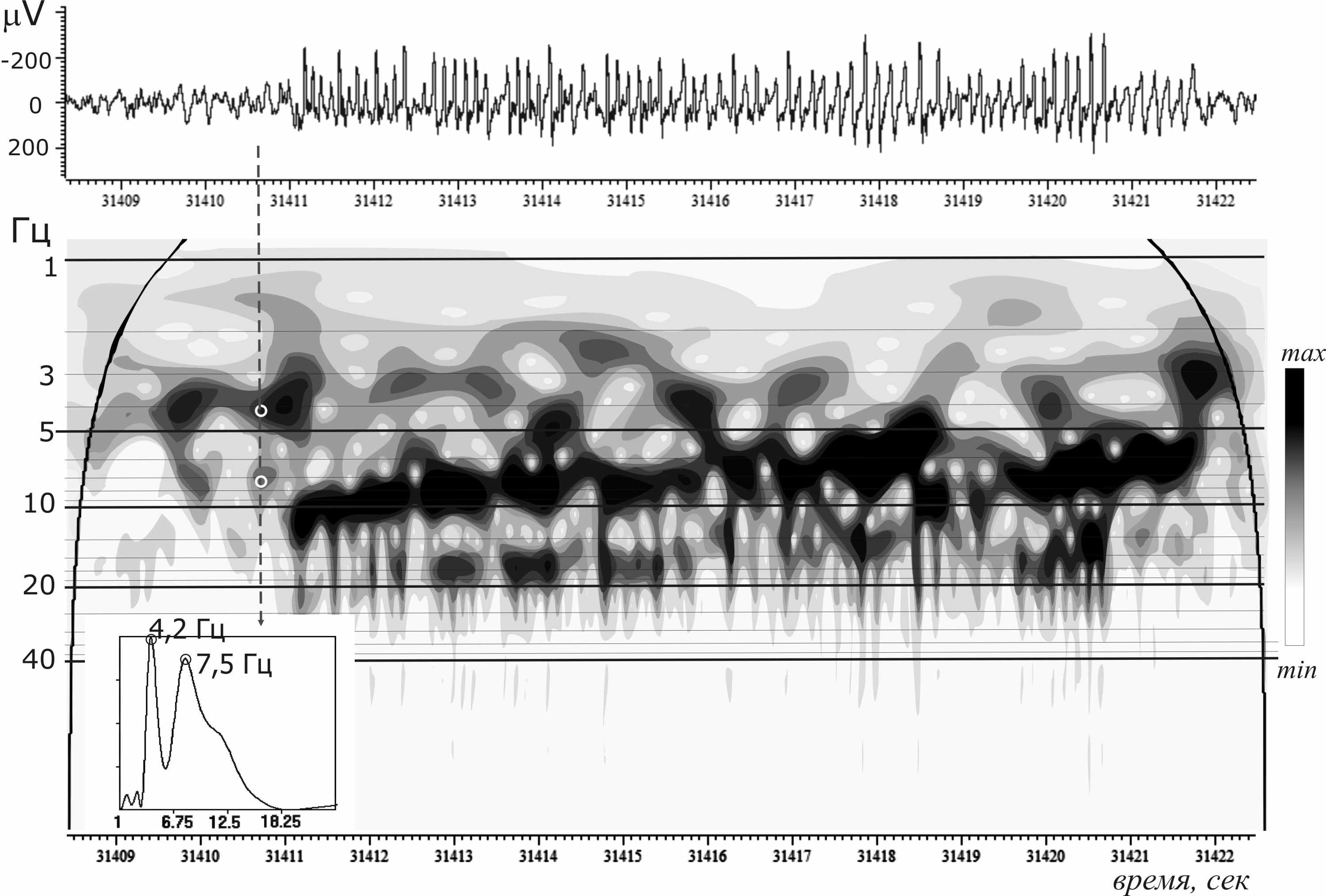
Рис. 2. Частотно-временной портрет пик-волнового разряда (непрерывное вейвлетное преобразование, комплексный вейвлет Морле). На выноске показано распределение мгновенной мощности вейвлетного спектра в диапазоне частот 1–24 Гц за 0,5 сек до начала разряда. На спектре видны медленно-волновые предшественники разряда в диапазоне дельта и тета частот.
Максимальная мощность вейвлетного спектра «предшественников разрядов» была локализована в диапазоне дельта (3–5 Гц) и тета частот (7–11 Гц, pис. 2). Низкочастотные, т.е. дельта компоненты предшествовали ~90 % пик-волновых разрядов, тета компоненты – 92 %. Средняя длительность предшественников составляла примерно полсекунды.
Найдены достоверные отрицательные корреляции между частотой и длительностью дельта-компонентов предшественников (–0,89, p < 0,05), т.е. чем выше частота дельта-компонента, тем он короче. Эту корреляцию можно объяснить существованием некоего гипотетического механизма, обрывающего дельта-предшественники, частота которых оказывалась слишком высокой. У тета-предшественников подобные корреляции отсутствовали.
Лишь у 5 % пик-волновых разрядов ритмические предшественники на ЭЭГ отсутствовали. Эти разряды появлялись на ЭЭГ на фоне десинхронизации. В 89% случаев перед началом пик-волновых разрядов обнаруживали оба компонента (дельта и тета). Такая ситуация была нетипична для фоновой ЭЭГ. Эпизоды времени, на протяжении которых дельта и тета компоненты присутствовали на ЭЭГ одновременно, в состоянии активного бодрствования составляли около 8,6 %. Во время глубокого сна – около 2,2 %, во время поверхностного сна – 9,3 % и во время пассивного бодрствования – 12,9 %. Таким образом, максимальный процент сочетанного появления дельта и тета компонентов на неэпилептической ЭЭГ наблюдался во время пассивного бодрствования и поверхностного сна. Как известно, именно эти состояния служат наиболее благоприятным фоном для возникновения пик-волновых разрядов у крыс WAG/Rij [12]: 33 % от общего числа эпилептических разрядов появляется в состоянии пассивного бодрствования и 48 % – во время неглубокого сна.
^ Гипотетический нейросетевой механизм инициации
абсанс-эпилепсии
Результаты спектрального анализа 1-секундных интервалов ЭЭГ (лобная кора) непосредственно до начала пик-волновых разрядов у крыс WAG/Rij [13] показали, что подавляющее большинство (95 %) этих эпизодов содержали дельта-компонент (1–4 Гц) и 73 % – тета-компонент (4,5–8 Гц). Известно, что дельта активность имеет внутрикортикальное или таламо-кортикальное происхождение [14, 15], в то время как тета активность формируется в септо-гиппокампальной сети. В нервной системе млекопитающих существует механизм, подавляющий осцилляторную активность кортико-таламо-кортикальной сети, когда активна септо-гиппокампальная сеть, и наоборот [14], поэтому в норме дельта и тета осцилляции наблюдаются на ЭЭГ разновременно. Синхронное появление дельта и тета компонентов свидетельствует о нарушении механизма, контролирующего ритмическую активность кортико-таламо-кортикальной и септо-гиппокампальной сетей. Нарушение баланса в работе этих двух осцилляторных систем может способствовать возникновению пик-волновых разрядов.
Сценарий развития пик-волновых разрядов на ЭЭГ, который мы наблюдали у крыс WAG/Rij, отличался от такового у крыс GAERS. Известно, что у крыс GAERS началу эпилептических разрядов предшествует
5–9 Гц ритм, который в течение нескольких секунд приобретает форму пик-волнового разряда [3]. Описанный нами тета-предшественник, средняя частота которого составила 8,6 Гц, в определенном смысле может оказаться аналогом 5–9 Гц предвестника эпилептических разрядов у крыс GAERS. Что касается медленноволнового предшественника абсанс-эпилепсии, то это совершенно новый про-эпилептический элемент на ЭЭГ. О существовании подобного элемента нет упоминаний ни в работах на крысах GAERS, ни на других моделях, ни на человеке. Этот вопрос предстоит исследовать в будущем.
Заключение
Описаны изменения ритмической активности головного мозга, связанные с инициацией пик-волновых разрядов во время абсанс-эпилепсии. Показано, что появлению пик-волновых разрядов предшествует нетипичная (для нормальной неэпилептической ЭЭГ) комбинация ритмических компонентов в диапазоне дельта и тета частот. В целом, наличие одновременно дельта и тета компонентов на ЭЭГ может свидетельствовать о «судорожной готовности» и об опасности возникновения пик-волновых разрядов.
Список литературы
Coenen A.M.L., van Luijtelaar E.L.J.M. Genetic animal models for absence epilepsy: a review of the WAG/Rij strain of rats // Behavior Genetics. 2003. Vol. 33. № 6. P. 635–655.
Kostopoulos GK. Spike-and-wave discharges of absence seizures as a transformation of sleep spindles: the continuing development of a hypothesis // Clinical Neurophysiology. 2000. Suppl. 2: S27–38.
Pinault D., Vergnes M., Marescaux C. Medium-voltage 5–9 Hz oscillations give rise to spike-and-wave discharges in a genetic model of absence epilepsy: in vivo dual extracellular recording of thalamic relay and reticular neurons // Neuroscience. 2001. Vol. 105. P. 181–201.
IFSECN, International Federation of Societies for Electroencephalography and Clinical Neurophysiology. Recommendations for the practice of clinical neurophysiology. Amsterdam, New York: Elsevier, 1983. 191 p.
Sitnikova E., van Luijtelaar G. Electroencephalographic characterization of spike–wave discharges in cortex and thalamus in WAG/Rij rats // Epilepsia. 2007. Vol. 48. P. 2296–2311.
van Luijtelaar E.L.J.M., Coenen A.M.L. Two types of electrocortical paroxysms in an inbred strain of rats // Neuroscince Letters. 1986. Vol. 70. P. 393–397.
Короновский А.А., Храмов А.Е. Непрерывный вейвлетный анализ и его приложения. М.: Физматлит. 2003. 176 с.
Sitnikova E., Hramov A.E., Koronovskii A.A., van Luijtelaar G. Sleep spindles and spike-wave discharges in EEG: their generic features, similarities and distinctions disclosed with Fourier transform and continuous wavelet analysis // Journal of Neuroscience Methods. 2009. Vol. 180. P. 304.
Drinkenburg W.H., van Luijtelaar E.L., van Schaijk W.J., Coenen A.M. Aberrant transients in the EEG of epileptic rats: a spectral analytical approach // Physiology and Behavior. 1993. Vol. 54. P. 779–783.
Midzianovskaia I.S., Kuznetsova G.D., Coenen A.M., Spiridonov A.M., van Luijtelaar E.L. Electrophysiological and pharmacological characteristics of two types of spike-wave discharges in WAG/Rij rats // Brain Research. 2001. Vol. 911. № 1. P. 62–70.
Bosnyakova D., Gabova A., Zharikova A., Gnezditski V., Kuznetsova G., van Luijtelaar G. Some peculiarities of time-frequency dynamics of spike-wave discharges in humans and rats // Clinical Neurophysiology. 2007. Vol. 118. № 8. P. 1736–1743.
Drinkenburg W.H.I.M, Coenen A.M.L., Vossen J.M.H., van Luijtelaar E.L.J.M. Spike-wave discharges and sleep-wake states in rats with absence epilepsy // Epilepsy Research. 1991.Vol. 9. P. 218–224.
Sitnikova E., van Luijtelaar G. Electroencephalographic precursors of spike-wave discharges in a genetic rat model of absence epilepsy: power spectrum and coherence EEG analyses // Epilepsy Research. 2009. Vol. 84. № 2–3. P. 159–171.
Buzsaki G., Draguhn A. Neuronal oscillations in cortical networks // Science. 2004. 304 № 5679. P. 1926–1929.
Steriade M. Neuronal substrates of sleep and epilepsy. Cambridge: Cambridge University Press. 2003.
Работа выполнена при поддержке РФФИ, проект № 09-04-01302-а.
УДК 004.032.26(06) Нейронные сети