
С. П. Сапожников Е. М. Лузикова О. И. Московская Общая и медицинская генетика Учебное пособие
| Вид материала | Учебное пособие |
- Темы: общая генетика, генетика человека, медицинская генетика. Вопросы для самоподготовки., 39.67kb.
- В. И. Александров Учебное пособие. Российская медицинская академия последиплом, 207.44kb.
- Н. А. Общая ботаника с основами геоботаники: Учебное пособие, 520.6kb.
- О. М. Мышалова общая технология мясной отрасли учебное пособие, 1355.07kb.
- Учебное пособие умо м., Медицинская книга 40 2000 Чадаев А. П. 2006, 776.08kb.
- Глинка Николай Леонидович. Общая химия: учебное пособие, 501.78kb.
- Эволюционная, популяционная и общая генетика. Положения, пересекающиеся с валеологией, 625.07kb.
- Учебное пособие Житомир 2001 удк 33: 007. Основы экономической кибернетики. Учебное, 3745.06kb.
- Учебное пособие, 2003 г. Учебное пособие разработано ведущим специалистом учебно-методического, 794.09kb.
- Учебное пособие, 2003 г. Учебное пособие разработано ведущим специалистом учебно-методического, 454.51kb.
ГЛАВА 2. Закономерности наследования признаков. Внутриаллельные отклонения от законов наследования
2.1. Учение о законах наследования различных признаков.
Данное учение начал развивать Г. Мендель. Благодаря его исследованиям выделились три закона наследования, которые носят статистический характер и являются универсальными. Но для проявления этих законов необходимо выполнение следующих условий:
- гены одной или нескольких аллелей должны находиться в одинаковых локусах гомологичных хромосом;
- между генами не должно быть сцепления;
- взаимодействие между генами должно происходить по принципу полного доминирования;
- должна быть равная вероятность образования гамет и зигот разного типа, а также равная вероятность выживания организмов с различными генотипами.
В табл. 2 приведены примеры признаков, наследуемых по доминантному или рецессивному типу.
Таблица 2
Менделирующие фенотипические признаки и болезни человека
| Признак | Тип наследования | |
| Доминантный | Рецессивный | |
| Волосы | Темные | Светлые |
| Не рыжие | Рыжие | |
| Курчавые | Прямые | |
| Поседение волос | В возрасте 25 лет | После 40 лет |
| Облысение | У мужчин | У женщин |
| Глаза | Большие | Маленькие |
| Цвет глаз | Карие, зеленые | Серые, голубые |
| Тип глаз | Монголоидный | Европеоидный |
| Разрез глаз | Прямой | Косой |
| Ресницы | Длинные | Короткие |
| Ямочки на щеках | Есть | Нет |
| Пигментация кожи и глаз | Нормальная | Альбинизм |
| Ушные мочки | Свободные | Приросшие |
| Слух и речь | Нормальные | Глухонемота |
| Губы | Полные | Тонкие |
| Ноздри | Широкие | Узкие |
| Пальцы | Брахидактилия | Нормальная длина |
| Форма носа | «Римский нос» | Прямой |
| Кончик носа смотрит прямо | Курносый | |
| С горбинкой | Прямая переносица | |
| Преобладающая рука | Правая | Левая |
| Веснушки | Есть | Нет |
| Способность свертывать язык трубочкой | Есть | Нет |
| Голос (у женщин) | Сопрано | Альт |
| Голос (у мужчин) | Бас | Тенор |
| Абсолютный музыкальный слух | Имеется | Отсутствует |
| Цвет кожи | Черная | Белая |
| Антигены системы АВО | А, В | О |
| Резус-фактор | Положительный | Отрицательный |
| Строение молекулы гемоглобина | Нормальное | Серповидно-клеточная анемия |
| Наличие фермента | Есть | Алькаптонурия |
| Есть | Фенилкетонурия | |
| Деятельность островковой ткани поджелудочной железы | Нормальное | Нарушение —сахарный диабет |
| Деятельность нервной системы | Мигрень | Нормальное состояние |
2.2. Затруднения в интерпритации наследования признаков при неполном доминировании.
При дальнейшем изучении передачи потомству родительских признаков было выяснено, что имеется множество вариантов, когда законы Г. Менделя не выполняются. Это отклонения наблюдаются как при внутриаллельном, так и при межаллельном взаимодействии генов.
Затруднения в интерпретации наследования признаков возникают: при плейотропии, наличии летальных генов, неполном доминировании, сверхдоминировании, кодоминировании, множественном аллелизме, аллельном исключении и при сцепленном наследовании.
Плейотропия – это явление, когда один ген отвечает за проявление нескольких признаков. Вероятно, это наблюдается в тех случаях, когда наследуется ген, обеспечивающий недостаточность фермента или структурного компонента, играющих важную роль в формировании нескольких типов тканей или в одной, но широко представленной в организме. Примерами плейотропного действия гена являются синдром Марфана и «голубых склер».
Иногда в плейотропном проявлении гена гомозиготы не жизнеспособны (брахидактилия). Известны случаи, когда летальные гены отвечают и за проявление одного признака (серповидно-клеточная анемия - видоизмененный гемоглобин). Неполное доминирование – это явление, когда доминантный ген не полностью подавляет рецессивный и потомки получают лишь часть доминантного признака. Известно и противоположное явление – сверхдоминирование. В этом случае доминантный ген проявляет себя сильнее в гетерозиготном состоянии, нежели в гомозиготном. Вероятно, это явление можно объяснить взаимодействием продуктов генной активности. Повышение жизнеспособности у потомков с увеличением количества гетерозиготных аллелей получило название гетерозиса.
При кодоминировании гены одной аллельной пары равнозначны, и, находясь в генотипе, оба проявляют свое действие. Типичным примером кодоминирования является наследование групп крови по системе АВО и MNS.
Множественные аллели – это явление, когда в популяции более чем два аллеля. Помимо доминантного и рецессивного генов имеются еще и промежуточные, которые подавляются доминантным геном, а для рецессивного являются доминантными.
Аллельное исключение наблюдается у гетерозиготных организмов, когда в одних клетках активна одна аллель, а в других – другая. Это внутриаллельное взаимодействие присутствует у всех женщин, т.к. в одних клетках спирализуется отцовская Х хромосома, а в других – материнская.
Сцепленное наследование генов. Особенности совместного наследования генов одной пары гомологичных хромосом (сцепленных генов) были изучены Т. Морганом и его сотрудниками с помощью дигибридных и тригибридных скрещиваний дрозофилы. Такие гены преимущественно наследуются сцепленно, однако эта тенденция с той или иной вероятностью может нарушаться за счет кроссинговеров, приводящих к генетическому обмену на разных участках гомологичных хромосом. Возможны два типа сцепления генов — полное и неполное (частичное). Полное сцепление было обнаружено в лаборатории Т. Моргана у самцов дрозофил. Выяснили, что в этом случае отсутствовал кроссинговер при конъюгации гомологичных хромосом в профазе первого деления мейоза. Так, у дигетерозиготной особи с условным генотипом АВ//аb происходило образование двух вариантов некроссинговерных гамет (АВ и аb) с равной вероятностью (1/2, или 50 %). Полное сцепление должно наблюдаться также и в отношении неаллельных генов негомологичных участков половых хромосом у организмов гетерогаметного пола (рис.14).
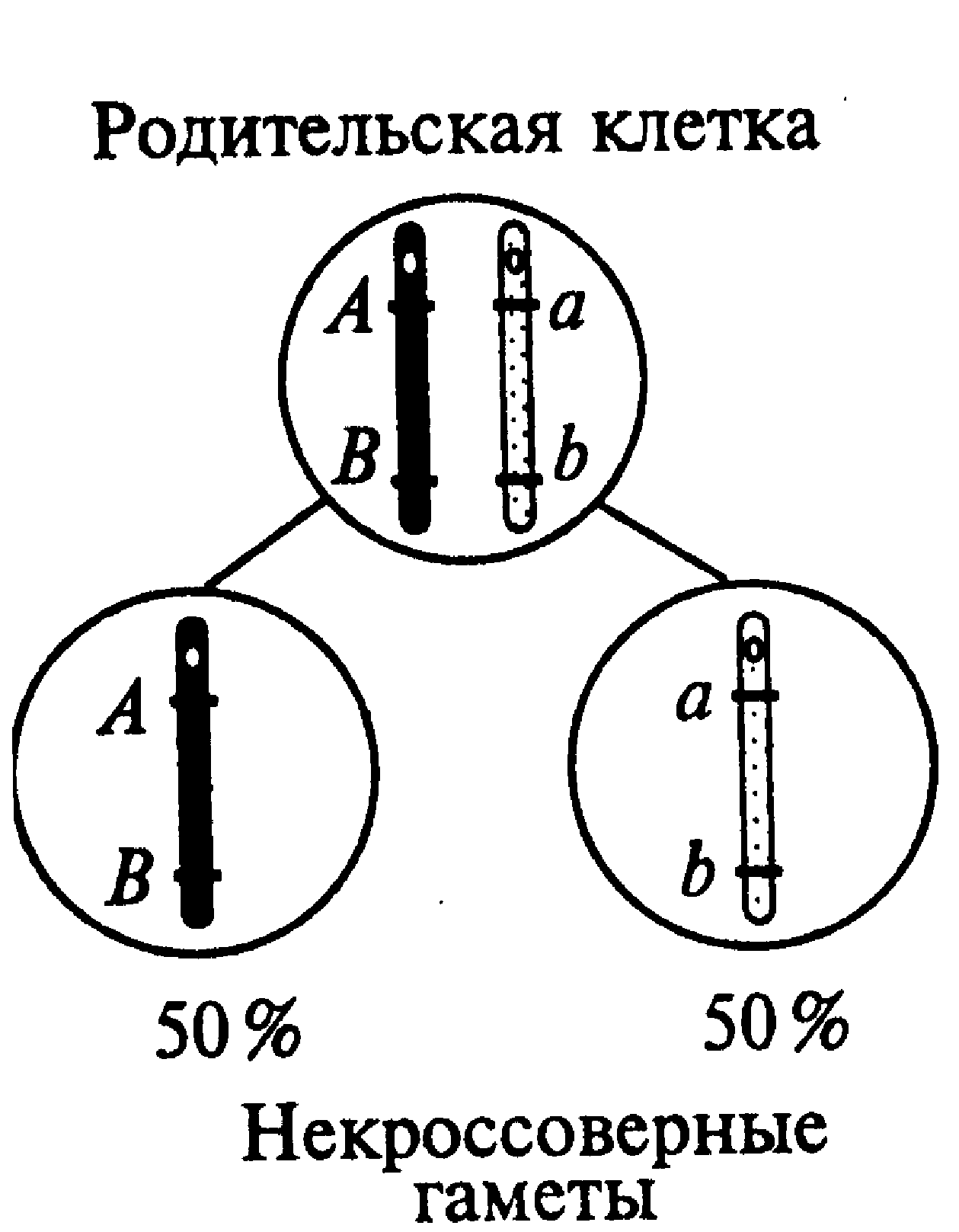 | 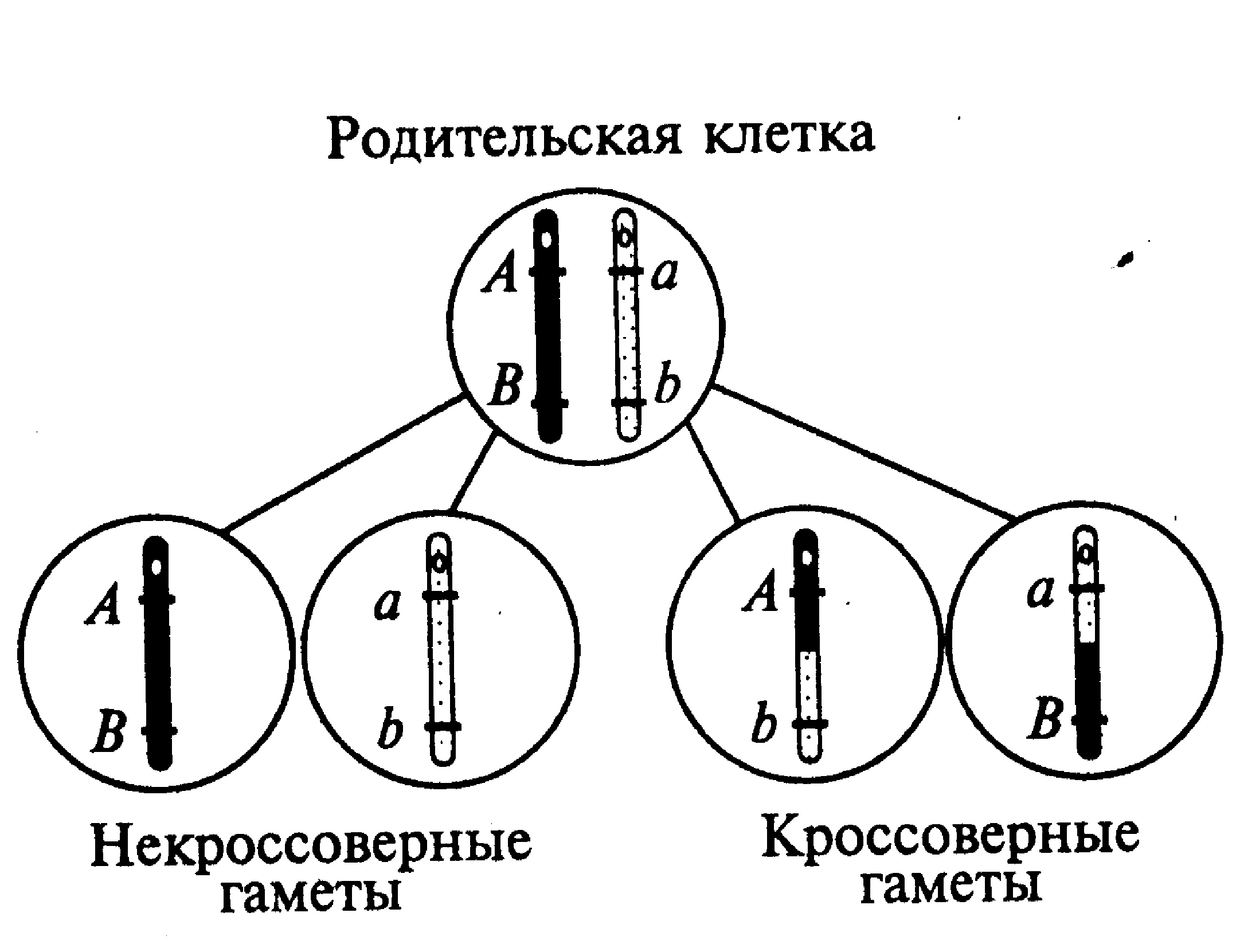 |
| Рис. 14. Образование гамет при полном сцеплении генов | Рис. 15. Образование гамет при неполном сцеплении генов |
Гораздо чаще встречается неполное (частичное) сцепление генов, в основе которого лежит генетическая рекомбинация (кроссинговеры) на тех или иных участках гомологичных хромосом во время мейотического деления клеток. Как видно на рис.15 у индивидуума с тем же условным генотипом АВ//аb формируются четыре варианта гамет — некроссинговерные (АВ, аb) и кроссинговерные гаметы (Аb, аВ). Поскольку кроссинговер представляет собой случайное и относительно редкое событие, то количество образовавшихся с его участием кроссинговерных гамет будет меньшим, чем количество некроссинговерных гамет. В сумме вероятность последних в каждом отдельном случае всегда больше, чем 0,5 (50%).
В соответствии с правилом, установленным Морганом, количество кроссинговеров на участках между локусами сцепленных генов пропорционально расстояниям между этими локусами на хромосомах. Следовательно, чем больше расстояние между двумя конкретными локусами, тем более значительным будет число различных кроссинговеров на участках двух гомологичных хромосом, расположенных между этими локусами, и более значительным количество соответствующих кроссинговерных гамет. Нужно, однако, иметь в виду, что на эффективность самого процесса рекомбинации у разных организмов могут оказывать влияние как их генетические особенности, так и некоторые факторы среды. Поэтому возможны, например, те или иные различия в частоте кроссинговеров на одном и том же хромосомном участке для ивидуумов, относящихся к одному виду, но имеющих разную половую принадлежность. При этом более низкая частота обычно характерна для особей гетерогаметного пола, а у самцов дрозофил в половых хромосомах кроссинговеры вообще не происходят.
Для количественной оценки кроссинговеров между локусами конкретных генов и соответствующих кроссинговерных гамет проводят анализирующие скрещивания особей с определенными различиями по двум либо трем парам сцепленных аллелей. В таких скрещиваниях анализу подвергается генетическая структура одного из родителей, имеющего доминантные гены (признаки), интересующие исследователя, тогда как второй родитель является гомозиготным по рецессивным аллелям изучаемых генов. В качестве примера рассмотрим дигибридное анализирующее скрещивание дрозофил, проведенное Т.Морганом и его сотрудниками с использованием дигетерозиготной самки, в генотипе которой имелись доминантные гены серой окраски тела (условный символ А) и длинных (нормальных) крыльев (символ В), и дигомозиготного самца с рецессивными аллелями черной окраски (а) и коротких («зачаточных») крыльев (b). Это скрещивание можно представить в виде схемы:
| Р |  АВ //аb х АВ //аb х  аb //аb аb //аb |
| G | АВ, аb, Аb, аВ аb |
| F1 | АВ //аb: аb//аb: Аb//аb : аВ//аb |
При исследовании полученного потомства были обнаружены четыре фенотипических варианта со следующими частотами: АВ (серый цвет, длинные крылья) — 41,5%, аb (черный цвет, короткие крылья) — 41,5%, Аb (серый цвет, короткие крылья) — 8,5 % и аВ (черный цвет, длинные крылья) — 8,5%. Таким образом, суммарная частота кроссинговерных фенотипов в потомстве (Ab + аВ) в этом скрещивании составила 17 %. Следовательно, количество соответствующих кроссинговерных генотипов потомства и кроссинговерных гамет самки также можно оценить величиной в 17%. По предложению Т. Моргана при составлении генетических карт дрозофилы расстояния между локусами сцепленных генов стали обозначать в виде количества процентов рекомбинационных событий на участках хромосом между этими локусами. Так, в рассмотренном примере расстояние между локусами генов окраски тела и длины крыльев дрозофилы на хромосомной карте будет определяться величиной в 17% рекомбинации. В дальнейшем 1% рекомбинации стали называть морганидой (сантиморганидой).
Исследование явления сцепления генов позволило Моргану сформулировать главную генетическую теорию — хромосомную теорию наследственности. Ее основные положения выглядят следующим образом:
- Каждый вид живых организмов характеризуется специфическим набором хромосом — кариотипом. Специфичность кариотипа определяется числом и морфологией хромосом.
- Хромосомы являются материальными носителями наследственности, и каждая из них играет определенную роль в развитии особи.
- В хромосоме гены располагаются в линейном порядке. Ген — это участок хромосомы, отвечающий за развитие признака.
- Гены одной хромосомы образуют единую группу сцепления и стремятся наследоваться совместно. Количество групп сцепления равно гаплоидному набору хромосом, так как гомологичные хромосомы представляют одну и ту же группу сцепления.
- Сцепление генов может быть полным (100%-е совместное наследование) или неполным. Неполное сцепление генов является результатом кроссинговера.
- Ч

Рис. 16. Схема одинарного перекреста на
стадии четырех хроматид в пахинеме мейоза
астота кроссинговера зависит от расстояния между генами на хромосоме. Чем дальше лежат гены друг от друга, тем чаще между ними образуется перекрест (рис.16).
Перекрест, происходящий в одном участке хромосомы, называют одинарным перекрестом. Поскольку хромосома представляет собой линейную структуру значительной протяженности, то в ней одновременно могут происходить несколько перекрестов: двойные, тройные и множественные.
Если кроссинговер идет одновременно в двух соседних участках хромосомы, то частота двойных перекрестов оказывается ниже той, которую можно рассчитать на основании частот одинарных перекрестов. Особенно заметное снижение наблюдается при очень близком расположении генов. В этом случае кроссинговер в одном участке механически препятствует кроссинговеру в другом участке. Это явление получило название интерференции. С увеличением расстояния между генами величина интерференции падает. Эффект интерференции измеряется отношением фактической частоты двойных перекрестов к теоретически ожидаемой их частоте в случае их полной независимости друг от друга. Это соотношение называется коинциденцией.
Теоретически частота двойных кроссинговеров согласно закону вероятности равна произведению частот двух одинарных перекрестов. Например, если в хромосоме имеются три гена а, b, с и кроссинговер между а и b идет с частотой 15 %, а между b и c с частотой 9% , то в случае отсутствия интерференции частота двойного кроссинговера равнялась бы 0,15 × 0,009 = 0,0135 = 1,35%. Существование множественных перекрестов приводит к увеличению изменчивости гибридного потомства, так как благодаря им возрастает число генных комбинаций и соответственно число типов гамет у гибридов.
На определении частот одинарных, двойных, тройных и т.д. перекрестов основан принцип построения генетических карт. Генетическая карта — это схема, отражающая порядок расположения генов в хромосоме (рис. 17). За основу расчета расстояния между генами берется процент одинарного кроссинговера между ними. К нему добавляются поправки на величину двойного и более сложных перекрестов, которые уточняют расчет. Если имеется три гена, то порядок их взаиморасположения в хромосоме определяется на основании фенотипа класса двойных кроссоверов. При двойном кроссинговере идет обмен средним геном. Следовательно, признак, по которому двойные кроссоверы отличаются от родительских особей, определяется этим геном. Например, если гомозиготная серая длиннокрылая самка дрозофилы с красными глазами (все признаки дикого типа доминантны) скрещивалась с гомозиготным темным (рецессивная мутация black) самцом с редуцированными крыльями (рецессивная мутация vestigal) и яркими глазами (рецессивная мутация cinnabar) и в первом поколении самыми малочисленными парными классами (т.е. двойными кроссоверами) были серые мухи с яркими глазами и длинными крыльями и черные с красными глазами и редуцированными крыльями, то, следовательно, ген, контролирующий окраску глаз, является средним.
Рак груди (протоков)
Недостаточность енолазы
Нейробластома
?Гемолитическая анемия Rh-ноль
Эритробластоз плода
Эллиптоцитоз-1
Эритрокератодермия вариабельная
Гипофосфатазия младенческая
Фукозидоз
Поздняя подкожная порфирия
Порфирия гепатоэритропоэтическая
Недостаточность галактоэпимеразы
Ceroid li pofuscinos, нейронный-1, детский
Недостаточность C8, тип I и II
Лейкемия/ лимфома, Т - клеточная
Недостаточность ацил-CоА-дегидрогеназы, средняя цепь
Болезнь кленового сиропа, тип 2
Недостаточность миоаденилатдезаминазы
Недостаточность АМФ – дезаминазы эритроцитов
Гиперплазия надпочечников, тип II
Гипотиреоидизм, незобный
Миопатия из-за недостаточности ЦТФазы
Гипераммонемия из-за недостаточности ЦТФазы
Болезнь Гоше, 2 или более типов
Гемолитическая анемия в результате недостаточности ПК
Эллиптоцитоз-2
Пиропойкилоцитоз
Сфероцитоз, рецессивный
Недостаточность антитромбина III
Болезнь Шарко-Мари-Тус, тип Ib
Подверженность в Vivax malaria
Синдром Криглера-Наджара
Немалиновая миопатия
Недостаточность фактора V
Lupus erythematosus, системный
Нейтропения, иммунная
Катаракта зонулярная пылевидная
Хронический гранулематоз в результате недостаточности NCF-2
Гликогеноз VII
Недостаточность фактора XIIIB
Недостаточность CR1
Недостаточность фактора H
Синдром Ашера, тип 2
Синдром Ван дер Вуда
Недостаточность фумаразы
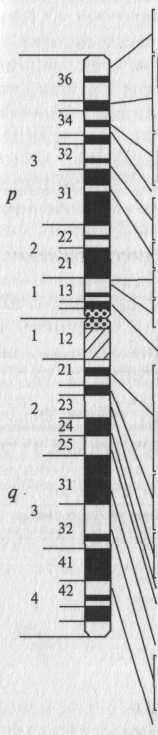
Рис. 17. Генетическая карта Х-хромосомы человека
На генетической карте любой хромосомы отсчет расстояния начинается с нулевой точки – локуса первого гена и отмечается не расстояние между двумя соседними генами, а расстояние в морганидах каждого последующего гена от нулевой точки. Генетические карты составлены только для хорошо изученных в генетическом отношении объектов, как прокариотических, так и эукариотических, таких как, например, фаг λ, Е. соli, дрозофила, мышь, кукуруза, человек. Они являются плодом огромного и систематического труда многих исследователей. Наличие таких карт позволяет предсказывать характер наследования изучаемых признаков, а при селекционной работе — вести сознательный подбор пар для скрещивания.
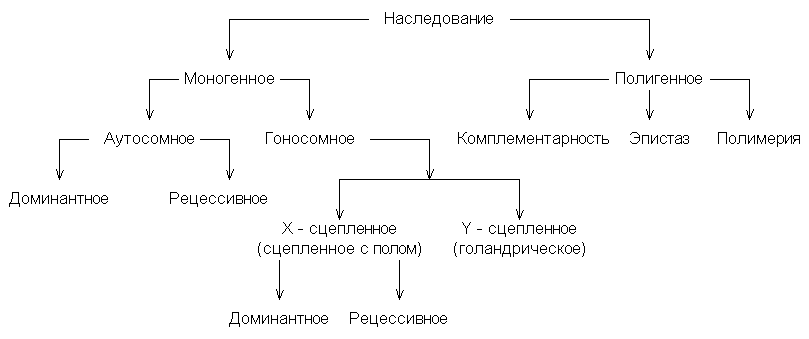
Учитывая количество генов, отвечающих за один признак, а также нахождение гена в соматических или половых хромосомах, все типы наследования можно представить в виде следующей схемы:
Типы наследования признаков

2.3. Группы крови. Резус - система
В настоящее время выявлено несколько антигенных систем, которые определяют групповую принадлежность.
Остановимся на разборе наиболее часто встречающихся. Так, группоспецифические антигены систем АВО, АВН и Lewis находятся на эритроцитах и могут секретироваться в различные жидкости организма. Локусы, контролирующие эти системы, находятся во взаимодействии.
Впервые популяционный полиморфизм по группам крови АВО был замечен в годы Первой мировой войны. По химическому строению группоспецифические вещества относятся к гликопротеинам и гликолипидам. Они (антигены) обнаружены не только на эритроцитах, но и в различных органах, тканях, секретах человека, а также на поверхности лейкоцитов и тромбоцитов. Локализация на генетической карте хромосом – 9g34. Кроме четырех основных генов (А1, А2; В и О) известны их многочисленные варианты различной активности и определенной специфичности, которые связаны с точечной нуклеотидной заменой.
Антигены системы групп крови АВО обладают выраженным плейотропным эффектом в предрасположенности к некоторым заболеваниям. Ряд авторов полагают, что на формирование географического распределения групп крови АВО оказали влияние эпидемии чумы, оспы и холеры. В частности, низкий процент людей с группой крови А в Южной, Восточной и Центральной Азии связывают с эпидемиями оспы. В этой части света преобладает группа крови В, а в Австралии – А.
Группоспецифические антигены (А, В и H) содержатся в секретах (сперма, слюна) не всех людей. Было выяснено, что наследование типов секреции обусловлено двумя аллельными генами (Se и se), локализованными на генетической карте хромосом – 19g13.3. Данный локус тесно сцеплен и взаимодействует с локусами 19g13.2 и 19p13.3 (контролирует миотоническую дистрофию). Биологическая роль групповых антигенов секретов и жидкостей организма человека до конца не ясна, но обнаружено, что среди больных язвенной болезнью желудка и двенадцатиперстной кишки частота невыделителей выше.
В 1946 году Моурант обнаружил, что сыворотка крови женщины по фамилии Lewis агглютинирует примерно 20% случайно отобранных образцов эритроцитов. Антигены системы Lewis были найдены и в слюне. Всего в системе групп крови Lewis известно четыре антигена: a, b, c и d. Их биологическая роль до конца не ясна. Описаны случаи посттрансфузионных осложнений, вызванных антителами Lea и Leb. Локализация на генетической карте хромосом - 19p13.3. Эритроцитарные фенотипы системы Lewis тесно связана с типами секреции АВН - антигенов. Лица с фенотипами Le (a + b -) являются невыделителями, а с Le (a – b +) — выделителями группоспецифических антигенов АВН.
Группы крови P были открыты К. Ландштейнером и П. Левиным в 1927 году, одновременно с системой MN. Биохимически антигены P представляют собой гликосфинголипиды и локализованы на 22-й хромосоме, g - плече, 11.2 локусе.
Локус p1 включает аллели p1, p2, p1k, p2k, p, из которых p1 – доминантный, p2 и p – рецессивные, а p1k и p2k – тормозные. Общее число генотипов равно восьми. Методами серологии определяют четыре фенотипа – p1, p2, pk и Tja. Антигенная несовместимость по системе групп крови Р может вызывать значительные осложнения после переливания крови. Отмечено, что антигены системы Р обладают выраженным плейотропным эффектом в предрасположенности к ряду заболеваний, в частности к остеохондрозу позвоночника.
Один из антигенов этой группы был назван M, другой – N, а через 20 лет обнаружили антиген S и s. MN и Ss – тесно сцепленные, но разделенные локусы находятся на генетической карте хромосом в пределах – 4g28 – g31. Кодоминантные аллели M и N, S и s образуют генотипы MM, MN, NN, SS, Ss и ss и генотипы MS, Ms, NS и Ns. Наследование этих групп крови осуществляется гаплотипами, образованными двумя тесно сцепленными локусами MN и Ss, которые могут образовывать десять генотипов. Фенотипически различимы только 9 из них MS (MSNs), MSs (MSMs), Ms (MsMs), MNS (MSNS), MNSs (MSNs и MsNS), MNs (MsNs), NS (NSNS), NSs (NSNs) и Ns (NsNs).
Установлено, что у больных сахарным диабетом частота группа крови MN достоверно выше, повышается вероятность заболеть острой пневмонией. Высокая частота группы крови M встречается у больных атеросклерозом. Группа крови N чаще встречается у больных остеохондрозом. Имеется также связь MN с артериальным давлением. Среди населения мира аллель M имеет крайне низкую частоту (0,2), но в некоторых районах Австралии и Новой Гвинеи и в ряде регионов Нового Света достигает частоты 0,9. На большей части Европы частота этого аллеля примерно равна 0,5, а частота аллеля S составляет 0,3–0,35 и чаще сочетается с аллелем M. Аллель N имеет еще больший разброс от 1 до 0,006.
В сыворотке крови человека по фамилии Cellano в 1949 году были найдены изоантитела, которые по своей серологической реакции являлись противоположными антителам анти Kell. Описаны многочисленные случаи гемотрансфузионных осложнений и гемолитической болезни новорожденных, возникшие на почве анти Kell и Cellano антигенной несовместимости.
Система Kell является сложной изосерологической системой с большим количеством групповых антигенов (24 – в настоящее время). Гены локализованы на 7-й хромосоме, плече g, в 33-м локусе и сцеплены с локусами, контролирующими ощущение вкуса фенилтиомочевины (7g и5p15).
В Европе группа крови Kell – K встречается с частотой от 5 до 10%, у русских (г. Москва) – 8,02%. Данная система практически отсутствует в Австралии и монголоидных популяциях, редко встречается в Африке и Индии.
Открытие групп крови Rhesus произошло в 1940 году Эритроциты, содержащие антиген, являются резус-положительными и обозначаются Rh (+), а не содержащие – резус-отрицательными Rh (-). Антигены этой системы достаточно выражены на всех этапах онтогенеза, а наименьший плод с Rhesus - антигенами имел возраст 8 недель.
Антигены Rhesus D, C, E локализованы на генетической карте хромосом – 1p36.2 – p34. В этой системе описаны более 46 вариантов антигенов. Частота возникновения гемолитической болезни у новорожденных, обусловленная антигенами системы группы крови Rhesus, убывает в ряду: D – C – c – E – Cw – e. Резус-положительные лица являются либо гомозиготами DD по доминантному аллелю, либо гетерозиготами Dd. Основной полиморфизм в создании гаплотипов в системе Rhesus обеспечивается последовательностью на хромосоме локусов C – E – D. Аллели (C, c, E, e, D и d) двух генных локусов RHD и RHCE обусловливают существование не менее 8 гаплотипов, которые проявляются в 18 фенотипах.
Выявлены закономерности групп крови Rhesus с язвой желудка, остеохондрозом и рядом инфекционных заболеваний. У Rhesus-положительных выше в крови содержание тироксина, а у Rhesus-отрицательных – АКТГ и СТГ.
Несовместимость по резус-фактору возникает в случае, когда резус-положительный плод развивается в матке резус-отрицательной матери. Проникающие в кровь матери резус-антигены плода вызывают синтез резус-антител. Попадание этих антител через плаценту в организм плода сопровождается их взаимодействием с резус-антигеном плода, что приводит к агглютинации эритроцитов и их лизису. В результате разрушения эритроцитов из гемоглобина образуется билирубин, который оказывает токсическое действие на клетки головного мозга плода. Это в свою очередь ведет к нарушению функции центральной нервной системы.
Заместительное переливание крови позволяет спасти многих таких детей, но возникающие нарушения ЦНС часто сказываются на их дальнейшем развитии. В результате резус-конфликта дети рождаются мертвыми либо погибают в течение первых дней жизни. В настоящее время используют метод иммунопрофилактики резус0конфликта у первородящих женщин в послеродовом периоде, связанный с введением им анти-Rh О(D) иммуноглобулина. При повторных беременностях у таких иммунизированных резус-отрицательных женщин не вырабатываются резус-антитела и дети рождаются здоровыми, хотя они и являются резус-положительными.
Тестовые задания
- Что такое гибридизация?
- скрещивание, при котором у родительских особей учитывается одна пара альтернативных признаков;
- скрещивание особей, отличающихся по генотипу;
- скрещивание, при котором у родительских особей учитываются две пары альтернативных признаков;
- скрещивание, при котором у родительских особей учитываются более чем две пары альтернативных признаков.
- Что такое полигибридное скрещивание?
- скрещивание, при котором у родительских особей учитывается одна пара альтернативных признаков;
- скрещивание особей, отличающихся по генотипу;
- скрещивание, при котором у родительских особей учитываются две пары альтернативных признаков;
- скрещивание, при котором у родительских особей учитываются более чем две пары альтернативных признаков.
- Что такое моногибридное скрещивание?
- скрещивание, при котором у родительских особей учитывается одна пара альтернативных признаков;
- скрещивание особей, отличающихся по генотипу;
- скрещивание, при котором у родительских особей учитываются две пары альтернативных признаков;
- скрещивание, при котором у родительских особей учитываются более чем две пары альтернативных признаков.
- Что такое дигибридное скрещивание?
- скрещивание, при котором у родительских особей учитывается одна пара альтернативных признаков;
- скрещивание особей, отличающихся по генотипу;
- скрещивание, при котором у родительских особей учитываются две пары альтернативных признаков;
- скрещивание, при котором у родительских особей учитываются более чем две пары альтернативных признаков.
- При скрещивании гибридов первого поколения (гетерозиготных организмов), анализируемых по одной паре альтернативных признаков, наблюдается расщепление в соотношении 3:1 по фенотипу, что соответствует:
- первому закону Менделя;
- второму закону Менделя;
- третьему закону Менделя.
- Законам Менделя присущи следующие характеристики:
- носят статистический характер;
- проявляются у всех организмов;
- не являются универсальными;
- выполняются на большом количестве особей.
- При скрещивании гибридов первого поколения (гетерозиготных организмов), анализируемых по одной паре альтернативных признаков, наблюдается расщепление в соотношении 1:2:1 по генотипу, что соответствует:
- первому закону Менделя;
- второму закону Менделя;
- третьему закону Менделя.
- Скрещивание гомозиготных особей, анализируемых по одной паре альтернативных признаков, создает единообразие гибридов первого поколения как по фенотипу, так и по генотипу. Это соответствует:
- первому закону Менделя;
- второму закону Менделя;
- третьему закону Менделя.
- При скрещивании гомозиготных организмов, анализируемых по нескольким парам альтернативных признаков, во втором поколении наблюдается независимое комбинирование генов разных аллельных пар и соответствующих признаков, что соответствует:
- первому закону Менделя;
- второму закону Менделя;
- третьему закону Менделя.
- Анализирующее скрещивание — это:
- скрещивание потомков с одним из родителей;
- скрещивание двух родительских особей (АaВb aabb и aabb АaВb);
- исследуемую особь скрещивают с рецессивной гомозиготной;
- исследуемую особь скрещивают с гомозиготной.
- Возвратное скрещивание — это:
- скрещивание потомков с одним из родителей;
- скрещивание двух родительских особей (АaВb aabb и aabb АaВb);
- исследуемую особь скрещивают с рецессивной гомозиготной;
- исследуемую особь скрещивают с гомозиготной.
- Реципрокное скрещивание — это:
- скрещивание потомков с одним из родителей;
- скрещивание двух родительских особей (АaВb aabb и aabb АaВb);
- исследуемую особь скрещивают с рецессивной гомозиготной;
- исследуемую особь скрещивают с гомозиготной.
- Гипотеза «чистоты гамет» сводится к следующим положениям:
- у гибридного организма гены не гибридизируются, а находятся в чистом аллельном состоянии;
- у гибридного организма гены гибридизируются;
- из аллельной пары в гамету попадает только один ген вследствие расхождения гомологичных хромосом и хроматид при мейозе.
- Для проявления законов Менделя необходимо соблюдение следующих условий:
- гены разных аллельных пар должны находиться в разных парах гомологичных хромосом;
- гены разных аллельных пар должны находиться в одной паре гомологичных хромосом;
- между генами не должно быть сцеплений и взаимодействий, кроме полного доминирования;
- должна быть равная вероятность образования гамет и зигот разного типа;
- не должно быть летальных генов.
- Приведите 2 примера проявления летальных генов.
- Приведите 2 примера явления плейотропии.
- Аллельные гены — это:
- гены, располагающиеся в одинаковых локусах гомологичных хромосом;
- гены, располагающиеся в разных локусах гомологичных хромосом;
- гены из одинаковых локусов гомологичных хромосом определяющие развитие взаимоисключающих признаков;
- гены из одинаковых локусов гомологичных хромосом, определяющие развитие одинаковых признаков.
- Отклонение от законов Менделя будет наблюдаться при следующих видах взаимодействия аллельных генов:
- полном доминировании;
- неполном доминировании;
- сверхдоминировании;
- кодоминировании;
- аллельном исключении.
- При каком внутриаллельном взаимодействии в первом поколении наблюдается промежуточное наследование, а во втором – расщепление по фенотипу и генотипу 1:2:1?
- полном доминировании;
- неполном доминировании;
- сверхдоминировании;
- кодоминировании;
- аллельном исключении.
- При каком внутриаллельном взаимодействии гомо- и гетерозиготы фенотипически одинаковы?
- полном доминировании;
- неполном доминировании;
- сверхдоминировании;
- кодоминировании;
- аллельном исключении.
- «Доза гена» - это такая активность гена:
- когда только доминантная гомозигота проявляет признак на 100%;
- когда гетерозигота проявляет признак на 100%;
- когда гетерозигота проявляет признак на 50%.
- При каком внутриаллельном взаимодействии доминантный ген в гетерозиготном состоянии проявляет себя сильнее, чем в гомозиготном?
- полном доминировании;
- неполном доминировании;
- сверхдоминировании;
- кодоминировании;
- аллельном исключении.
- При каком внутриаллельном взаимодействии оба гена при нахождении в генотипе проявляют свое действие?
- полном доминировании;
- неполном доминировании;
- сверхдоминировании;
- кодоминировании;
- аллельном исключении.
- При каком внутриаллельном взаимодействии генов у гетерозигот в разных тканях активны разные аллели?
- полном доминировании;
- неполном доминировании;
- сверхдоминировании;
- кодоминировании;
- аллельном исключении.
- Приведите примеры кодоминирования.
- Множественными аллелями называются аллели, когда в популяции они представлены:
- двумя аллельными состояниями;
- тремя аллельными состояниями;
- одним аллельным состоянием;
- четырьмя аллельными состояниями.
- Приведите примеры аллельного исключения.
- Чистые линии – это организмы, которые:
- являются гомозиготными по изучаемому признаку;
- являются гетерозиготами по изучаемому признаку генотипически;
- при скрещивании получают потомство, не похожее на родителей;
- при скрещивании получают потомство, неотличимое от родителей, но отличное по генотипу;
- при скрещивании дают единообразное потомство как по генотипу, так и по фенотипу.
- Моногенное наследование бывает:
- аутосомное;
- гомосомное;
- полигенное.
- Генотипом называется:
- набор генов организма, развивающихся под воздействием факторов среды;
- совокупность признаков и свойств, развивающихся на базе генотипа под воздействием факторов среды;
- набор генов организма, которые он получает от своих родителей;
- совокупность генов популяции.
- Фенотипом называется:
- набор генов организма, развивающихся под воздействием факторов среды;
- совокупность признаков и свойств, развивающихся на базе генотипа под воздействием факторов среды;
- набор генов организма, которые он получает от своих родителей;
- совокупность генов популяции.
- Генофондом называется:
- набор генов организма, развивающихся под воздействием факторов среды;
- совокупность признаков и свойств, развивающихся на базе генотипа под воздействием факторов среды;
- набор генов организма, которые он получает от своих родителей;
- совокупность генов популяции.