
Федеральная программа книгоиздания России Рецензенты: канд психол наук С. А. Исайчев, доктор биол наук И. И. Полетаева Равич-Щербо И. В. и др. Р12
| Вид материала | Программа |
СодержаниеГенотипическая обусловленность Влияние генотипа на параметры вп Потенциалы мозга, связанные с движением |
- Вестник балтийской педагогической академии вып. 94. – 2010 г. Актуальные проблемы нравственного, 2431.92kb.
- Рецензенты: профессор, доктор психол наук Филонов Л. Б., вед науч сотрудник, канд психол, 2609.63kb.
- Научный выпуск вестник балтийской педагогической академии вып. 29. – 2000 г. Поиск, 1745.18kb.
- Общеобразовательная программа дошкольного образования Авторский коллектив, 5619.19kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Введенским Игорем Витальевичем Рецензенты доктор психол наук В. А. Лабунская канд психол, 375.9kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- Пояснительная записка, 12621.4kb.
- В организации совместных отношений, 1137.14kb.
ГЕНОТИПИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ ОТДЕЛЬНЫХ ПАРАМЕТРОВ И КОМПОНЕНТОВ ВП
Есть основания считать, что индивидуальная изменчивость обшей конфигурации ВП, как и рисунка ЭЭГ в целом, в значительной степени определяется генетическими влияниями. Неоднократно было показано, что изменчивость волновой формы зрительных и слуховых ВП зависит от генотипа, причем наследуемость колеблется в довольно широких пределах, в среднем составляя 0,5 [132, 316]. Латентные периоды отдельных компонентов сенсорных ВП на простые стимулы (вспышки и тоны) также, по-видимому, в значительной степени детерминируются факторами генотипа.
Что можно сказать об источниках изменчивости амплитудных параметров ВП? По одним данным амплитуды контролируются генотипом в меньшей степени, чем латентности [108], по другим, напротив, влияние факторов генотипа более отчетливо выступает именно в амплитудах ВП. Например, в исследовании Дж. Раста [384] оценка наследуемости составляла 80-880% для амплитуд всех компонентов слухового ВП. Для латентностей оценка наследуемости была ниже — 0,35-81%. В то же время, по данным Б.И. Кочубея [84], существуют различия в наследуемости амплитуд ранних и поздних компонентов слуховых ВП: для первых коэффициент генетической детерминации составляет 73-83%, для вторых — 36%. Относительно высокий уровень генетического контроля был обнаружен в показателях динамики амплитудных параметров ВП при изменении интенсивности стимула в процессе изучения феномена «увеличения-уменьшения» [217]. В целом больше данных говорит в пользу генетической обусловленности амплитудных параметров ВП.
324
Таким образом, почти все изученные показатели ВП — волновая форма, отражающая пространственно-временное распределение активных генераторов электрической активности, латентные периоды, характеризующие временной режим распространения возбуждения в ЦНС, и амплитуды, представляющие число активно работающих генераторов и меру их согласованности, — в той или иной степени контролируются генотипом. С позиций информационного подхода эти данные позволяют сделать вывод: межиндивидуальная вариативность скоростных и энергетических аспектов приема и переработки элементарной сенсорной информации зависит от генотипической вариативности.
Однако компоненты ВП и ССП различаются по своему происхождению и функциональному значению. В связи с этим возникает вопрос: какова роль генотипа в изменчивости отдельных компонентов, в первую очередь экзогенных и эндогенных? Считается, что первые определяются внешними факторами — параметрами стимула, тогда как вторые — преимущественно внутренними, такими, как инструкция, мотивация, уровень бодрствования и т.д.
Известно, что сенсорная стимуляция поступает в проекционные зоны коры по коротким путям с минимальным числом, переключений. Однозначно предопределенный характер этих связей говорит в пользу их генотипической обусловленности и дает основания ожидать высокий уровень генетического контроля в параметрах компонентов ВП, отражающих активность этих структур. В то же время передача возбуждения в неспецифических системах мозга может быть высоко вариативной и меняться в зависимости от внесенсорных факторов. В связи с этим есть основания полагать, что роль факторов генотипа в межиндивидуальной изменчивости эндогенных (поздних, неспецифических) компонентов ВП может оказаться ниже, чем в экзогенных (ранних, специфических).
Однако генотип-средовые соотношения в изменчивости отдельных компонентов ВП и их параметров изучены мало, а имеющиеся данные противоречивы. Сравнительный анализ ранних и поздних фрагментов волновой формы сенсорных ВП в одних случаях не выявил существенных различий в их генетической обусловленности, в других, вопреки ожиданиям, был установлен относительно больший генетический вклад в дисперсию поздних компонентов ВП.
При анализе вариативности амплитуд и латентностей ВП отчетливых различий между ранними и поздними компонентами также не выявлено. По одним данным генетические влияния сильнее выражены в поздней части ответа, по другим — равно обнаруживаются в изменчивости всех компонентов ВП, по третьим — генетические влияния преобладают в параметрах! ранних компонентов ВП [84, 132, 244, 316]. Сравнительный анализ наследуемости экзогенных и эндогенных компонентов ВП проводился в исследованиях, выполненных с применением oddball paradigm. Однако ожидаемых различий в фак-
325
торах, формирующих изменчивость ранних экзогенных и поздних эндогенных компонентов слуховых ВП, обнаружено не было.
Таким образом, несмотря на теоретически прогнозируемые различия в наследуемости экзогенных и эндогенных компонентов ВП, экспериментальные данные говорят о том, что генотип-средовые соотношения в вариативности амплитуд но-временных параметров экзогенных (ранних специфических) и эндогенных (поздних, неспецифических) компонентов ВП и ССП приблизительно одинаковы.
ВЛИЯНИЕ ГЕНОТИПА НА ПАРАМЕТРЫ ВП
В РАЗНЫХ ЗОНАХ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
При любых условиях ВП и ССП, зарегистрированные в разных зонах коры больших полушарий, имеют свои особенности. Например, в проекционных зонах лучше выражены ранние, экзогенные компоненты, в ассоциативных, напротив, преобладают поздние, эндогенные компоненты. Эта особенность делает ВП и ССП незаменимым инструментом при оценке генетического вклада в изменчивость функциональной активности отдельных зон коры мозга.
Однако данных о региональных особенностях генетической детерминации ВП пока очень мало. В исследовании Э. Льюиса с соавторами генетическая детерминация зрительных, слуховых и соматосенсорных ВП обнаруживалась приблизительно одинаково по всей коре больших полушарий, независимо от зоны [316]. Но наряду с этим, как и при изучении наследуемости параметров ЭЭГ, получены факты, свидетельствующие о существенных межзональных различиях в проявлении генетической обусловленности отдельных компонентов ВП и их параметров. Например, выявлены межзональные различия в генетической обусловленности зрительных ВП на вспышки, регистрируемые из трех зон. Установлено, что генетический контроль обнаруживается в 43, 52 и 20% показателей при регистрации ответов из трех зон: соответственно затылочной, вертекса и лобной области левого полушария [132, гл. IV]. В другом исследовании были установлены межпо-лушарные различия в наследуемости параметров зрительных ВП на стимулы разного типа: в целом параметры ВП в левом полушарии меньше зависят от генотипа по сравнению с аналогичными ответами правого, причем наиболее четко это обнаруживается при сравнении ответов височных зон левого и правого полушарий [132, гл. IV]. Убедительные доказательства топографических различий наследуемости амплитудно-временных параметров ВП представлены в работе О'Коннера с соавторами (см. рис. 14.5), причем показано, что «фокус наследуемости» амплитуды Р3 локализован в задних отделах коры больших полушарий.
Чем можно объяснить тот факт, что дисперсия фенотипически сходных показателей ВП в разных зонах коры мозга имеет разную структуру? Из нейрофизиологических исследований известно, что
326
компоненты ВП в разных зонах коры больших полушарий могут иметь различающиеся источники. Иными словами, они, во-первых, могут формироваться за счет действия различных генераторов и, во-вторых, испытывать на себе разные влияния со стороны неспецифических систем мозга. Можно предположить, что нейрофизиологические механизмы, отвечающие за компоненты ВП разных зон, по-разному зависят от генотипа, и благодаря этому возникают межзональные различия в генетической обусловленности ВП.
Есть некоторые основания полагать, что доля генетической изменчивости в параметрах ВП разных зон варьирует в определенной зависимости от эволюционного возраста структуры и времени ее созревания в онтогенезе. Так, ВП в эволюционно более молодых передних отделах коры (височные и фронтальные области), по-видимому, в меньшей степени зависят от генетической изменчивости. Однако эти предположения требуют дальнейшего изучения.
ПОТЕНЦИАЛЫ МОЗГА, СВЯЗАННЫЕ С ДВИЖЕНИЕМ
Среди событийно-связанных потенциалов особое место занимают потенциалы мозга, связанные с движением (ПМСД). Изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и хронометрировать эти процессы, т.е. установить временные границы их протекания [104, 237].
Впервые комплекс колебаний, отражающий процессы подготовки, выполнения и оценки движения, был зарегистрирован в 60-х годах. Оказалось, что движению предшествует медленное отрицательное колебание — потенциал готовности (ПГ). Он начинает развиваться за 1,5-0,5 мс до начала движения. Этот компонент регистрируется преимущественно в центральных и лобно-центральных отведениях обоих полушарий. Принято считать, что ПГ возникает в моторной коре и связан с процессами планирования и подготовки движения. Он относится к классу медленных негативных колебаний потенциала мозга, возникновение которых объясняют активацией нейрональных элементов соответствующих участков коры.
За 500-300 мс до начала движения ПГ становится асимметричным — его максимальная амплитуда наблюдается в прецентральной области, контралатеральной движению. Примерно у половины взрослых испытуемых на фоне этого медленного отрицательного колебания незадолго до начала движения регистрируется небольшой по амплитуде положительный компонент — Р1. Следующее по порядку быстро нарастающее по амплитуде отрицательное колебание N2, так называемый моторный потенциал (МП), начинает развиваться за 150 мс до начала движения и достигает максимальной амплитуды над областью, в которой находятся корковые центры управления движу-
327

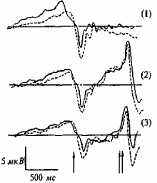
Рис. 14.6. Схема потенциалов, связанных с реализацией двигательного действия.
Одной вертикальной стрелкой обозначено начало движения, двумя — сигнал обратной связи. Компонент Рь обозначен пунктирной линией, поскольку выделяется не у всех испытуемых. Одна стрелка — начало движения, две — сигнал обратной связи [104]. ПГ — потенциал готовности; УНВ — условная негативная волна.
щейся конечностью. Примерно через 200 мс после начала движения возникает положительный компонент Р2. Завершается этот комплекс потенциалов появлением компонентов N3 иР3 (рис. 14.6).
Еще один электрофизиологический феномен по своей сути близок потенциалу готовности. Речь идет об отрицательном колебании потенциала, регистрируемого в передних отделах коры мозга в период между действием предупреждающего и пускового (требующего реакции) сигналов. Это колебание имеет ряд названий: Е-волна, волна ожидания, условная негативная волна (УНВ). Е-волна возникает после предупредительного сигнала, ее длительность растет с увеличением интервала между первым и вторым стимулами. Амплитуда Е-волны возрастает прямо пропорционально скорости двигательной реакции на пусковой стимул. Она увеличивается при напряжении внимания и повышении волевого усилия, что свидетельствует о связи этого электрофизиологического явления с механизмами произвольной регуляции двигательной активности и поведения в целом.
Обнаруживая индивидуальную специфичность и стабильность, эти потенциалы дают возможность оценить вклад генетических факторов в изменчивость биоэлектрических коррелятов индивидуальных особенностей организации и построения движений.
Генетическое исследование ПМСД было проведено С.Б. Малыхом [104] на близнецах 18-30 лет (по 25 пар МЗ и ДЗ близнецов). Особенность этой работы — изучение генетической обусловленности ПМСД в трех психологически различающихся ситуациях: в первой требовалось произвольно (без предварительного сигнала) нажимать на кнопку; во второй и третьей надо было прогнозировать появление звуковых стимулов, организованных в ряд с фиксированной последовательностью и разной вероятностью. Испытуемому сообщалось, что звуки в правом и левом наушниках организованы в блоки, а именно:
328
звук слева может встречаться по 2 или 3 раза подряд, а справа — по 2 или 4 раза подряд. Надо было предсказать появление звука в правом или левом наушнике, сообщая о характере прогноза нажатием на одну из двух кнопок, расположенных справа. Описанная организация стимульного ряда позволяла получать прогноз событий, наступающих с вероятностьюр = 1,0 ир = 0,5.
ПМСД получали усреднением ЭЭГ, зарегистрированной монопо-лярно в зонах F3, F4, C3, С4. Для каждого из четырех отведений у одного испытуемого было получено по три потенциала: при простых произвольных движениях без внешнего сигнала (1), при нажатии кнопки в
ситуации прогноза двух равноверо- Рис. 14.7. ПМСД, УНВ и Р300, ятных сигналов (2) и в ситуации зарегистрированные в отведении заведомо истинного прогноза (3). С3 у пары МЗ близнецов в трех эк-
В ситуации (1) анализировались спериментальных ситуациях [104]. потенциал готовности (ПГ), ком- Одной стрелкой отмечено начало дви-поненты Рь N2, Р2, N3, Р3 отра- жения, двумя — сигнал обратной жавшие соответственно процессы связи. подготовки, реализации и оценки
произведенного движения, В ситуациях (2) и (3) помимо указанных компонентов ПМСД регистрировалась условная негативная волна (УНВ), появлявшаяся между нажатием кнопки и ожидаемым сигналом обратной связи (т.е. звуком, который подтверждал или не подтверждал прогноз), сменявшаяся после предъявления звука положительным компонентом Р300 (рис. 14.7). Два последних компонента отражают процессы ожидания и оценки прогноза. Генетический анализ включал оценку внутрипарного сходства амплитуд и латентных периодов всех выделенных компонентов с помощью внутриклассовой корреляции.
Анализ влияний генотипа и среды на изменчивость потенциалов мозга, связанных с реализацией двигательного действия, дал достаточно пеструю картину (табл. 14,4). Для того чтобы выделить наиболее общие тенденции, по каждому признаку был подсчитан процент тех позиций, в которых обнаруживался генетический контроль. Оказалось, что в амплитудных параметрах генетический контроль обнаруживается чаще (63,2% всех случаев), чем во временных (21%). Правда, латентности компонентов ПМСД измеряются менее надежно,
329
чем в сенсорных ВП, из-за отсутствия внешнего сигнала, от которого обычно ведется счет времени. Возможно, это и сказалось на оценке наследуемости.
Выявились также определенные межзональные различия: менее всего генетический контроль выступает в показателях ПМСД левой лобной области (26,3%), в остальных трех зонах он выше (от 44,7 до 50%).
Наиболее важным представляется тот факт, что вклад генетической и средовой составляющих существенно меняется в зависимости от характера деятельности испытуемого. Чаще всего влияние генотипа обнаруживается в ситуации прогноза равновероятных событий (53,6% всех исследованных признаков), реже всего (20%) — при простых произвольных движениях, и промежуточное положение (46,4%) занимает ситуация с заведомо истинным прогнозом. Таким образом, при осуществлении прогнозирования генетический контроль обнаруживается чаще, чем при простых произвольных движениях, не являющихся ответом на внешний стимул и осуществляемых по субъективной команде (см. табл. 14.4).
Закономерно возникает вопрос: почему ПМСД, сопровождающие по фенотипическому проявлению одно и то же движение, обнаруживают столь разную степень генетической обусловленности? Объяснение этого факта видится в разном функциональном значении данного движения в изучаемых ситуациях. При простом произвольном нажатии кнопки испытуемый сам инициирует движение. Иначе говоря, в соответствии с инструкцией цель действия — само нажатие на кнопку. Во второй ситуации целью является прогноз, а движение становится лишь средством достижения этой цели. Поскольку двигательные акты в первом и втором случаях различаются, прежде всего по степени произвольности и осознаваемости и, следовательно, по уровню нейрофизиологического обеспечения, постольку изменение роли генотипа в вариативности соответствующих ПМСД можно объяснить, по-видимому, отражением в их параметрах активности разных функциональных систем.
Конфигурация ВП, а также их основные количественные параметры относятся к числу индивидуально устойчивых особенностей человека, что дает основания изучать роль генотипа и среды в происхождении межиндивидуальной вариативности по этим признакам.
В целом имеющиеся в литературе данные свидетельствуют о существенном влиянии генотипа на межиндивидуальную вариативность вызванных и событийно-связанных потенциалов. Генетическую обусловленность обнаруживают сенсорные (зрительные и слуховые) ВП, а также потенциалы мозга, связанные с движением.
330
Таблица 14.4
Наличие генетической составляющей в вариативности амплитудно-временных характеристик компонентов
потенциала в трех экспериментальных ситуациях [104]
| | Амплитуды | Временные | i характеристики | | | | | |||||||||||||||||
| Комп | 1 | | 2 | | | | 1 | | 2 | | | 3 | | | ||||||||||
| о- | | | | | | | | | | | | | | | ||||||||||
| нент | F3 | | С3 | C4 | F3 | | С3 | С4 | F3 | | С3 | C4 | F3 | | C3 | С4 | F3 | | C3 | С4 | F3 | | С3 | C4 |
| ы | | | | | | | | | | | | | | | | | | | | | | | | |
| ПГ | | | + | | | + | + | | | + | + | + | | | | + | | | + | | | | | + |
| N2 | | | | | + | + | + | + | | + | + | + | | | | | + | | | | | | | |
| P2 | | + | | | + | + | + | + | | + | + | + | | | | + | + | | + | + | + | + | + | + |
| N3 | | + | | | + | + | + | | | + | + | + | | | | | | | | | | | | |
| Рз | | + | + | + | | + | + | | | + | + | + | | | | | | | | | | | | |
| УНВ | — | — | — | — | + | + | + | + | | | + | | — | — | — | — | | | | + | + | + | | |
| P300 | — | — | — | — | + | + | + | + | + | + | | + | — | — | — | — | | | + | | | | | |