
Федеральная программа книгоиздания России Рецензенты: канд психол наук С. А. Исайчев, доктор биол наук И. И. Полетаева Равич-Щербо И. В. и др. Р12
| Вид материала | Программа |
СодержаниеВлияние генотипа на формирование параметров энцефалограммы Межзональныеразличиявнаследуемостипараметров альфа-ритма ээг |
- Вестник балтийской педагогической академии вып. 94. – 2010 г. Актуальные проблемы нравственного, 2431.92kb.
- Рецензенты: профессор, доктор психол наук Филонов Л. Б., вед науч сотрудник, канд психол, 2609.63kb.
- Научный выпуск вестник балтийской педагогической академии вып. 29. – 2000 г. Поиск, 1745.18kb.
- Общеобразовательная программа дошкольного образования Авторский коллектив, 5619.19kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Введенским Игорем Витальевичем Рецензенты доктор психол наук В. А. Лабунская канд психол, 375.9kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- Пояснительная записка, 12621.4kb.
- В организации совместных отношений, 1137.14kb.
Объектом изучения в этом случае являются количественные непрерывно распределенные в популяции показатели ЭЭГ. Признаками, подлежащими изучению, служат оценки частот, индексов, амплитуд, суммарных энергий и других показателей того или иного ритма ЭЭГ, полученные в группах близнецов, семьях и т.д.
В первых генетических исследованиях, которые проводились на основе визуального анализа энцефалограмм, объектом анализа были в основном параметры альфа-ритма: альфа-индекс, амплитуда, частота. Благодаря использованию автоматического частотного анализа, разлагающего ЭЭГ на частотные диапазоны, появилась возможность исследовать и другие ритмические составляющие в полосе дельта-, тэта- и бета-ритмов. Кроме того, с помощью автоматического анализа стало возможным отдельно оценивать суммарную энергию каждого ритма, а спектральный анализ позволил установить еще одну энергетическую характеристику: доли спектральной мощности, приходящиеся на каждый частотный диапазон. Суммарные энергии и спектральные плотности отражают представленность каждого ритма в общем паттерне ЭЭГ.
В табл. 13.2 из работы ТА Мешковой [132, гл. III] объединены данные ряда исследований, выполненных сходными методами. Усредненные коэффициенты внутриклассовой корреляции характеризуют сходство МЗ и ДЗ близнецов по таким параметрам, как суммарные энергии, спектральные плотности и частоты основных ритмов ЭЭГ. Они свидетельствуют о большем сходстве МЗ близнецов по суммарным энергиям всех ритмов, за исключением медленной ритмики (дельта и тэта). Наибольшее сходство МЗ близнецов характерно для альфа-полосы. Примерно на том же уровне — корреляции по бета-ритму. Очевидно, доля генетической составляющей в популяционной дисперсии этих характеристик альфа- и бета-ритмов довольно велика. Частоты указанных ритмов, за исключением альфа, анализируются в
294
Таблица 13.
Коэффициенты внутриклассовой корреляции МЗ и ДЗ близнецов по суммарным энергиям и спектральным
плотностям отдельных ритмических составляющих (по данным разных авторов) [132, гл. III]
2
| Автор | Год публи- | Число | пар | Диапазоны ЭЭГ | | | | | | | |
| дельта | тэта | альфа | бета | ||||||||
| работы | МЗ | ДЗ | | Гдз | Гмз | Гдз | гмз | rДЗ | Гмз | Гдз | |
| | | | Сумма | рные | энерги и | | | | | | |
| М. Камитаке (М. Kamitake) Дж. Юнг и др. (J. Young et al. ) НФ Шляхта Н Ф. Шляхта, Т А. Пантелеева Г А. Шибаровская НФ Шляхта Т А. Мешкова | 1963 1972 1972 1972 1978 1981 1978 | 26 17 15 19 30 26 20 | 19 15 13 19 26 22 20 | 0,60 0,24 0,61 0,74 0,66 0,58 | 0,11 0,31 0,58 0,66 0,57 0,29 | 0,8 0,66 0,72 0,85 0,78 0,85 | 0,52 0,34 0,95 0,66 0,54 0,41 | 0,96 0,52 0,69 0,95 0,81 0,90 0,93 | 0,15 0,29 0,94 0,43 0,53 0,66 0,34 | 0,84 0,90 0,87 0,77 0,82 0,81 0,79 | 0,17 0,56 0,8 0,24 0,56 0,39 0,49 |
| Средние | 0,59 | 0,44 | 0,79 | 0,65 | 0,87 | 0,55 | 0,83 | 0,49 | |||
| Спектральные плотности | |||||||||||
| Д. Ликкен и др. (D. Lykken et Д. Ликкен и др. (D. Lykken ct al. | 1974 1982 | 39 114 | 27 53 | 0,76 0,88 | -0,01 0,26 | 0,86 0,79 | -0,03 0,04 | 0,82 0,90 | -0,20 0,13 | 0,82 0,67 | 0,15 0,37 |
| Частота ритмов | |||||||||||
| Н.Ф. Шляхта, Т.А. Г А. Шибаровская НФ Шляхта Т А. Мешкова | 1978 1978 1981 1978 | 19 30 26 20 | 19 26 22 20 | 0,11 0,65 -0,14 | -0,13 0,07 -0,02 | 0,18 0,85 0,52 | -0,10 0,82 0,02 | 0,75 0,87 0,43 0,89 | -0,62 0,48 0,36 0,59 | 0,39 0,39 0,47 0,40 | 0,07 0,46 0,29 0,49 |
295
очень немногих работах. Можно видеть, что по частотам всех ритмов, кроме альфа, в основном нет существенной разницы в уровнях внут-рипарного сходства МЗ и ДЗ близнецов, что заставляет предположить наличие средовых влияний в межиндивидуальной вариативности данных признаков.
Высокая наследственная обусловленность параметров альфа-ритма получила подтверждение и в более поздних исследованиях. Так, при оценке альфа-индекса и альфа-частоты в группах, состоящих из 42 пар МЗ близнецов, выросших вместе, и 35 пар МЗ близнецов разлученных [208], не было обнаружено практически никаких различий в степени внутрипарного сходства (коэффициент внутриклассовой корреляции для обеих групп в среднем составлял 0,8).
В совокупности данные подавляющего большинства работ показывают, что независимо от области отведения, способа регистрации и анализа ЭЭГ, возрастного состава и количества испытуемых наиболее значительные наследственные влияния обнаруживаются в диапазоне альфа-ритма. Практически ни в одной работе не отмечается несходства МЗ близнецов по альфа-параметрам.
Вероятно, значительной наследственной обусловленностью именно альфа-характеристик можно объяснить и значительное сходство общего рисунка ЭЭГ МЗ близнецов, поскольку именно альфа-ритм является доминирующим в ЭЭГ покоя.
Влияние генотипа на параметры отдельных ритмов ЭЭГ изучалось также в семейном исследовании, проведенном в сельской популяции туркмен [6]. В ходе исследования изучалась природа популяционной дисперсии абсолютной и относительной мощности основных ритмов ЭЭГ (тэта, альфа, бета-1, бета-2) в лобной, височной и затылочной зонах обоих полушарий. Структура фенотипической дисперсии по каждому показателю анализировалась с помощью генетико-статистичес-ких методов. Для большинства показателей абсолютной мощности вклад генетических факторов оказался достаточно высоким. Аддитивная генетическая составляющая дисперсии варьировала от 18 до 50% для тэта-ритма, от 24 до 68% для альфа-ритма и от 30 до 80% для бета-1-ритма (в зависимости от зоны регистрации). Анализ наследуемости относительной мощности дал более противоречивую картину, тем не менее и в этом случае ряд показателей обнаружил сравнительно высокий уровень наследуемости. В их числе находятся в первую очередь относительные мощности всех анализируемых ритмов ЭЭГ затылочной и височной областей. Аддитивная составляющая дисперсии в подавляющем большинстве случаев превышает 50%.
В общем, результаты близнецовых и семейных исследований так или иначе свидетельствуют о вкладе генотипа в межиндивидуальную изменчивость разных параметров практически всех ритмических составляющих ЭЭГ. Однако наибольшая определенность существует в отношении альфа-ритма. В отношении остальных ритмических состав-
296
ляюших ЭЭГ, во-первых, имеется меньше данных, во-вторых, эти данные, особенно касающиеся медленных составляющих, более разноречивы, и потому пока трудно сделать окончательный вывод относительно любого из ритмов, кроме альфа.
В исследовании А.П. Анохина было также установлено, что у родителей с высокими значениями такой характеристики альфа-ритма, как альфа-индекс, дети в большинстве своем имеют значения альфа-индекса выше среднего, и напротив, у родителей с низким значением альфа-индекса дети чаще всего имеют сравнительно низкий альфа-индекс.
МЕЖЗОНАЛЬНЫЕРАЗЛИЧИЯВНАСЛЕДУЕМОСТИПАРАМЕТРОВ АЛЬФА-РИТМА ЭЭГ
Несмотря на то что в генетических исследованиях ЭЭГ нередко регистрировалась в разных областях, специальное сопоставление отдельных отведений по их отношению к генотипу практически не проводилось. Между тем характер наследственных влияний на биоэлектрическую активность отдельных областей коры, в том числе альфа-ритма, может существенно различаться. Об этом свидетельствует исследование Т.А. Мешковой [132, гл. III], в котором сопоставлялась генетическая обусловленность параметров ЭЭГ из десяти зон: F3, F4, С3, С4, Р3, Р4, Т3, Т4, О1, О2 (зарегистрированная монополярно по системе «10—20»). В экспериментах участвовали 20 пар МЗ и 20 однополых пар ДЗ близнецов 18-26 лет. Кроме того, случайным объединением членов дизиготных пар была составлена контрольная группа однополых пар неродственников (HP). Определялись частота, амплитуда и индекс альфа-ритма во всех перечисленных отведениях.
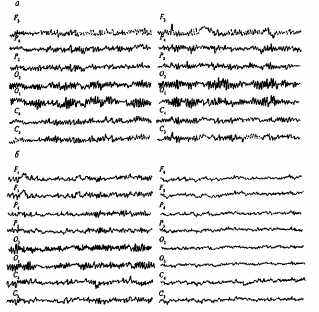
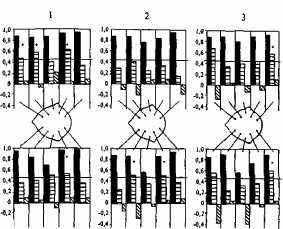
Визуальный анализ ЭЭГ свидетельствует о высоком сходстве паттернов ЭЭГ у МЗ и преимущественно несходстве таковых у ДЗ близнецов (рис. 13.2). На рис. 13.3 графически представлены коэффициенты внутриклассовой корреляции, характеризующие внутрипарное сходство МЗ и ДЗ близнецов по анализируемым параметрам альфа-ритма. Из диаграммы видно, что количественные параметры альфа-ритма очень сходны у МЗ близнецов (соответствующие коэффициенты корреляции высоки и значимы); у ДЗ коэффициенты сходства в большинстве случаев не достигают уровня значимости. Если же коэффициенты сходства ДЗ близнецов статистически значимы, то различия между аналогичными коэффициентами МЗ и ДЗ близнецов статистически достоверны. В группе HP параметры альфа-ритма оказываются совершенно разными: значимые положительные корреляции вообще отсутствуют.
Что же можно сказать о межзональных различиях в наследуемости альфа-ритма? Наиболее высокие коэффициенты внутрипарного сходства дают затылочные отведения, а самые низкие характерны для ЭЭГ
297
Рис. 13.2. Электроэнцефалограммы: а — МЗ близнецов; б — ДЗ близнецов [132, гл. Ш].
левого височного отведения. Целесообразно обратить внимание на соотношение величин коэффициентов внутриклассовой корреляции: в Т3 он у МЗ близнецов намного ниже, чем в любом другом отведении, а у ДЗ близнецов это отведение дает один из наиболее высоких коэффициентов, т. е. разница во внутрипарном сходстве МЗ и ДЗ очень мала, коэффициент наследуемости, по Игнатьеву, равняется всего 0,28. Дополнительную информацию дает генетико-статистический анализ. Разложение фенотипической дисперсии амплитуды альфа-ритма приведено в табл. 13.3, которая показывает, что доля аддитивного генетического компонента по амплитуде альфа-ритма весьма велика (от 57 до 96%). Наиболее высок вклад генотипической составляющей в межиндивидуальную дисперсию амплитуды альфа-ритма в затылоч-
298
-0.4.
Рис. 13.3. Коэффициенты внутриклассовой корреляция по параметрам альфа-ритма: 1 — амплитуда; 2 — индекс; 3 — частота; между диаграммами дана схема отведений; р<0,05 для гю>гв(отмечено звездочкой). «-М3;д=з -ДЗ; ш-НР.
ных и лобных отведениях (95-96%), а самый низкий — в левом височном и правых центральном и теменном (57-60%), причем здесь дисперсия признака за счет средовых влияний в значительной мере определяется факторами систематической среды (21-32,5%).
Такое же разложение по альфа-индексу в основном повторяет данные, полученные для амплитуды. Доля генотипической составляющей, если исключить левое височное отведение, колеблется от 63 до 96%. Наиболее высокие величины характерны для затылочной ЭЭГ (91—96%). Наибольшие средовые влияния — случайные и систематические (40 и 14% соответственно) — выявлены, как и для альфа-амплитуды, в левом височном отведении.
Итак, налицо межполушарные различия в степени генетических влияний, и прежде всего большая подверженность действию средовых факторов некоторых параметров ЭЭГ височной зоны левого полушария. Хотя в общем параметры альфа-ритма довольно жестко детерминированы генотипом, ЭЭГ левого височного отведения по сравнению с другими зонами имеет в парах МЗ близнецов гораздо меньшее
299
Таблица 13.3
Коэффициенты внутриклассовой корреляции и разложение (в %) фенотипической дисперсии амплитуды альфа-ритма [132, гл. III]
| Отведени я | Гмз | Гдз | VА | VD | VW | VC | х2 | P |
| Т4 | 0,85 | 0,44 | 78 | — | 15 | 1 | 6,8 | <0,01 |
| Т3 | 0,64 | 0,50 | 57 | — | 19 | 24 | 1,11 | 0,25-0,50 |
| F4 | 0,95 | 0,45 | 95 | — | 5 | — | 4,06 | 0,10-0,20 |
| F3 | 0,96 | 0,39 | 96 | — | 4 | — | 3,1 | 0,10-0,25 |
| Р4 | 0,92 | 0,61 | 60 | — | 7,5 | 32,5 | 0,62 | 0,25-0,50 |
| Р3 | 0,95 | 0,51 | 85 | — | 4,5 | 10,5 | 0,91 | 0,25-0,50 |
| О2 | 0,96 | 0,33 | 95 | — | 5 | — | 1,94 | 025-0,50 |
| | 0,97 | 0,34 | 96 | — | 4 | — | 3,40 | 0,1-0,25 |
| С4 | 0,88 | 0,49 | 60 | — | 19 | 21 | 0,98 | 0,25-0,50 |
| С3 | 0,82 | 0,40 | 82 | — | 16,5 | 1,5 | 0,56 | 0,25-0,50 |
Обозначения: Т — височное; F — лобное; Р — теменное; О — затылочное; С — центральное; четные номера — правое полушарие, нечетные — левое; генетические составляющие: VA— аддитивная, VD — доминантная (в данном разложении оказалась непредставленной); средовые составляющие: VW— случайная (индивидуальная), VC— систематическая (общая); r— коэффициент внутриклассовой корреляции.
сходство. На рис. 13.3 представлены коэффициенты внутриклассовой корреляции по параметрам альфа-ритма (амплитуде, альфа-индексу и частоте), из которых отчетливо видно, что внутрипарное сходство МЗ близнецов по всем трем перечисленным параметрам в левом височном отведении (третий по счету столбик) меньше, чем для всех остальных отведений левого полушария.
Сравнительно меньшая генотипическая обусловленность параметров альфа-ритма левой височной области объясняется, по мнению Т.А. Мешковой, относительно молодым филогенетическим возрастом височной области коры, длительным периодом ее созревания в онтогенезе, а также особой ролью в осуществлении речевых функций.