
Федеральная программа книгоиздания России Рецензенты: канд психол наук С. А. Исайчев, доктор биол наук И. И. Полетаева Равич-Щербо И. В. и др. Р12
| Вид материала | Программа |
СодержаниеПрирода межиндивидуальной Природа межиндивидуальной Индивидуальная специфичность и стабильность вп и ссп 2. Вп и ссп как объекты генетического исследования Влияние особенностей стимула |
- Вестник балтийской педагогической академии вып. 94. – 2010 г. Актуальные проблемы нравственного, 2431.92kb.
- Рецензенты: профессор, доктор психол наук Филонов Л. Б., вед науч сотрудник, канд психол, 2609.63kb.
- Научный выпуск вестник балтийской педагогической академии вып. 29. – 2000 г. Поиск, 1745.18kb.
- Общеобразовательная программа дошкольного образования Авторский коллектив, 5619.19kb.
- Образовательная программа дошкольного образования Москва «Просвещение», 5670.3kb.
- Введенским Игорем Витальевичем Рецензенты доктор психол наук В. А. Лабунская канд психол, 375.9kb.
- Отчет о проведении Международной научной конференции-семинара «Современные методы психологии», 97.76kb.
- Приглашение и программа разнообразие почв и биоты северной и центральной азии, 521.14kb.
- Пояснительная записка, 12621.4kb.
- В организации совместных отношений, 1137.14kb.
Общий паттерн ЭЭГ, а также основные количественные параметры ЭЭГ относятся к числу индивидуально устойчивых особенностей человека, что дает основания изучать роль генотипа и среды в происхождении межиндивидуальной вариативности по этим признакам. Результаты большинства генетических работ свидетельствуют о значительном влиянии наследственных факторов на общий рисунок ЭЭГ. Для нескольких редко встречающихся вариантов ЭЭГ установлен тип наследования (аутосомно-доминантный), выделен ген, ответственный за один из этих вариантов (низковольтная ЭЭГ).
При анализе ритмических составляющих ЭЭГ показано значительное влияние генотипа на параметры альфа-ритма, но с существенными межзональными и межполушарными различиями. Использование автоматического спектрального анализа ЭЭГ позволяет выявить высокую степень наследственной обусловленности и для других ритмов ЭЭГ. Установлено, что как структура спектра ЭЭГ покоя, так и соотношение случайных и периодических составляющих в нем испытывают на себе значительное влияние со стороны генотипа.
Генетическая обусловленность характерна не только для ЭЭГ в состоянии покоя, но и для реактивных изменений, возникающих при сенсорной стимуляции, а также при приеме алкоголя.
Оценки наследственной обусловленности ЭЭГ могут варьировать в зависимости от зоны регистрации и исследуемого параметра ЭЭГ, уровня бодрствования и возраста испытуемых.
Глава XIV
ПРИРОДА МЕЖИНДИВИДУАЛЬНОЙ ВАРИАТИВНОСТИ БИОЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ МОЗГА: ВЫЗВАННЫЕ ПОТЕНЦИАЛЫ
1. ВЫЗВАННЫЕ ПОТЕНЦИАЛЫ КАК МЕТОД ИЗУЧЕНИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ МОЗГА. ОПИСАНИЕ И СПОСОБЫ АНАЛИЗА ВЫЗВАННЫХ ПОТЕНЦИАЛОВ
Вызванные потенциалы (ВП) — биоэлектрические колебания, возникающие в нервных структурах в ответ на внешнее раздражение и находящиеся в строго определенной временной связи с началом его действия. Стимулами могут служить относительно элементарные, поддающиеся количественной оценке раздражители, например, вспышки определенной интенсивности, шахматные поля с разным разме-
309
* * *
Общий паттерн ЭЭГ, а также основные количественные параметры ЭЭГ относятся к числу индивидуально устойчивых особенностей человека, что дает основания изучать роль генотипа и среды в происхождении межиндивидуальной вариативности по этим признакам. Результаты большинства генетических работ свидетельствуют о значительном влиянии наследственных факторов на общий рисунок ЭЭГ. Для нескольких редко встречающихся вариантов ЭЭГ установлен тип наследования (аутосомно-доминантный), выделен ген, ответственный за один из этих вариантов (низковольтная ЭЭГ).
При анализе ритмических составляющих ЭЭГ показано значительное влияние генотипа на параметры альфа-ритма, но с существенными межзональными и межполушарными различиями. Использование автоматического спектрального анализа ЭЭГ позволяет выявить высокую степень наследственной обусловленности и для других ритмов ЭЭГ. Установлено, что как структура спектра ЭЭГ покоя, так и соотношение случайных и периодических составляющих в нем испытывают на себе значительное влияние со стороны генотипа.
Генетическая обусловленность характерна не только для ЭЭГ в состоянии покоя, но и для реактивных изменений, возникающих при сенсорной стимуляции, а также при приеме алкоголя.
Оценки наследственной обусловленности ЭЭГ могут варьировать в зависимости от зоны регистрации и исследуемого параметра ЭЭГ, уровня бодрствования и возраста испытуемых.
Глава XIV
ПРИРОДА МЕЖИНДИВИДУАЛЬНОЙ ВАРИАТИВНОСТИ БИОЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ МОЗГА: ВЫЗВАННЫЕ ПОТЕНЦИАЛЫ
1. ВЫЗВАННЫЕ ПОТЕНЦИАЛЫ КАК МЕТОД ИЗУЧЕНИЯ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ МОЗГА. ОПИСАНИЕ И СПОСОБЫ АНАЛИЗА ВЫЗВАННЫХ ПОТЕНЦИАЛОВ
Вызванные потенциалы (ВП) — биоэлектрические колебания, возникающие в нервных структурах в ответ на внешнее раздражение и находящиеся в строго определенной временной связи с началом его действия. Стимулами могут служить относительно элементарные, поддающиеся количественной оценке раздражители, например, вспышки определенной интенсивности, шахматные поля с разным разме-
309
ром ячейки (зрительные ВП), звуковые тоны заданной частоты и интенсивности (слуховые ВП), слабые удары электрическим током известных параметров (соматосенсорные ВП) [134].
Наряду с этим существующие способы выделения сигнала из шума позволяют отмечать в записи ЭЭГ изменения потенциала, достаточно строго связанные во времени с любым фиксированным событием, в том числе субъективным, «внутренним». Благодаря этому выделился еще один круг физиологических явлений — событийно-связанные потенциалы (ССП). Примерами их служат: колебания, связанные с активностью двигательной зоны коры больших полушарий (моторный потенциал, или потенциал, связанный с движением); потенциал, связанный с намерением произвести определенное действие (так называемая Е-волна); потенциал, возникающий при пропуске ожидаемого стимула.
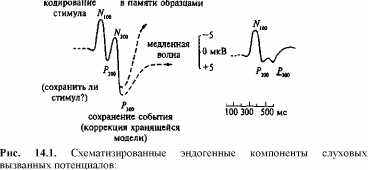
Сенсорные вызванные потенциалы представляют собой последовательность позитивных и негативных колебаний, регистрируемых, как правило, в интервале 0-500 мс. Событийно-связанные потенциалы включают и поздние колебания в интервале до 1000 мс и более. Количественные методы оценки ВП и ССП предусматривают в первую очередь оценку амплитуд и латентнос-тей. При описании их компонентного состава в большинстве случаев указывают полярность компонента (отрицательный, негативный — N; положительный, позитивный — Р) и его порядковый номер от начала ответа или временные параметры. (Напр., позитивное колебание в интервале 300-600 мс обозначается как Р3 или Р300.)
Как правило, компоненты ВП делят на экзогенные и эндогенные. Первые отражают активность специфических проводящих путей, по которым в кору поступают афферентные сигналы, и зон, в которых они обрабатываются. Вторые более тесно связаны с активностью неспецифических ассоциативных систем мозга. Длительность тех и других оценивается по-разному для разных модальностей. Например, в зрительной системе экзогенные компоненты ВП регистрируются в течение первых 100 мс с момента стимуляции.
В интерпретации компонентов и параметров ВП и ССП в настоящее время широко используется понятийный аппарат информационного подхода, при котором вся совокупность реакций такого типа трактуется как отражение процессов приема и переработки информации (рис. 14.1) [62].
ИНДИВИДУАЛЬНАЯ СПЕЦИФИЧНОСТЬ И СТАБИЛЬНОСТЬ ВП И ССП
Высокая межиндивидуальная изменчивость свойственна всем видам электрофизиологических реакций, связанных с внешними воздействиями: сенсорным вызванным потенциалам, потенциалам мозга, связанным с движением (ПМСД), волне ожидания. Коэффициенты корреляции, характеризующие сходство общей конфигурации
310
сравнение с хранящимися
а — ответ на релевантный задаче стимул; б — ответ на иррелевантный стимул [по: 132, гл. IV].
сенсорных ВП различных модальностей и ПМСД в парах неродственников, подобранных по полу и возрасту, составляют в среднем 0,2-0,3 [132, 209, 308], что говорит о выраженных различиях в ВП и ССП любых двух людей. Коэффициенты корреляции, полученные у одних и тех же испытуемых при регистрации ВП и ССП в разные дни, колеблются от 0,6 до 0,9 в зависимости от вида стимула.
Факторами, определяющими индивидуальное своеобразие ВП, предположительно являются уникальные особенности морфологии ЦНС и различных показателей ее функционирования: биохимических, электрофизиологических и пр. ЭЭГ взрослого человека высоко индивидуализирована; устойчивым индивидуально-специфическим
признаком является соотношение «сигнал-шум», на основе которого происходит отделение ВП от фоновой электроэнцефалограммы. Индивидуально специфична и относительно стабильна пространственная картина биоэлектрических колебаний.
Существенным фактором в межиндивидуальной изменчивости ВП и ССП являются половые различия. Наиболее изучены в этом отношении сенсорные ВП. У женщин отмечаются более короткие латентные периоды и более высокая амплитуда зрительных, слуховых и со-матосенсорных ВП [132, 134].
Вместе с тем имеются данные, подтверждающие возможность генетического контроля половых различий по амплитудам ВП [цит. по: 132]. При параллельном сопоставлении зрительных и слуховых ВП трех групп испытуемых — здоровых мужчин и женщин, а также пациентов с кари-
11
отипом 45Х0 (фенотипически это женщины) — были выявлены большие значения амплитуд в двух последних группах. По-видимому, в отсутствии Y-хромосомы формирование ВП идет по женскому типу.
Несмотря на то что во многих исследованиях отмечается индивидуальное своеобразие ВП и ССП, не выделено каких-либо особых типов этих реакций, свойственных тем или иным группам людей, как это, например, было показано Ф. Фогелем применительно к ЭЭГ (см. гл. XIII). Однако были установлены достоверные различия в амплитудах и латен-тностях зрительных и слуховых ВП у индивидов с разными вариантами ЭЭГ. С точки зрения Ф. Фогеля, это говорит о связи индивидуальных различий в переработке информации с генетически обусловленным признаком — паттерном ЭЭГ. Такой подход открывает новую перспективу для изучения биологических основ поведения в целом.
Тем не менее в общем определенные принципы деления ВП на типы по каким-либо фиксированным особенностям этих реакций не выделены. Как правило, индивидуальные различия сводятся к особенностям компонентного состава ВП и различиям в амплитудах и латентностях.
Исключение составляет феномен «увеличения—уменьшения» (augmenting-reducing) [217, 218]. Он заключается в следующем: с увеличением интенсивности стимула амплитуда ВП сначала увеличивается, а затем, несмотря на продолжающееся усиление стимула, либо стабилизируется, либо снижается. Однако указанная закономерность проявляется по-разному у разных испытуемых. Наиболее отчетливо индивидуальные различия обнаруживаются при высоких значениях интенсивности стимуляции: у испытуемых-«увеличителей» амплитуда ВП продолжает увеличиваться, у испытуемых-«уменьшителей» — уменьшаться. Таким образом, индивидуально-специфическим признаком здесь служит крутизна возрастания линии, отражающей зависимость амплитуды ВП от интенсивности стимула. Ретестовая надежность данного показателя относительно велика, корреляции между результатами повторных исследований одних и тех же испытуемых составляют 0,6-0,8.
Предполагается, что этот феномен отражает функционирование механизмов индивидуальной адаптации при переработке сенсорного опыта. Он обнаруживает связь с некоторыми психологическими показателями: интеллектом, когнитивными стилями, особенностями темперамента и личностными характеристиками. Например, тенденция к ослаблению ответа связана с ин-тровертированностью и более низкими значениями по шкале «поиска ощущений». Кроме того, феномен «увеличения-уменьшения» связан с биохимической индивидуальностью человека, в частности с особенностями метаболизма медиаторов из группы катехоламинов [460]. По мнению М. Закермана, сильная индивидуальная выраженность «охранительных» механизмов проявляется на уровне электрофизиологических реакций в виде ослабления ответов, а в поведении — в склонности к избеганию стимуляции.
В заключение следует сказать, что ВП представляют собой уникальный инструмент для изучения генотип-средовых соотношений в индивидуальных особенностях физиологических механизмов переработки сенсорной информации. Они сочетают в себе все условия, не-
312
обходимые для такого исследования: 1) ВП в целом рассматриваются как электрофизиологический коррелят информационного процесса; 2) компонентная структура ВП соотносима с отдельными этапами, или стадиями, процесса переработки информации; 3) благодаря региональной специфичности ВП дают возможность оценить вклад генотипа в особенности функционирования различных зон мозга; 4) ВП относятся к числу индивидуально-специфических реакций, параметры которых характеризуются непрерывной изменчивостью, что позволяет ставить вопрос о роли генотипа в происхождении этой изменчивости и использовать для его решения методы биометрического анализа.
2. ВП И ССП КАК ОБЪЕКТЫ ГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
СЕНСОРНЫЕ ВЫЗВАННЫЕ ПОТЕНЦИАЛЫ
Первые генетические исследования ВП появились в 60-х годах, с тех пор их было проведено в общей сложности около 20 [см.: 132,431). В подавляющем большинстве они выполнены методом близнецов, и для них характерны те же особенности, которые отличают генетические исследования ЭЭГ, т.е. относительно небольшое количество пар и большой возрастной диапазон испытуемых, а также разные условия регистрации ЭЭГ и стимуляции.
В то же время в ряде работ используются сходные принципы постановки исследования и последующей обработки данных. Так, в нескольких случаях оценка внутрипарного сходства в парах МЗ и ДЗ близнецов проводилась по волновой форме ВП, т.е. по цифровому ряду, описывающему конфигурацию ВП с шагом дискретизации в несколько миллисекунд. Полученные цифровые ряды или их фрагменты в каждой паре близнецов подвергали корреляционному сопоставлению. Для получения усредненных по группам значений коэффициентов корреляции использовались z-преобразования индивидуальных коэффициентов с последующим усреднением их. Оценка достоверности различий между усредненными значениями z может проводиться с помощью Т-критерия Стьюдента, применяется также однофакторный дисперсионный анализ и некоторые другие варианты анализа. Сопоставление волновых форм дает наиболее общую оценку сходства потенциалов, однако не учитывает, за счет каких именно компонентов возникает сходство. Необходимо также иметь в виду, что при достаточно сходной форме двух ВП небольшой сдвиг латентных периодов может значительно снизить коэффициенты внутрипарного сходства.
В связи с функциональной неоднородностью ВП особое значение имеет генетический анализ отдельных компонентов ВП и их параметров. Использование некоторых формальных критериев позволяет выя-
вить наиболее характерную для данных условий последовательность компонентов ВП. При этом латентные периоды вычисляются от момента подачи стимула до пика соответствующего компонента. Оценка амплитуд может проводиться как от средней линии, так и от пика до пика. Возможно также использование более сложных алгоритмов вычисления амплитудных параметров ВП [244, 316], а также топографическое картирование параметров ВП [338].
Генетический анализ ВП целесообразно начинать с оценки парной конкордантности МЗ и ДЗ близнецов по компонентной структуре ВП, а уже затем в парах конкордатных близнецов устанавливать меру генотипической обусловленности амплитудно-временных параметров ВП. В большинстве работ оценка сходства проводилась с помощью коэффициентов корреляции соответствующих показателей. В некоторых исследованиях наследуемость отдельных параметров оценивалась с помощью методов биометрической генетики с разложением фенотипической дисперсии на компоненты и подбором моделей [132,346].
Анализ гено- и паратонической обусловленности ВП и ССП целесообразно проводить отдельно в трех ракурсах в зависимости от: 1) особенностей стимула (модальности, интенсивности, качественных характеристик); 2) особенностей экспериментальной ситуации; 3) анализируемого компонента (его параметров и места в общей структуре ВП) и области его регистрации. Рассмотрим указанные направления анализа.
ВЛИЯНИЕ ОСОБЕННОСТЕЙ СТИМУЛА
Наиболее существенной характеристикой стимула является его модальность, т.е. принадлежность к определенной сенсорной системе. По этому признаку выделяются зрительные, слуховые и соматосен-сорные ВП. Закономерно возникает вопрос: какое влияние оказывают факторы генотипа на формирование межиндивидуальной изменчивости ВП в каждой модальности?
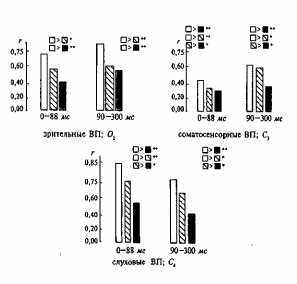
Роль факторов генотипа и среды наиболее изучена для зрительных, слуховых и, в меньшей степени, соматосенсорных ВП. Влияния генотипа неоднократно выявлялись в изменчивости ВП на вспышки умеренной интенсивности и звуковые тоны и щелчки в основном в диапазоне 60—80 дБ. Наиболее полное исследование было проведено Э. Льюисом с соавторами [316]. В нем принимали участие 44 пары МЗ, 44 пары ДЗ близнецов и 46 пар HP от 4 до 40 лет и фактически впервые было показано, что генотип вносит свой вклад в изменчивость волновой формы и амплитудных параметров ВП на зрительные и слуховые стимулы (рис. 14.2).
В этой работе анализировались также соматосенсорные ВП на электрокожное раздражение правого указательного пальца с интенсивно-
314
Рис. 14.2. Усредненные коэффициенты корреляции, полученные при сравнении волновой формы ВП.
В каждой из трех групп (МЗ, ДЗ, HP) волновую форму попарно коррелировали, а затем усредняли (М6|. П - МЗ; Q- ДЗ; ■ - HP; • - р > 0,05; •• - р > 0,01.
стью, в 2 раза превышающей субъективный порог. По волновой форме соматосенсорных ВП статистически достоверных различий между МЗ и ДЗ получено не было. Изучение амплитудных показателей позволило выявить генетические влияния в изменчивости соматосенсорных ВП, но они все равно оказались меньше, чем в зрительных и слуховых ВП.
Следующей характеристикой стимула является его интенсивность. В каждой модальности стимулы могут быть слабыми, умеренными и сильными, соответственно амплитуда ВП изменяется в зависимости от интенсивности стимула. Если стимулы (вспышка, звуковой тон) слабые, то ВП имеют небольшую амплитуду. При усилении стимула амплитуда компонентов ВП возрастает, но только до определенного предела, различного у разных индивидов. По данному признаку они делятся на «уменьшителен» и «увеличителей»: у первых этот предел наступает раньше, чем у вторых.
Закономерно возникает вопрос: зависят ли проявления генетического контроля ВП от силы раздражителя? Судя по некоторым данным, генетическая обусловленность амплитуды проявляется по-раз-
15
ному в ответах на стимулы различной силы [ 132, гл. IV]. В слуховых ВП увеличение интенсивности раздражителя приводит к более отчетливому проявлению генотипического контроля амплитуд. Различия в факторах, формирующих изменчивость ВП на тоны 80 дБ и 105 дБ, анализировались Б.И. Кочубеем [84]. Влияние генотипа отчетливо сказывалось в амплитудах компонентов N1 Р2 (интервал 100-200 мс): показатели наследуемости составили 0,46 и 0,79 в реакции на тон 80 дБ и 0,58 и 0,81 в реакции на тон 105 дБ.
Эта проблема исследовалась также применительно к зрительным и слуховым ВП в связи с изучением генетического контроля феномена «увеличения—уменьшения» [217, 218]. Изучение внутрипарного сходства динамики ВП при изменении интенсивности стимулов (вспышки) у 33 пар МЗ и 34 пар ДЗ близнецов дало внутриклассовые корреляции, свидетельствующие об увеличении наследуемости амплитуды ВП на световые стимулы по мере возрастания яркости стимула. В табл. 14.1 приведены данные для амплитудных показателей, вычисленных в интервале 76-112 мс. Из этих данных следует, что по
Таблица 14.1
Наследуемость амплитудных параметров зрительных ВП
| Амплитудные | Коэффициенты | Коэффициенты | Показатель |
| показатели | корреляции МЗ | корреляции ДЗ | наследуемости |
| При разных интен- | | | |
| сивностях стимула: | | | |
| 1-й уровень | -0,20 | -0,21 | 0,00 |
| 2-й уровень | 0,31 | 0,07 | 0,25 |
| 3-й уровень | 0,40 | 0,02 | 0,39 |
| 4-й уровень | 0,51 | -0,04 | 0,52 |
| Средние амплитуды: | | | |
| Р1 00 -N140 | 0,59 | 0,36 | 0,35 |
| N140 - P200 | 0,57 | 0,10 | 0,52 |
| Крутизна наклона кри- | | | |
| вой при росте интен- | | | |
| сивности стимула: | | | |
| P1 00 -N140 | 0,51 | -0,04 | 0,52 |
| N140 - P200 | 0,56 | -0,10 | 0,60 |