
Гидрофильная флора сосудистых растений как модельный объект для инвентаризации и анализа флоры (на примере тульской и сопредельных областей) 03. 02. 01 ботаника
| Вид материала | Автореферат диссертации |
- Ю. Д. Нухимовская 1, И. А. Губанов 2 в первом Национальном доклад, 745.41kb.
- Флора и фитогеография печеночников (marchantiophyta, anthocerotophyta) камчатки и прилегающих, 596.98kb.
- Итоги интродукции древесных растений североамериканской флоры в челябинской области, 373.53kb.
- «Нарочанский». Анализ синантропного компонента флоры, 701.71kb.
- Видовой состав лекарственных растений местной флоры, 396.37kb.
- Рабочей программы учебной дисциплины ботаника уровень основной образовательной программы, 95.26kb.
- «Национальный характер как объект социально-философского анализа» (на примере таджикской, 352.02kb.
- Рабочая программа дисциплины «ботаника» Код дисциплины по учебному плану опд., 301.75kb.
- Структурные и функциональные свойства цитокинин-связывающих белков растений Arabidopsis, 249.12kb.
- Флора и растительность копаней ярославской области 03. 00. 05 ботаника, 463.62kb.
Всего в водную флору изученной территории включено 216 видов сосудистых растений, относящихся к 94 родам и 47 семействам, из которых 74 слагают ее «водное ядро», 46 — прибрежно-водную компоненту и остальные 96 являются заходящими в воду растениями. 62 вида «водного ядра» флоры являются природными, 1 — адвентивным, полностью натурализовавшимся (Elodea canadensis Michx.) и 11 — адвентивными, не натурализующимися. Среди прибрежно-водных растений видами природной флоры являются 39, адвентивными, не натурализующимися, — 7. Среди заходящих в воду растений — 90 видов природной флоры, 5 — адвентивных, полностью натурализовавшихся, и 1 — адвентивный, не натурализующийся. Кроме того, в «Конспекте флоры» приведено 14 гибридов, входящих в ее «водное ядро», и 1 гибрид, являющийся прибрежно-водным растением (Typha × glauca Godron).
В процессе работ, а также в гербариях было обнаружено 5 таксонов, новых для Центрального федерального округа ( Sagittaria ambigua J. G. Smith, Elodea densa (Planchon) Caspary, Vallisneria americana Michx., Lemna minuta Kunth, Wolffia globosa (Roxb.) Hartog et Plas), 8, ранее не указанных для Московского региона (Potamogeton acutifolius Link, P. nodosus Poir., P. rutilus Wolfg., Sagittaria ambigua, Elodea densa, Vallisneria americana, Lemna minuta, Wolffia globosa), 12 — для Тульской области (Salvinia natans (L.) All., Typha elatior Boernm, Potamogeton acutifolius, P. gramineus L. s.l., P. pusillus L. s.str., Phragmites altissimus (Benth.) Mabille, Eleocharis mamillata Lindb. fil. s.l., Lemna gibba L., Ranunculus kauffmanii Clerc, R. pseudofluitans (Syme) Newbould ex Baker et Foggitt, Lythrum virgatum L., Hottonia palustris L.), 9 — для Рязанской области (Typha laxmannii Lepech., Potamogeton × fluitans Roth, P. × sparganiifolius Laest. ex Fries, Zannichellia palustris L. s.l., Vallisneria spiralis L., Phragmites altissimus, Scirpus tabernaemontani C. C. Gmel., Lemna gibba, Ceratophyllum submersum L.) и 16 — для Орловской области (Potamogeton acutifolius, P. alpinus Balb., P. friesii Rupr., P. nodosus, P. pusillus s.str., P. trichoides Cham. et Schlecht., Zannichellia palustris, Alisma gramineum Lej., Phragmites altissimus, Bolboschoenus planiculmis (F. Schmidt) T. V. Egorova, Lemna gibba, Nuphar × spenneriana Gaud., Ceratophyllum submersum, Elatine hydropiper L., Utricularia intermedia Hayne, Galium trifidum L.). Для ряда таксонов было подтверждено произрастание на территории соответствующих регионов (Caulinia minor (All.) Coss. et Germ — в Тульской и Орловской областях и C. tenuissima (A. Br. ex Magnus) Tzvelev и Nymphaea alba L. — в Рязанской области), в ряде случаев ранее указывавшиеся для регионов таксоны были из их флоры исключены (Elatine alsinastrum L., приводившийся для флоры Орловской области на основании неправильно определенного образца Hippuris vulgaris L.), обнаружены новые местонахождения многих редких и охраняемых видов сосудистой водной флоры регионов, существенно изменены представления о распространении отдельных видов по территории регионов или флористических выделов.
Для сравнения по параметрам видового богатства были взяты «водные ядра» флор (Щербаков, 1991, 2006; Щербаков, Тихомиров, 1994, и др.), поскольку именно слагающие их истинно-водные и земноводные растения в максимальной степени связаны с водной средой обитания и именно их легче всего отграничить от иных экологических групп, входящих в состав флор водоемов.
Наиболее богаты в пределах изученной территории «водные ядра» флор Московского региона и Рязанской области, имеющих наиболее высокие коэффициенты озерности, причем доминирование по этому показателю «водного ядра» флоры Московского региона обеспечивается адвентивными видами, тогда как по природной компоненте оно уступает Рязанской области (табл. 1). «Водные ядра» флор Тульской и Орловской областей значительно беднее и практически равны между собой. Примерно такие же тенденции демонстрируют и «водные ядра» флор сопредельных регионов Средней России, наиболее богатой из которых оказалась имеющая максимальный коэффициент озерности Тверская область.
Таблица 1. Видовое богатство «водных ядер» флор изученных регионов
| Группа таксонов | Регион | |||
| Московский регион | Рязанская область | Тульская область | Орловская область | |
| Водные природные и адвентивные натурализовавшиеся | 48 | 50 | 40 | 38 |
| Земноводные природные | 10 | 10 | 9 | 7 |
| Адвентивные не натурализующиеся | 10 | 1 | 1 | 1 |
| Всего видов | 68 | 61 | 50 | 46 |
| Гибриды | 10 | 8 | | 1 |
Из анализа дислокации местонахождений реликтовых видов на изученной нами территории (причем Salvinia natans и Trapa natans L. s.l., по нашему мнению, здесь являются псевдореликтами), возможности флористического анализа ее региональных водных флор на основе критериев различия для целей флористического районирования крайне ограничены, хотя в дальнейшем в некоторых случаях мы их использовали.
Самым простым способом сравнения флор является прямое сопоставление флористических списков, широко практикуемое в геоботанике. Для этих целей в обычно используют коэффициенты сходства Жаккара (Шмидт, 1980) и Чекановского (Шмидт, 1980): K = 2c : (a + b). Мы также попытались использовать этот метод для флористического районирования изученной нами территории на основе «водных ядер» флоры. Применительно к Московскому региону за основу было взято районирование, приведенное в «Определителе растений Московской области» (Ворошилов и др., 1966), с некоторыми изменениями, сделанными на основе наших натурных наблюдений.
Рисунок 1. Районирование изученных регионов по «водным ядрам» их флор, выполненное на основе метода экспертных оценок
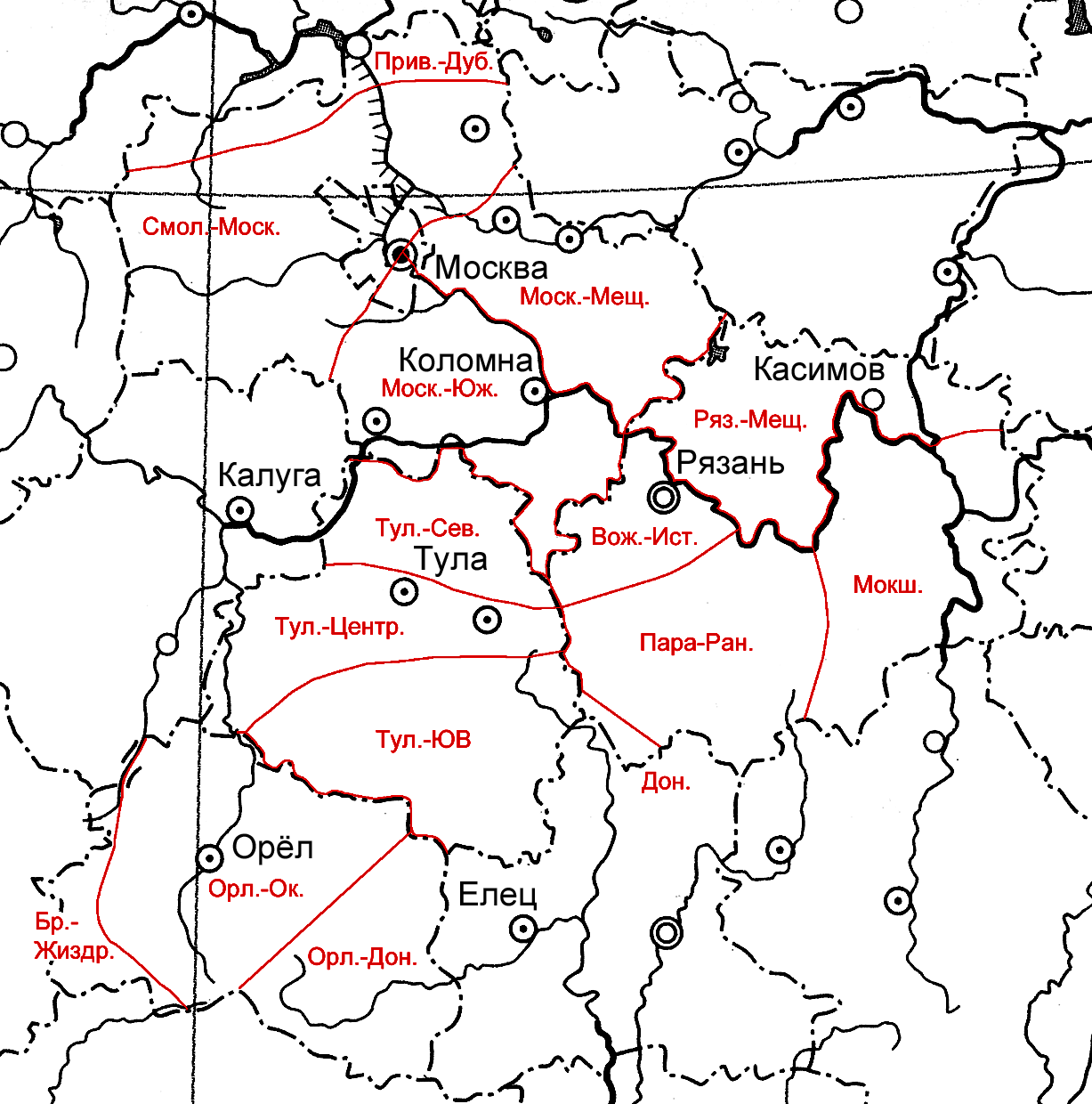
Результаты сравнения «водных ядер» смежных флористических выделов Московского региона на основании коэффициента сходства Жаккара приведены в табл. 2.
Таблица 2. Коэффициенты сходства Жаккара (сверху и справа) и Евклидовы расстояния (см. с. 20) (снизу и слева) для «водных ядер» смежных флористических выделов Московского региона
| | Прив.-Дубн. | Смол.-Моск. | Моск.-Мещ. | Южный |
| Прив.-Дубн. | Х | 0,81 | | |
| Смол.-Моск. | 7,8 | Х | 0,82 | 0,89 |
| Моск.-Мещ. | | 8,8 | Х | 0,81 |
| Южный | | 7,1 | 9,8 | Х |
К сожалению, при использовании данного метода обычно стороной обходится вопрос о полноте выявления видового состава сравниваемых флор. Так, Л. И. Малышев в работе «Флористические исследования на плато Путорана» (1976) пишет: «Обычно требовалось около 20 дней, чтобы выявить видовой состав растений в предельных случаях с полнотой ориентировочно 95—97%», причем данная фраза относится к конкретным флорам площадью около 100 км2. Проведенная нами математически проверка справедливости этого утверждения показала, что оно излишне оптимистично и полнота выявления флоры едва ли превышала 75%. Несмотря на очевидность факта, что реально при флористическом анализе сравнивают не флоры, а лишь более или менее полные выборки из них, во флористической литературе нам не удалось обнаружить практических шагов по учету действия данного фактора на результаты флористического анализа, и мы попытались восполнить этот пробел.
С этой целью мы взяли 400 описаний сосудистой водной флоры Московского региона, равномерно распределенных по его территории. Затем мы разделили эту полную выборку описаний на две, одну из которых составили только четные описания, а другую — только нечетные. В описаниях мы учитывали только природные и полностью натурализовавшиеся виды «водного ядра». Затем эти выборки сравнили друг с другом (табл. 3).
Таблица 3. Сравнение двух выборок «водного ядра» флоры Московского региона с использованием коэффициента сходства Жаккара (Щербаков, 2006, 2010)
| Показатель | Величина |
| Общее число видов «водного ядра» флоры региона во всех 200 нечетных описаниях (а) | 47 |
| Общее число видов «водного ядра» флоры региона во всех 200 четных описаниях (b) | 46 |
| Число видов только в нечетных описаниях | 7 |
| Число видов только в четных описаниях | 6 |
| Число общих видов (с) | 40 |
| Коэффициент сходства Жаккара (К) | 0,75 |
Сравнивая таблицы 2 и 3, можно убедиться, что коэффициенты сходства Жаккара для «водных ядер» флор разных флористических выделов Московского региона оказываются выше, чем для двух взаимодополняющих выборок из «водного ядра», каждая из которых содержит примерно по 85% флоры. Иными словами, анализируя эти таблицы, можно сделать вывод, что в данном случае величины коэффициентов, получаемые при сравнении «водных ядер» флор выделов, на самом деле вполне могут относиться к практически идентичным объектам, а различия между ними быть обусловленными не только реальными различиями между флорами, но и случайными различиями между выборками, то есть быть «ошибкой метода».
Мы попытались определить, какую часть «водного ядра» флоры можно выявить при использованной нами методике региональных флористических исследований. Были составлены математические ряды, показывающие среднее число видов, приходящихся на определенное число описаний для «водных ядер» флор Московской, Рязанской, Калужской и Тульской областей.
Анализ полученных рядов был произведен с помощью программы «CurveExpert 1.3». Данная программа самостоятельно определяет, какие из зависимостей, имеющиеся в ее базе данных, могут в принципе объяснить тот или иной ряд, подбирает к этим зависимостям коэффициенты, определяет достоверность описания ряда и среднеквадратичное отклонение, а затем на основании этих параметров ранжирует модели по убыванию точности объяснения ими анализируемого ряда.
Процесс выявления «водных ядер» региональных флор в нашем случае лучше всего описывали зависимости, не имеющие предела («MMF-Model» из имеющихся в базе данных программы «CurveExpert 1.3» и логарифмическая, являющаяся полным аналогом экологической модели Г. Глизена — y = a + b × lnx (Gleason, 1922, 1925)), что для участков ограниченной площади выглядит нелогичным. Эту нелогичность, с нашей точки зрения, можно объяснить только динамичностью флоры и относительно длительным периодом ее изучения.
Результаты анализа процесса выявления полной флоры на локальном уровне, для чего нами были использованы материалы флористических исследований в национальном парке «Орловское Полесье», были аналогичными (Щербаков и др., 2002). Кроме того, анализ процесса выявления флоры национального парка «Орловское Полесье» показал, что модель Е. Арчибальда — y = b : (c + x–z (Archibald, 1949), как и некоторые другие логистические модели, при описании процесса выявления флоры может быть использована, чтобы по начальной части ряда (не менее 30 описаний, равномерно распределенных по территории, для локальной флоры и 40—50 — для региональной парциальной флоры) попытаться рассчитать число видов, которые одномоментно могут находиться на изучаемой территории.
Если в качестве счетной единицы использовать не вид, а баллы его встречаемости или активности, большее значение при сравнении будет иметь не присутствие в одной флоре очень редкого вида при его отсутствии в другой, а резкое различие встречаемости одного и того же вида в сравниваемых флорах.
С целью проверки этой гипотезы мы сравнили те же самые четыре флористических выдела Московского региона по критерию Евклидова расстояния (Джонгман и др., 1999) — ED = √∑ (kj – ki)2, где ED — Евклидово расстояние, kj — балльная оценка встречаемости вида k во флоре j, ki— балльная оценка встречаемости вида k во флоре i, а также определили ошибку этого метода, связанную с неполнотой анализируемых выборок и случайными различиями между ними.
При сравнении тех же двух выборок из флоры, что и при оценке коэффициента сходства Жаккара (табл. 3), Евклидово расстояние оказалось равным 5,7. Именно эта цифра является ошибкой данного метода.
Сравнение данных из табл. 2 (слева и снизу) с ошибкой метода свидетельствует о достоверности различий «водных ядер» природных флор этих выделов. При этом хорошо заметны различия между «водными ядрами» флор низменностей и возвышенностей, а также относительно меньшее различие по этому показателю «водных ядер» Смоленско-Московской возвышенности и южной части Московской области, где имеется относительно широкая полоса взаимопроникновения флор на стыке Смоленско-Московской возвышенности и Москворецко-Окской равнины, с одной стороны, а также относительно богатая заболоченными участками долина Оки, с другой стороны. В целом же на одном и том же материале данный метод показывает бóльшую точность, и мы применили его для флористического районирования изученной территории по гидрофильной компоненте ее природной флоры (рис. 2).
Анализ современного флористического районирования изученной территории по гидрофильной компоненте ее флоры показал, что оно в основном определяется ее геоморфологическим строением, а также продолжительностью, интенсивностью и характером хозяйственного использования отдельных выделов, причем оценка сходства гидрофильных флор по критерию Евклидова расстояния показывает примерно равную степень участия двух этих факторов.
В нашей работе изначально территория выделялась по политико-административному принципу. Как отмечалось ранее, изучив территорию каждого из регионов, мы на основе существующих схем районирования и собственных наблюдений составляли рабочую схему флористического районирования территории, а затем математически на основе определения Евклидовых расстояний и использования критериев различия совершенствовали ее, приводя к окончательному виду. При этом велось сравнение флористических выделов, а не административных регионов, которые в данном случае выступали лишь в качестве элементов организации исследований, а сама работа напоминала сборку мозаики в известной игре «Puzzles».
Рисунок 2. Флористическое районирование изученной территории по «водному ядру» ее флоры, выполненное на основе вычислений Евклидовых расстояний, дополненное использованием критериев различия (для участком малой площади, границы которых даны пунктиром)
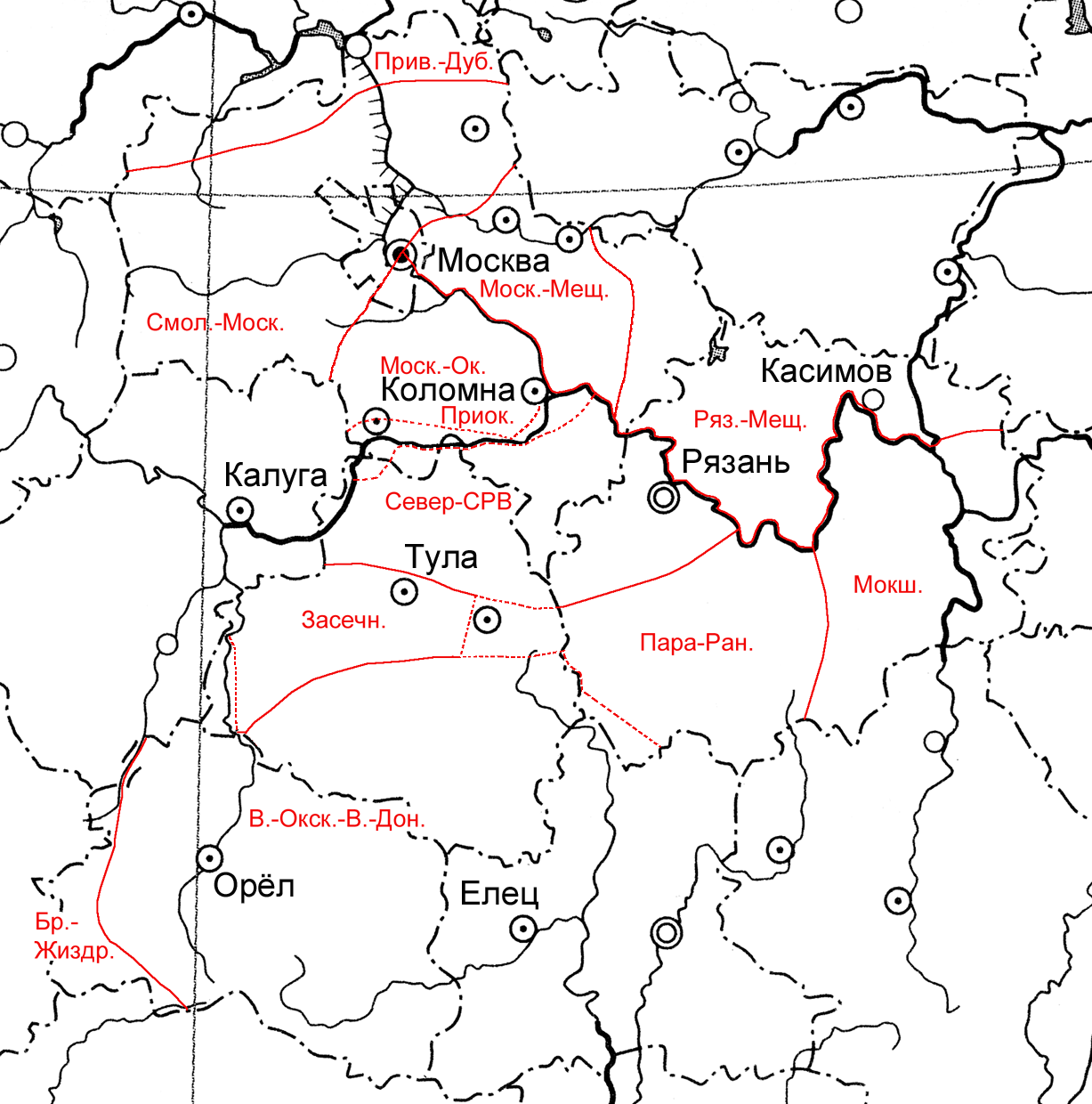
Точно так же можно исследовать территорию, выделяя топографические контуры на основе бассейнового подхода, изучая флору водных объектов слагающих ее геоморфологических образований, или используя какие-либо другие подходы к районированию. Иными словами, принципы выбора выделов регионального ранга, слагающих ту или иную исследуемую территорию (геоморфологический, административно-территориальный, бассейновый или какой-то еще), применительно к решению задачи создания флористического районирования или его уточнения не имеют существенного значения. Важно лишь, чтобы эти выделы были обследованы достаточно квалифицированно, работы были бы выполнены методически грамотно, отсутствовали бы пропуски крупных участков, а данные по отдельным выделам могли бы быть объективно сопоставлены друг с другом.
Неполнота флористических данных, необходимых для анализа, может быть нивелирована обращением к производным параметрам, то есть анализом не отдельных видов, а флористических спектров (систематического, ботанико-географического, эколого-биоморфологического и др.) (Толмачев, 1974; Малышев, 1976, и др.). Для количественной оценки сходства систематических спектров О. В. Ребристой, В. М. Шмидтом (Ребристая, Шмидт, 1972) и Л. И. Малышевым (1976) было предложено использовать коэффициент ранговой корреляции Кендела, или тау-коэффициент. В настоящее время в аналитической части отечественных флористических работ приведение систематического спектра флоры стало каноническим, что, в частности, нередко вело к его некритическому распространению на локальные (не конкретные!) и парциальные флоры. В связи с этим мы решили проверить данный метод и попытаться выявить пределы его использования. В качестве модельных объектов нами, с одной стороны, были взяты флоры Тульского региона (Цингер, 1885; Шереметьева и др., 2008), современные флоры Рязанской и Липецкой областей, а также наши данные по водной флоре. Выяснилось, что в существующем виде таксономический анализ флоры в значительной степени исчерпал себя и для его совершенствования требуется поиск каких-то кардинальных решений:
а) на систематический спектр полной региональной флоры в староосвоенных районах влияет не только ботанико-географическое положение и история развития его флоры, но и степень изученности их адвентивной флоры;
б) на равнинных территориях Средней России систематический спектр природной компоненты флоры определяет ее зональное положение в пределах Бореальной флористической области, а потому нового знания не несет, поскольку такие спектры на Северную Евразию ранее уже были составлены Л. И. Малышевым (1972);
в) степень изученности адвентивной компоненты флоры может быть легко определена через число входящих в нее видов и едва ли требует дополнительных математических расчетов;
г) применительно к парциальным флорам данный метод практически неприменим из-за недостаточности выборки облигатно связанных с определенными группами местообитаний видов и сильной «размытости» границы между заходящими на эти местообитания видами и прочими видами флоры, что ведет к ненужному «обогащению» парциальной флоры относительно чуждыми для нее видами.
Из положительных выводов укажем на допустимость включения агриофитов в природную компоненту флоры для последующего сравнительно-флористического анализа.
Глава 4. Ботанико-географический
и эколого-биоморфологический анализ флоры
Как и в случае систематического анализа, существенное влияние на результаты ботанико-географического анализа флоры оказывает принимаемый в исследовании объем таксонов. Причем на результатах ботанико-географического анализа исключительно негативное влияние могут оказать неверные определения типов ареалов «мелких» видов, связанные с неполнотой данных об их истинном распространении.
Ареалы всех видов, приведенных в «Конспекте флоры» были составлены нами самостоятельно на основе ряда недавно вышедших многотомных академических сводок, а также материалов многочисленных региональных работ, причем оценивались не все области распространения таксонов, а только их ареалы повышенной встречаемости (Щербаков, 1991), которые мы строили на основе формальных количественных показателей региональной (или локальной) встречаемости отдельных видов.
Изученная нами территория находится близ границы умеренных гумидных областей с умеренными аридными. В связи с этим мы сгруппировали все ареалы видов в три блока: умеренных гумидных, умеренных аридных и плюризональных микротермных. На нашу территорию проникают немногочисленные плюризональные макротермные виды (Salvinia natans, Najas major, Trapa natans), однако выделять отдельную группу ареалов в данном случае мы сочли нецелесообразным и объединили умеренные аридные и плюризональные макротермные виды в единую группу видов южного тяготения. Виды, имеющие умеренные гумидные ареалы, в данной работе мы называем видами северного тяготения, а плюризональные микротермные — просто плюризональными. Полностью натурализовавшуюся Elodea canadensis, приняв ее ареал за неокосмополитный, мы включили в группу плюризональных видов.
В природной компоненте «водного ядра» флор изученных регионов имеются постоянная и переменная составляющие. Первая представлена плюризональными видами, которых в «водных ядрах» флор отдельных регионов от 24 до 27, а отдельных флористических выделов — 24—26. Различия в видовом богатстве «водных ядер» флор регионов и отдельных выделов обеспечиваются узкоареальными и адвентивными не натурализующимися видами. По нашим данным, из-за разной величины флор в качестве счетных единиц лучше использовать не виды, а суммы их встречаемостей, а в качестве показателей — не абсолютные значения (суммы встречаемостей видов), а относительные (доли сумм встречаемостей).
Если вместо показателя видового богатства мы возьмем суммы встречаемостей видов, сходство между разными регионами и флористическими выделами по плюризональным видам будет еще более заметным (табл. 4), отражая сравнительное обилие и разнообразие типов водных объектов, а также степень развития в них сосудистых водных растений.
Таблица 4. Участие видов разных ботанико-географических групп в сложении «водного ядра» природной флоры изученных регионов
| Географическая группа | Область или регион | |||
| Московский | Рязанская | Тульская | Орловская | |
| Общая сумма встречаемостей | 189 | 197 | 149 | 147 |
| Плюризональные, сумма встречаемостей | 124 | 117 | 109 | 104 |
| Плюризональные, доля сумм встречаемостей, % | 66 | 60 | 73 | 71 |
| Северного тяготения, сумма встречаемостей | 44 | 39 | 22 | 19 |
| Северного тяготения, доля сумм встречаемостей, % | 23 | 19 | 15 | 13 |
| Южного тяготения, сумма встречаемостей | 21 | 41 | 18 | 24 |
| Южного тяготения, доля сумм встречаемостей, % | 11 | 21 | 12 | 16 |
При движении с севера на юг и юго-восток суммы встречаемостей видов северного тяготения постепенно убывают, как и их доли в «водном ядре» флор, а видов южного тяготения — увеличиваются. При этом уменьшение доли видов северного тяготения особенно заметно по линии север—юг (от Приволжско-Дубнинского флористического района Московской области к Верхнеокско-Верхнедонскому), а увеличение доли видов южного тяготения — по линии северо-запад—юго-восток (от Приволжско-Дубнинского флористического района Московской области к Мокшинскому флористическому району Рязанской области). Данные показатели изменяются не линейно и не постоянно, а лишь демонстрируют некоторые тенденции к этому. Дело в том, что распределение видов различных ботанико-географических групп определяется не только климатом, но и рельефом территории, почвообразующими и подстилающими их породами, а также особенностями и интенсивностью хозяйственного использования территории. Сходные закономерности демонстрирует и распределение по изученным регионам видов прибрежно-водных растений.
Выявленные различия в видовом богатстве и видовом составе «водных ядер» флор отдельных регионов и флористических выделов связаны со степенью разнообразия имеющихся на этих территориях экотопов (в нашем случае — водных объектов). Проведенный по данному параметру анализ показал, что природные водные объекты, как правило, имеют большее видовое богатство, чем водные объекты искусственные, что вполне логично объясняется значительно более долгими сроками их существования и возможностями развития в них растительного покрова. Стоячие природные водоемы, как правило, имеют большее видовое разнообразие, чем природные водотоки.
Плюризональные виды по своему числу равномерно распространены по разным типам водных объектов: в крупных природных водных объектах (реках и озерах) их число практически постоянно (23—24 вида), а в искусственных (пруды и водохранилища) также различается незначительно (22—24 вида). Если в природных типах водоемов на долю плюризональные виды приходится около половины состава их флоры (42—51%), то в реках этот показатель повышается почти до 2/3 (64%). В искусственных водоемах доля плюризональных видов больше половины — от 57% в декоративных и рекреационных прудах до почти 4/5 (78%) в прудах прочих типов.
Видовое богатство «водного ядра» флоры изученных регионов в основном оказывается связанным с обилием природных водоемов: материковых и пойменных озер. При этом виды северного тяготения в основном сосредоточены в материковых озерах и обводненных болотах (около трети видового богатства «водных ядер» флор этих экотопов), а южного тяготения — в пойменных озерах и эфемерных водоемах (также около трети видового богатства «водных ядер» флор). Среди искусственных водоемов доля водных и земноводных видов северного тяготения выше всего в водохранилищах, декоративных и рекреационных прудах.
Из искусственных водоемов сходный с материковыми озерами аридных и семиаридных областей гидрологический режим имеют водохранилища-водонакопители рыбхозов и водопроводных систем, водохранилища гидроэнергетического назначения, пруды, предназначенные для полива и водопоя скота. С эфемерными водоемами определенное сходство демонстрируют ежегодно облавливаемые путем водоспуска рыбоводные пруды. а также пересыхающие в засушливые годы пруды противопожарные.
Характерной чертой «водного ядра» флоры является его абсолютная или относительная бедность адвентивными видами. Помимо Elodea canadensis, в природные сообщества успешно внедрились и фактически стали элементами природной флоры изученной нами территории восточноазиатский Acorus calamus L. и североамериканские Epilobium adenocaulon Hausskn. и Bidens frondosa L.
Адвентивные водные и некоторые прибрежно-водные растения, проникающие в нашу флору, можно разделить на две основные группы: а) интродуцируемые в открытые водоемы с той или иной целью, а затем большее или меньшее время удерживающиеся в местах интродукции (Eichhorrnia crassipes (Mart.) Solms, Pistia stratiotes L., Nymphoides peltatum (S. G. Gmel.) O. Kuntze, Zizania sp. и др.); б) аквариумные растения, попадающие в водные объекты с поступлением термальных вод (Elodea densa, Vallisneria sp., Wolffia globosa и др.). Вид в пределах одного и того же региона может одновременно быть и природным, и заносным. На рассматриваемой территории таковыми являются, например, Scirpus tabernaemontani (в Московском регионе и Тульской области), Lythrum virgatum (природный — в Рязанской области и адвентивный в Московской и Тульской), Trapa natans (в Московской области) и некоторые другие.
В настоящее время в Средней России мы наблюдаем наложение цикла флуктуационного изменения климата, имеющего, с нашей точки зрения, скорее природную причину, и процесса перманентной трансформации водных объектов и их окружения в результате хозяйственной деятельности. Только флуктуационными изменениями климата мы можем объяснить расширение ареала и повышение численности таких видов, как Potamogeton nodosus и Najas major All. Вероятно, исключительно природным причинам мы обязаны и повышению встречаемости Caulinia minor в Московской и Рязанской областях.
Следующая группа видов причиной своего прогресса в Средней России обязана не только природным факторам, но и хозяйственной деятельности. Водохранилища гидроэнергетического, водопроводного и рыбохозяйственного назначения, а также пруды, предназначенные для полива и водопоя скота имеют гидрологический режим, сходный с таковым в материковых озерах аридных и семиаридных областей, что способствует внедрению в них таких видов южного тяготения, как Zannichellia palustris, Alisma gramineum, Ceratophyllum submersum, Typha laxmannii и некоторых других.
Создание мелководных, хорошо прогреваемых противопожарных прудов, а также небольших карьеров различного назначения благоприятно сказываются на расширении ареала и увеличении численности таких теплолюбивых видов, как Potamogeton acutifolius и P. trichoides. Водоемы с поступлением в них термальных вод способствуют еще более успешному распространению по Средней России Lemna gibba и адвентивной L. minuta. Прекращение в Рязанской области промыслового облова пойменных озер обметывающими орудиями лова благотворно сказалось на обитающих в них популяциях Trapa natans.
Некоторые галофильные виды прибрежно-водных и заходящих в воду растений ( Typha laxmannii, Scirpus tabernaemontani и др.), проникают в нашу флору по придорожным кюветам и затем большее или меньшее время удерживаются в них.
Вероятно, природными причинами вызвана исключительная редкость Caulinia tenuissima, являющейся «затухающим» видом. Причины резкого сокращения численности и встречаемости в Московской области Montia fontana L., видимо, также носят природный характер.
За последнее столетие существенно уменьшили свою встречаемость и численность виды материковых озер гумидных областей и обводненных сфагновых болот. В наиболее угрожаемом состоянии находятся виды олиго- и мезотрофных озер, сохранившиеся до наших дней лишь благодаря относительной малодоступности мест их произрастания, такие как Isoёtes echinospora Durieu, I. lacustris L., Sparganium angustifolium Michx., S. gramineum Georgi. Антропогенные аналоги данных местообитаний отсутствуют и в случае эвтрофикации или иных негативных воздействий на экосистемы озер, где эти виды пока сохранились, данные популяции будут обречены на гибель.
Некоторые виды северного тяготения, не являющиеся исключительно лимнофильными и способные произрастать в водотоках ( Potamogeton alpinus и P. praelongus Wulfen), в настоящее время демонстрируют тенденцию некоторого расширения своих ареалов, однако число угрожаемых видов «водного ядра» природной флоры изученной территории превышает число прогрессирующих видов.
Эколого-биоморфологический анализ флоры показывает, что если среди истинно водных растений преобладают многолетники, то среди земноводных статистически достоверное большинство составляют однолетники. Способность пройти на освободившемся участке поверхности водопокрытого или переувлажненного местообитания свой жизненный цикл вне зависимости от погодных условий конкретного года дает этим растениям дополнительные конкурентные преимущества на участках с переменным водным режимом.
Среди многолетних водных и земноводных растений нашей флоры вегетативно неподвижных видов всего два. Все остальные многолетники являются вегетативно высокоподвижными растениями за счет свободного перемещения особей по поверхности воды или в ее толще с ветрами и течениями, образованием перемещающихся в толще воды вегетативных диаспор (турионов) или захвата пространства вегетативными органами (ползучими побегами, столонами, длинными корневищами). Обилие длиннокорневищных, столонообразующих и ползучих растений среди истинно водных связанно с относительной гомогенностью как самой водной среды, так и водопокрытого грунта. В этих условиях конкурентно важнее оказывается захват нового участка, а не удержание ранее завоеванного.
Особо ценной в умеренных областях, где зимой природные водоемы покрываются льдом, оказывается способность водных растений образовывать турионы. Среди 48 видов водных многолетних растений природной флоры изученной территории этой способностью обладают лишь немногим менее половины — 22 вида. Способность к турионообразованию как в условиях умеренного гумидного, так и умеренного аридного дает обладающим ей водным растениям ряд конкурентных преимуществ.
Виды, относящиеся к разным жизненным формам, оказывают определенное предпочтение отдельным типам водных объектов, где та или иная адаптация дает им конкурентные преимущества над другими растениями.
Земноводные однолетники явно тяготеют к эфемерным водоемам, а водные однолетники избегают ривальных экосистем и обводненных болот. Их тяготение к эфемерным водоемам и связанным с ними нарушенным переувлажненным местообитаниям логично объясняется тем, что здесь при определенных погодных условиях конкретного года невозможно заранее предположить, в условиях какого фитоценоза, водного или наземного, растению придется проходить свой жизненный цикл. Способность в этих условиях пройти его в виде как водного, так и наземного растения, дает обладающим этим свойствам видам дополнительные конкурентные преимущества.
В условиях ривальных экосистем возможности для семян закрепиться в грунте, а затем и образовать побеги весьма ограниченны, особенно в умеренных широтах, где продолжительность вегетационного сезона в водных объектах редко превышает 60—80 дней. Потому характерных для таких экосистем однолетних видов в нашей флоре очень мало, и они представлены только встречающейся в Орловской и Рязанской областях термофильной Najas major.
Как и среди водных растений, среди многолетних земноводных и прибрежно-водных растений, обитающих на необсыхающих мелководьях, велика доля вегетативно высокоподвижных видов. Это связано также с относительной гомогенностью субстрата в этих местообитаниях и его хорошей проницаемостью корневищами и подземными столонами. Однако в этих экотопах в условиях поясного типа зарастания имеются участки с уже поделенным между разными особями пространством, где некоторые конкурентные преимущества начинают получать виды, вегетативно малоподвижные или вегетативно неподвижные, обладающие хорошей способностью удерживать за собой участок поверхности в месте произрастания. Все однолетники, встречающиеся на необсыхающих мелководьях, представлены водными формами земноводных растений, которые обычно обитают на подводных участках с нарушенным растительным покровом.
Полностью погруженные водные и земноводные растения тяготеют к природным водоемам (материковым и пойменным озерам), где прозрачность воды, как правило, выше, чем в водотоках или в искусственных водоемах. Данная группа в нашей флоре в основном представлена видами, имеющими отчетливое зональное тяготение: а) характерными для олиго- или дистрофных озер умеренных гумидных областей, требовательными к качеству воды (виды из рода Isoёtes, Caulinia tenuissima, Callitriche hermaphroditica L., Elatine hydropiper, E. triandra Schkuhr); б) характерными для мелководий водных объектов аридных областей теплолюбивыми Caulinia minor, Zannichellia palustris, Trapa natans.
Группа свободно плавающих растений, как полностью погруженных, так и плавающих на поверхности воды, напротив, относительно однородна и широко представлена почти во всех типах водоемов. Разнообразия в нее добавляют только попадающие в термальные водоемы из аквариумной или декоративной культуры Lemna minuta, Wolffia globosa и Pistia stratiotes.
Виды остальных групп особых приуроченностей к тем или иным группам водоемов не демонстрируют.