
Н. Е. Булыгин дендрология 2-е издание. Переработанное и дополненное допущено Государственным комитетом СССР по на- родному образованию в качестве учебник
| Вид материала | Учебник |
- В. М. Смолевского Издание 3-е, переработанное и дополненное Допущено Государственным, 5319.11kb.
- А. И. Соловьев введение в политологию Издание третье, переработанное и дополненное, 6711.57kb.
- А. И. Соловьев введение в политологию Издание третье, переработанное и дополненное, 6532.51kb.
- А. Г. Кучерена адвокатура второе издание, переработанное и дополненное Допущено Учебно-методическим, 12778.36kb.
- Учебник 3-е издание, переработанное и дополненное, 10138.23kb.
- А. Г. Асмолов психология личности принципы общепсихологического анализа Допущено Государственным, 4733.92kb.
- Учебник издание пятое, переработанное и дополненное проспект москва 2001 Том 3 удк, 11230.01kb.
- Учебник издание пятое, переработанное и дополненное проспект москва 2001 Том 3 удк, 11433.24kb.
- Учебник. 3-е издание, переработанное и дополненное, 10586.44kb.
- Наших дней, 5300.94kb.
ОСНОВЫ УЧЕНИЯ О РАСТИТЕЛЬНОМ ПОКРОВЕ
3.1. Ботанический вид и его ареал
Ботанический вид - это основная таксономическая единица в систематике растений, в их географии и экологии, в селекционной работе, при использовании растений в практических целях. Со-гласно академику В. Л. Комарову (1940) «... вид есть совокуп-ность поколений, происходящих от общего предка и под влиянием среды и борьбы за существование обособленных отбором от остального мира живых существ; вместе с тем вид есть опреде-ленный этап в процессе эволюции» *. Комплекс признаков и свойств, присущих виду, составляет его диагноз. К нему относятся жизненная форма, особенности морфологического и анатомиче-ского строения вегетативных и генеративных органов, кариотип (число и форма хромосом), биологические, экологические и дру-гие различия разных видов, современный естественный ареал.
Разные ботанические виды характеризуются различными ареалами, рассмотренными ниже.
Растения с широким ареалом - виды широкой экологической амплитуды, распространенные на огромных территориях одного или двух смежных материков, в пределах нескольких природных зон (сосна обыкновенная, береза повислая, ива козья).
Растения с узким ареалом - виды ограниченного распростра-нения на части материка со сравнительно однородным комплексом условий местопроизрастания. Если современный узкий ареал яв-ляется остаточным от некогда широкого ареала, сузившегося в результате сильных изменений внешней среды, его называют ре-ликтовым. Реликтовые ареалы присущи реликтовым видам (т. е. сохранившимся до настоящего времени с геологических эпох прошлого), однако не все реликтовые виды имеют реликтовые ареалы. Например, такие реликты мелового и третичного периодов, как черника, брусника, багульник, имеют довольно широкие со-временные ареалы. Следовательно, необходимо различать поня-
* Комаров В. Л. Учение о виде растений,- М.: АН СССР, 1940.- С. 212.
тия реликтовый ареал (всегда остаточный, узкий) и ареал ре-ликта, который может быть как узким, так и широким.
Эндемические растения, или эндемы - это виды обычно с уз-ким ареалом, приуроченным к какому-то определенному флори-стическому району, например, ель восточная и пихта Нордмана, образующие леса в западной части Кавказа; фисташка, растущая в предгорьях Средней Азии; кизильник блестящий - в Забай-калье и др. Эндемичными могут быть не только виды, но и роды, семейства, порядки растений.
Многие ботанические виды с эндемичными и реликтовыми ареалами относятся к категории редких и исчезающих и в СССР занесены в «Красную книгу».
Ареалы растений бывают трех типов: сплошные, разорванные и ленточные.
В сплошном ареале растения вида равномерно занимают все местообитания ареала (пихта сибирская, береза пушистая).
Ареал называется разорванным, когда территория, занятая видом, распадается на две или более обособленные части или помимо сплошной части ареала имеются островные местообитания этого же вида, удаленные на значительные расстояния (сосна обыкновенная и кедровая стланиковая, осина, береза повислая, дуб черешчатый, ольха черная).
Так как в различных частях разорванного ареала эволюция вида идет разными путями, то разрыв ареала может приводить к образованию новых видов, получивших название замещающих, или викарных (викарирующих). Так, викарными по отношению друг к другу являются дальневосточный орех маньчжурский и североамериканский орех серый; европейский клен остролистный и дальневосточный клен мелколистный.
К ленточным ареалам относят территории, занимаемые видом, вытянутые полосами по берегам рек или вдоль их древних русел (чозения, тополь черный, ива белая, ольха черная и бородатая). В ленточные могут переходить сплошные и разорванные ареалы на северных или южных их границах. Такие ареалы могут зани-мать дуб черешчатый в лесостепной и степной зонах, в подзоне южной тайги; ленточные боры сосны обыкновенной в Казахстане.
Ареалы растений изображают на географических картах тре-мя способами: очерчиванием границ ареала, сплошной его штри-ховкой и обозначением точками всех основных местообитаний растений вида. Последний способ наиболее информативен (см. приложение 3).
С детальной характеристикой ареалов древесных растений СССР рекомендуется знакомиться по справочным атласам С. Я. Соколова и др. «Ареалы деревьев и кустарников СССР» (1977-1986).
3.2. Внутривидовая изменчивость
и ее классификация у древесных растений
Любому ботаническому виду присуща в той или иной мере выраженная амплитуда варьирования признаков и свойств -формовое разнообразие. Так, деревья сосны обыкновенной могут различаться между собой габитусом кроны, энергией роста, дли-ной хвои, продолжительностью ее жизни, засухо- и зимостой-костью, смолопродуктивностью и т. д. У ели сибирской и колючей есть формы с серебристой, сизой и зеленой хвоей; у осины и дуба черешчатого - рано- и позднораспускающиеся фенологические формы. Все подобные примеры характеризуют присущий древес-ным растениям внутривидовой полиморфизм, обусловленный про-явлением разных форм внутривидовой изменчивости. Многообра-зие форм ее проявления у растений определило необходимость выделения соответствующих внутривидовых классификационных единиц (таксонов). Международным кодексом ботанической номенклатуры (1980) признаны следующие последовательно соподчиненные таксоны рангом ниже вида (species, сокращенно -sp.): подвид (subspecies, сокращенно - ssp.), разновидность (varietas, var.), подразновидность (subvarietas), форма (forma, f.), подформа (subforma). С учетом других современных клас-сификаций внутривидовых категорий у древесных растений (Прав-дин Л. Ф., 1967; Завадский К. М., 1968) ниже приводится краткая характеристика указанных таксонов.
Подвид, или географическая раса, экотип - наиболее крупная таксономическая единица внутри вида. Определяется как группа свободно скрещивающихся особей, характеризующихся одним или несколькими наследственными признаками и имеющая свой внутривидовой ареал. Как экотипы подвиды характеризуются разной экологической приспособленностью к определенным усло-виям местопроизрастания - климатическим, эдафическим, фито-ценотическим.
Подвиды имеются у многих древесных пород с широким ареа-лом: сосны обыкновенной, дуба черешчатого, берез повислой и карликовой.
Разновидность, или климатическая раса, климатический эко-тип (климатип) - таксон, выделяемый внутри вида или подвида. К одной разновидности относятся популяции (определение попу-ляции дано ниже), распространенные в регионе со сходными кли-матическими условиями. Чем разнообразнее климатические усло-вия в ареале вида, тем больше можно ожидать у него климатипов. Разновидности известны у сосны обыкновенной, ели европейской, дуба черешчатого. Имеют их и древесные породы с горно-высот-ными ареалами (пихта кавказская, орех грецкий).
Выделение разновидностей имеет большое значение для лесо-культурной практики, так как на их основе производят райониро-
вание заготовок семян древесных пород и создают лесные куль-туры. Очень важно учитывать экологические особенности клима-типов при интродукции, так как различные климатипы совершенно по-разному приспосабливаются к новым климатическим условиям за пределами естественного ареала.
Экологические особенности, специфика роста и развития под-видов и разновидностей являются наследственными.
Подразновидность, или эдафический тип (эдафотип), объеди-няет популяции в пределах границ разновидности (климатипа): например, нагорный, пойменный и солонцовый экотипы дуба че-решчатого, низинный и боровой экотипы ели европейской.
Помимо эдафотипов, к подразновидности иногда относят ценотипы - экотипы, связанные с определенными фитоценозами.
Модификационные (ненаследственные) формы растений, при-уроченные и приспособленные к определенным местообитаниям, называют экадами. Например, на верховых сфагновых болотах образуются болотные экады сосны обыкновенной, отличающейся карликовым или даже кустовидным ростом, укороченной хвоей, мелкими шишками. Однако после осушения болота болотная сосна резко увеличивает прирост и восстанавливает морфологические признаки, присущие лесным особям сосны.
Форма, или морфобиологическая группа - совокупность осо-бей вида, отличающихся от других особей того же вида по своим морфологическим, анатомическим признакам, биологическим или физиолого-биохимическим свойствам. Различают формы морфологические, биологические, фенологические, физиологиче-ские, биохимические, иммунологические.
Морфологические формы выделяют по одному или нескольким морфологическим признакам: пирамидальные и плакучие формы; широко- и узкокронные формы, различающиеся по конфигурации, размерам, окраске листьев, цветков, шишек, плодов, семян и т. д. Все морфологические формы имеют большое значение в декора-тивном садоводстве, но их важно учитывать и в практике лесной селекции.
Биологические формы отличаются по энергии роста, долговеч-ности, репродуктивной способности (быстрорастущие формы сосны, начинающие рано плодоносить формы ореха грецкого, устойчиво и обильно плодоносящие формы дуба).
Фенологические формы являются также биологическими, но различаются сроками прохождения фенофаз, продолжительно-стью роста побегов и вегетации, циклов формирования плодов и семян (рано- и позднораспускающиеся формы ели европейской, дуба черешчатого, ясеня обыкновенного; рано- и поздноцветущие формы липы мелколистной; формы с различными сроками созре-вания плодов у ореха грецкого, дуба черешчатого, черешни). В тех случаях, когда фенологические формы отражают экологи-ческие различия разных климатипов, их рассматривают как фено-
логические разновидности, или сезонные расы (рано- и поздно-распускающиеся формы дуба черешчатого).
Физиологические формы отличаются особенностями проявле-ния своих физиологических функций. Так, особи вяза мелколист-ного различаются по солевыносливости, ели европейской - по заморозкоустойчивости.
Биохимические формы объединяют группы особей, различаю-щихся содержанием химических веществ в их органах. Например, у сосны обыкновенной известны формы высоко- и низкосмоло-носные; у рябины, кизила, облепихи - формы с различным содер-жанием витаминов в плодах.
Иммунологические формы отличаются устойчивостью к болез-ням и насекомым-вредителям. У осины и других видов тополя, например, имеются формы, устойчивые и не устойчивые к ство-ловой гнили.
Формы могут быть ареальными и безареальными. В первом случае их распространение приурочено к определенным геогра-фическим районам и условиям местообитания, во втором они встречаются по всему ареалу без явной приуроченности к каким-либо конкретным районам и экологическим условиям.
Подформа, или биотип - наименьший внутривидовой таксон, объединяющий группы генетически одинаковых особей. У пере-крестноопыляемых растений, в том числе у древесных, фактически каждый индивид является биотипом, так как образуется в резуль-тате слияния генетически разнородных гамет. К подформе отно-сится и клон - вегетативное потомство одной особи. Клоны об-разуют все древесные растения, способные к вегетативному раз-множению (естественному и искусственному).
Среди внутривидовых подразделений особое место принадле-жит популяции - ареальной категории, таксономически зани-мающей промежуточное положение между экотипом (разновид-ностью, подразновидностью) и формой. Популяция - это группа свободно скрещивающихся или потенциально способных к скре-щиванию особей одного вида, в течение большого числа поколе-ний населяющих соответствующую территорию и обнаруживаю-щих определенные пространственно-временные взаимоотношения.
По современным представлениям, каждый биологический вид - это сложная система популяций, через последовательную цепь поколений которых осуществляется эволюционный процесс. Элементарным источником для начала этого процесса у растений служит их индивидуальная изменчивость, порождающая много-образие внутривидовых форм биотипов. Однако эволюционирует не особь, а популяция. Поэтому в системе внутривидовых таксонов популяция занимает центральное место как эволюционирующая единица.
В результате селекционной работы ученые создали и продол-жают создавать множество хозяйственно ценных сортов растений
(культиваров). Сорт - совокупность культивируемых растений, четко отличающихся морфологическими, физиологическими, ци-тологическими, биохимическими признаками и сохраняющих при воспроизведении (половом или бесполом) свои отличительные признаки. Сорта особенно широко распространены в практике плодоводства и декоративного садоводства. Латинские названия сортов указывают после видового, с прописной буквы, в кавычках. Например, ясень обыкновенный, сорт плакучий - Fraxinus excel-sior L. cv. 'Pendula', или F. excelsior L. 'Pendula'.
У лесоводов очень популярен термин «древесная порода». Но конкретного ботанического ранга он не имеет, так как в одних случаях под древесной породой понимают род, а в других - вид.
3.3. Интродукция древесных растений и ее значение
Под интродукцией растений понимают целенаправленную деятельность человека по введению в культуру новых видов, форм и сортов путем разведения их за пределами естественного ареала (виды, подвиды, разновидности) или продвижения в новые районы сортов. Интродуцированные растения называют интродуцентами, или экзотами, в отличие от местных видов, которые относят к аборигенным, или автохтонным.
Интродукция древесных растений включает распространение семян, черенков, а иногда и молодых растений целиком. Семенной способ разведения интродуцентов является более эффективным, так как обеспечивает лучшую адаптацию интродуцируемых дре-весных растений к новым условиям внешней среды. Во всех слу-чаях при интродукции человек имеет дело не с видом в целом (вид как система популяций существует только в своем естественном ареале), а лишь с определенными представителями его популяций. Интродукция растений своими корнями уходит в глубокую древ-ность, но с каждым десятилетием она осуществляется во все воз-растающих масштабах. Например, площади под лесными культу-рами хозяйственно ценных экзотов за рубежом уже превышают 50 млн га.
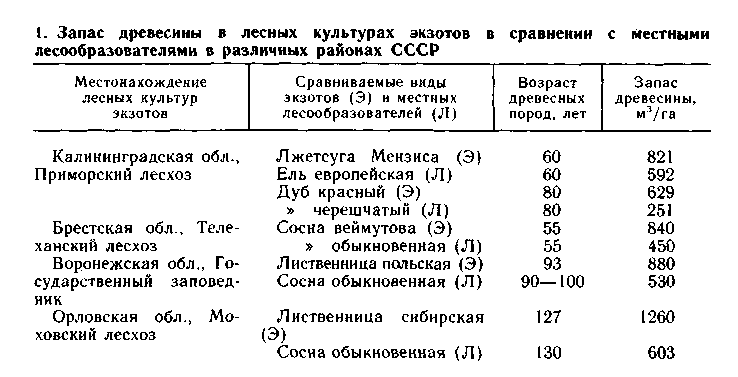
Для лесного хозяйства прежде всего важна интродукция таких лесообразователей, которые способны обеспечить значительное повышение производительности лесов и сокращение сроков вы-ращивания высококачественной древесины. В СССР имеется не-мало примеров успешного решения этой задачи, о чем свидетель-ствуют материалы табл. 1, составленной по данным публикаций П. И. Лапина и др. (1979), Г. И. Редько и Е. А. Федорова (1982). Но в специальной дендрологической литературе (Соколов С. Я. и др., 1949-1962; Гурский А. В., 1957; Мауринь А. М., 1970; Щепотьев Ф. Л., Павленко Ф. А., 1975; Калуцкий К. К., 1981; Ярославцев Г. Д., 1983 и др.) содержится много и других фактов исключительно высокой продуктивности лесных культур интроду-
цированных лесообразователей, конечно, в почвенно-климатиче-ских условиях, соответствующих их экологическим особенностям.
Например, разведение таких быстрорастущих и хозяйственно ценных древесных пород, как секвойя, секвойядендрон, метасек-войя, криптомерия, кедр гималайский, из-за их высокого тепло-любия и низкой зимостойкости возможно и практически целесо-образно преимущественно в отдельных районах советских суб-тропиков (Южный берег Крыма, Закавказье, оазисы южных районов Средней Азии) или в южной части Приморья Дальнего Востока. Совершенно уникальным памятником успешной интро-дукции таежных лесообразователей, проведенной еще по указу Петра I, является существующая и поныне в 70 км к северу от Ленинграда заповедная Линдуловская лиственничная роща (разновозрастные культуры лиственницы Сукачева, подробно охарактеризованные в работе Г. И. Редько, 1984).
Такие интродуценты, как гледичия, белая акация, айлант, вяз мелколистный, ясень ланцетолистный, разные виды и сорта топо-лей, в СССР имеют большое значение для практики агролесоме-лиорации, степного и полезащитного лесоразведения. Они ценятся за быстроту роста, засухоустойчивость и солевыносливость, спо-собность предотвращать водную и воздушную эрозию (разруше-ние) почвы, закреплять пески и склоны оврагов. Широкое распро-странение получили древесные экзоты при облесении карьеров, отвалов, терриконов.
Но особенно популярны интродуценты при озеленении. В город-ских насаждениях нашей страны суммарный состав древесных экзотов превышает 500 видов, а с учетом декоративных форм и сортов эту цифру практически можно удвоить. Почти всюду в озеленительных посадках интродуценты явно доминируют над древесными видами местной флоры (аборигенными видами).
Большое внимание к интродуцентам в практике озеленения обус-ловлено тем, что в условиях урбанизированной среды они во мно-гих случаях оказываются более устойчивыми и долговечными, чем местные виды; их использование обеспечивает существенное повышение эстетических и санитарно-гигиенических свойств озе-ленительных посадок, способствует сокращению затрат на выра-щивание посадочного материала, содержание городских зеленых насаждений.
Существует обширная дендрологическая литература по интро-дуцентам, применяемым в СССР для озеленения: шеститомное издание «Деревья и кустарники СССР» (1949-1962), дендроло-гические справочники А. А. Качалова (1970) и В. И. Некрасова (1986), «Декоративная дендрология» А. И. Колесникова (1974) и др. Не менее широко древесные экзоты применяют в сельском и в других отраслях народного хозяйства СССР. С различными направлениями их хозяйственного использования рекомендуется знакомиться по специальной ресурсоведческой литературе (Вульф Е. В., Малеева О. Ф., 1969; Приступа А. А., 1973).
Интродукция растений, особенно древесных, далеко не всегда завершается успехом. Удачной она оказывается лишь в тех слу-чаях, когда новые условия внешней среды в достаточно полной мере соответствуют биологическим особенностям и экологическим свойствам интродуцента. При отсутствии такого соответствия интродуцированные растения очень плохо приспосабливаются за пределами природного ареала, а нередко и гибнут, т. е. не ак-климатизируются.
Акклиматизация - это процесс приспособления растений к новым условиям среды за счет изменения исходного генотипа (изменения наследственных свойств). При акклиматизации раз-личают фенотипические (ненаследственные) изменения, проис-ходящие в онтогенезе растений на уровне особи, и изменения гено-типические (наследственные), которые реализуются только через цепь интродукционных популяций на основе жесткого естествен-ного и искусственного отбора. При интродукции растений разли-чают также натурализацию - перенесение растений в экологи-ческие условия, подобные или даже более благоприятные, чем в естественном ареале.
Введению в производственную культуру любого нового интро-дуцента всегда предшествует широкий комплекс длительных ис-следований, связанных с выбором исходного растительного мате-риала для интродукции, с организацией и проведением интродук-ционных испытаний растений, с изучением их реакции на воздей-ствие новых условий внешней среды, с разработкой агротехники, обеспечивающей наиболее успешное приспособление растений к этой среде. В СССР в настоящее время интродукционные испы-тания древесных растений проводят свыше 200 научно-производ-
ственных центров - ботанических и дендрологических садов, лесных и плодово-ягодных опытных станций и других учреждений. Более подробно с теоретическими основами и методами интро-дукции древесных растений, их акклиматизацией рекомендуется знакомиться по работам П. И. Лапина (1979) и В. И. Некрасова (1980).
3.4. Основные понятия лесной фитоценологии и биогеоценологии
Фитоценоз и растительная ассоциация. Любой участок суши, занятый растениями, не представляет собой случайного сочетания какого-либо числа видов и биотипов, живущих независимо друг от друга. Каждое растительное сообщество возникает и развивает-ся при определенных условиях внешней среды, в результате слож-ного взаимодействия между растениями сообщества и другими компонентами среды их обитания. Раздел ботаники изучающий закономерности формирования растительных сообществ, их вза-имодействия со средой, получил название фитоценологии. Основы этой науки заложил акад. В. Н. Сукачев.
Основным объектом изучения фитоценологии является фито-ценоз - конкретное растительное сообщество на определенной территории, характеризующееся своим составом, строением и взаимодействием между растениями, а также между ними и средой. Эти взаимодействия проявляются в различных направ-лениях.
Прежде всего в фитоценозе происходит конкуренция между разными видами и особями внутри вида за свет, воду, минеральные вещества, пространство. Эта конкуренция приводит к отмиранию огромного числа особей в период формирования сообщества, к угнетенности значительного числа видов и оказывает формирую-щее влияние на растения фитоценоза. Так, учет всходов березы на гари показал, что в двухлетнем возрасте их может быть до 3,5 млн шт. на 1 га, однако к возрасту спелого леса остается 400-600 деревьев на той же площади, остальные погибают. Оставшиеся и выросшие в лесу деревья имеют стройный, почти цилиндриче-ский ствол, узкую, высокоподнятую крону, тонкие ветви, характе-ризуются сравнительно слабым плодоношением. Березы, выросшие на свободе, имеют широкую низкоопущенную крону, разветвлен-ный на толстые сучья ствол, характеризуются более устойчивым и обильным цветением и плодоношением. Аналогичные примеры можно привести и в отношении других древесных пород.
Все виды деревьев, выросших на свободе, выполняют свою биологическую функцию - размножение лучше, чем деревья, выросшие в сообществе и испытывающие определенное угнетение со стороны других компонентов фитоценоза. В то же время в рас-тительном сообществе одни виды растений создают для других
благоприятные и даже необходимые условия жизни. Так, древес-ные растения создают благоприятные условия для лесных кустар-ников, трав, мхов, лишайников. Необходимые условия жизни создают автотрофные растения для гетеротрофных (сапрофитов и паразитов), а также растения-симбионты, например микотроф-ные грибы.
Так как в образовании фитоценоза принимают участие расте-ния разных видов и жизненных форм, обладающих различными экологическими особенностями, сообщество приобретает особую структуру в форме ярусности. Ярусность присуща любым фито-ценозам, но особенно ярко она выражена в лесу. Наиболее высокие деревья здесь составляют первый ярус; менее высокие - второй; кустарники подлеска - третий; кустарнички, травы, мхи и ли-шайники - четвертый и пятый. Временно растения могут нахо-диться в несвойственном им ярусе; например, всходы деревьев -в пятом, подрост - в пологе четвертого или третьего яруса.
Ярусы в фитоценозах неоднородны, имеют определенную структуру и состоят из частей, различающихся жизненными фор-мами растений, их видовым составом и экологическими свойст-вами. Такие структурные части фитоценоза получили название синузий. Например, в еловом лесу сплошные заросли на опреде-ленных площадях из черники, брусники или мхов являются сину-зиями четвертого и пятого ярусов. К синузиям в лесу относят и внеярусную растительность: лианы, мхи, водоросли на стволах.
При более детальном изучении структуры лесных фитоценозов выделяют парцеллы - территориально обособленные раститель-ные микрогруппировки, например сосново-ландышевая парцелла, осиновая парцелла в ельнике и т. д.
В годичном цикле развития фитоценоза всегда имеет место сезонная смена аспектов, связанная с неодновременностью про-хождения растениями различных видов фенологических фаз. В лесу, например, многие виды трав цветут и вегетируют до рас-пускания листьев у деревьев.
Ярусность, сезонная смена аспектов, синузийность приводят к более полному и равномерному использованию фитоценозом занимаемого пространства, позволяют существовать большому числу особей на одной и той же территории.
Растительное сообщество, действуя на среду территории, из-меняет ее, создает свою среду, отличную от среды территории, не занятой сообществом. В лесу, например, режим климатических факторов (температура, осадки, влажность воздуха, освещен-ность, ветер) будет иным, чем на открытом месте. Почва же вообще является результатом жизнедеятельности растительного сооб-щества. Поэтому фитоценоз не является простым, механическим соединением растений, где свойства их оказываются лишь суммой свойств отдельных растений. В сообществе возникает целый ряд новых свойств, присущих лишь ему, как определенному сочетанию
растений, находящихся в сложном взаимодействии между собой и средой обитания.
Растительное сообщество существенно влияет и на эволюцию составляющих его видов растений. Как всякое явление в природе, оно возникает, развивается, достигает своего полного развития, а затем сменяется или молодым подобным фитоценозом, или дру-гим по составу видов, структуре и среде. Это приводит к динамике фитоценозов, или изменяемости их во времени. Различают эндо-динамические и экзодинамические смены фитоценозов.
Оба типа смен являются результатом изменения среды обита-ния сообщества. Но в первом случае это происходит под давлением жизнедеятельности самого фитоценоза (например, из-за измене-ния сообществом эдафических условий или конкурентных взаимо-отношений между растениями), а во втором - по причинам, не связанным с его жизнедеятельностью (пожары, ветровалы, буре-ломы, вырубка леса, осушение болот и т. д.). Процессы смены фитоценозов получили название растительной сукцессии. Лесо-воды смену лесных фитоценозов обычно понимают как смену лес-ных пород (например, смену елового леса осиновым или бере-зовым) .
Совершенно тождественных фитоценозов нет, но весьма сход-ные фитоценозы можно обнаружить в различных местах и объеди-нить их в определенную группу. Фитоценозы, имеющие одинаковую структуру (одинаковое число ярусов), одинаковый видовой состав ярусов и занимающие однородную среду, понимают как тип фито-ценоза, или растительную ассоциацию. При установлении расти-тельной ассоциации основным показателем является видовой состав ярусов растительности. Но между видами, входящими в ассоциацию, очень важно выделять те, которые определяют структуру сообщества и обусловливают создание особой среды, присущей данному сообществу. Такие виды растений называют эдификаторами ассоциаций. В лесных ассоциациях их эдифика-торами являются преобладающие виды деревьев - лесообразо-вателей, в кустарниковых зарослях - виды кустарников, на бо-лотах - мхи, на лугах - отдельные виды трав.
Лесные ассоциации устанавливают по преобладающим дре-весным породам, по видовому составу и развитию других ярусов, по эдафическим условиям. Для примера рассмотрим дюны, порос-шие сосновым лесом. Вершины дюн обычно заняты сосняками с лишайниковым покровом, склоны и неглубокие впадины между дюнами - сосновым лесом с покровом из гипновых зеленых мхов, более глубокие понижения - сосняками с густым травяным по-кровом. Если же есть понижения с избыточным застойным увлаж-нением, то живой напочвенный покров в сосновом лесу форми-руют сфагновые мхи, образующие торф. Хотя участки леса (лесные фитоценозы) на вершинах дюн не будут строго одинаковыми (одни отличаются возрастом деревьев, другие - их густотой или
густотой и составом лишайникового покрова и др.), все же фито-ценозы, занимающие вершины дюн, по своим основным признакам (структуре, составу ярусов, среде обитания) однородны в такой степени, что могут быть объединены в одну растительную, в дан-ном случае лесную, ассоциацию. То же можно сказать и о других лесных фитоценозах - о мшистых, травяных или сфагновых сосняках.
Таким образом, растительную ассоциацию можно определить как совокупность фитоценозов, однородных по взаимоотноше-ниям между видами растений, образующими их в соответствии с условиями среды, однородных по структуре, видовому составу ярусов и занимающих условия местопроизрастания с однородным комплексом экологических факторов.
Растительная ассоциация является основной систематической единицей растительности.
Подобно виду в систематике растений, каждой растительной ассоциации также дается определенное название, обычно двойное. Первое слово названия - родовое, соответствует названию эди-фикатора ассоциации; второе (видовое) чаще всего дают по на-званию характерного растения другого яруса, травяного или мохового покрова или по названиям растений-индикаторов. На-ряду с русскими названиями, в научных работах приводят и ла-тинские названия, например сосняк лишайниковый - Pinetum cladinosum; кедровник долгомошниковый - Cembretum poly-trichosum; ельник черничный - Piceetum myrtillosum; листвяг кисличный - Laricetum oxalidosum и т. д.
Для лесоводов лесные ассоциации важны тем, что участки леса, относящиеся к одной и той же ассоциации, характеризуются сходством всех лесоводственных особенностей (видового состава древостоя, его производительности и других таксационных эле-ментов, быстроты изреживания, хода естественного возобновле-ния и т. д.). Однако в своей практической деятельности лесоводы оперируют не лесными ассоциациями, а типами леса - понятием более широким, чем растительная ассоциация. Согласно учению акад. В. Н. Сукачева (1964) о биогеоценозе в лесоведении тип леса рассматривают как тип лесного биогеоценоза, включающего в себя все компоненты биоценоза и экотопа.