
Физиологическая цена адаптаций к подземному образу жизни: обыкновенная слепушонка ( ellobius talpinus pall.) В сравнении с наземными грызунами
| Вид материала | Автореферат диссертации |
- Председатель, 417.14kb.
- Адаптации животных к нормальным и экстремальным факторам среды, 44.24kb.
- Рабочая программа дисциплины «Адаптивная изменчивость организмов» Код дисциплины, 90.26kb.
- Понятие адаптации, 125.13kb.
- Паразитология балашов Ю. С. Паразито-хозяинные отношения членистоногих с наземными, 282.93kb.
- Литература. Тема урока: «Очерк жизни и творчества И. А. Гончарова. Три романа- «Обыкновенная, 18.56kb.
- Программа «Формирование активной жизненной позиции и стремления к здоровому образу, 78.18kb.
- Сорта яблонь антоновка обыкновенная (зимний), 137.97kb.
- Задачи: дать понятие о роли здоровья в жизни человека; прививать детям стремление, 75.46kb.
- «Приветствие», 177.29kb.
На правах рукописи
Новиков Евгений Анатольевич
ФИЗИОЛОГИЧЕСКАЯ ЦЕНА АДАПТАЦИЙ К ПОДЗЕМНОМУ ОБРАЗУ ЖИЗНИ: ОБЫКНОВЕННАЯ СЛЕПУШОНКА (ELLOBIUS TALPINUS PALL.) В СРАВНЕНИИ С НАЗЕМНЫМИ ГРЫЗУНАМИ
Специальность 03.00.08 - зоология
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
НОВОСИБИРСК – 2008
Работа выполнена в лаборатории физиологических адаптаций позвоночных животных Института систематики и экологии животных Сибирского отделения Российской академии наук
Научный консультант: доктор биологических наук, профессор
М. П. Мошкин
Официальные оппоненты: доктор биологических наук К.А. Роговин
ИПЭЭ РАН
доктор биологических наук Л.В. Осадчук
ИЦиГ СО РАН
доктор биологических наук В.Д. Гуляев
ИСиЭЖ СО РАН
Ведущая организация: Институт экологии растений и животных УРО РАН
Защита диссертации состоится 9 декабря 2008 года в 10 часов на заседании диссертационного совета Д 003.033.01 в Институте систематики и экологии животных СО РАН по адресу: Новосибирск, 630091, ул. Фрунзе, 11. Факс (383)2170973, e-mail: dis@eco.nsc.ru
С диссертацией можно ознакомиться в библиотеке Института систематики и экологии животных СО РАН
Автореферат разослан ____________ 2008 года.
Ученый секретарь диссертационного совета,
кандидат биологических наук Л.В. Петрожицкая
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы: Возможность функционирования организма в определенном диапазоне значений экологических факторов определяется комплексом адаптаций, затрагивающих различные стороны его жизнедеятельности (Пианка, 1982). Однако любая адаптация в процессе своего формирования требует привлечения дополнительных энергетических и пластических ресурсов и, следовательно, имеет определенную физиологическую цену (Меерсон, Пшенникова, 1988). Ограниченность внутренних ресурсов организма неизбежно приводит к тому, что между различными его системами возникает конкуренция, результатом которой является достижение компромисса жизненных интересов (van Noordwijk, de Jong, 1986; Stearns, 1992; Ricklefs, Wikelski, 2002). В последнее десятилетие ведется активный поиск таких компромиссов, сформировавший новое направление эволюционной экологии (Stearns, 2000; Ricklefs, Wikelski, 2002). Однако большинство работ этого направления посвящено рассмотрению частных взаимоотношений между отдельными системами организма, тогда как для понимания механизмов, лежащих в основе формирования адаптационных стратегий, необходимо комплексное изучение всех жизненно-важных функций и их взаимосвязей. Эта задача технически представляется трудновыполнимой, однако методические сложности можно, по крайней мере, частично, устранить использованием сравнительно-экологического подхода. Действительно, для выяснения меры участия какой-либо функции в формировании адаптационной стратегии нам важны не столько абсолютные значения характеризующих эту функцию физиологических параметров, сколько диапазон и направление их изменчивости в градиенте экологических факторов.
Таким образом, на современном этапе развития экологии назрела необходимость выполнения работы по комплексному анализу ресурсной емкости основных жизненно-важных функций у видов различной экологической специализации. Однако, до сих пор сравнительный анализ функциональной активности систем организма с позиций их физиологической стоимости и ресурсоемкости у животных, относящихся к различным биологическим типам, не проводился. Поскольку данная задача в полной мере не выполнима силами одной исследовательской группы, мы вынуждены были сконцентрировать внимание на одном модельном виде с меньшей глубиной проработки остальных объектов. Анализ имеющихся в литературе данных об основных физиологических механизмах формирования адаптационной стратегии, которой следует изучаемый вид, позволил определить набор параметров, наиболее важных в контексте исследования.
В качестве основного (модельного) объекта исследования нами была выбрана обыкновенная слепушонка ( Ellobius talpinus Pall.) - вид грызунов, экологически специализированный к обитанию в подземной среде. Объектами сравнительного анализа послужили симпатричные, таксономически- и размерно-близкие виды наземных грызунов, относящиеся к биологическим типам хомяков, полевок и мышей, т.е. имеющих различную экологическую специализацию.
Цель работы: на примере специализированного подземного вида, обыкновенной слепушонки, исследовать особенности распределения ресурсов организма на различные стороны жизнедеятельности как физиологическую основу формирования адаптационной стратегии.
Задачи исследования:
- Сопоставить уровни функциональной активности различных систем организма: терморегуляторной, репродуктивной, иммунной и эндокринной у обыкновенной слепушонки и наземных видов грызунов.
- У обыкновенной слепушонки проанализировать сезонную и эко-географическую изменчивость рассматриваемых показателей.
Положения, выносимые на защиту:
- Обитание в стабильной подземной среде позволяет обыкновенной слепушонке минимизировать затраты на поддержание жизнедеятельности, развитие, размножение, терморегуляцию, иммунитет. Уменьшаются и мобилизационные возможности организма при действии стрессоров физической и популяционной природы. Ограничение функциональной активности жизненно важных систем организма высвобождает ресурсы, необходимые для энергетического обеспечения роющей деятельности.
- Сезонные особенности биологии обыкновенной слепушонки, выявленные в северной части видового ареала (активация терморегуляторной функции в холодное время года и подснежное размножение), определяются состоянием кормовой базы. Увеличение доступности пищи в осенне-весенний период снижает затраты на роющую деятельность и обеспечивает ресурсами как терморегуляторную, так и репродуктивную функции организма.
- В популяциях обыкновенной слепушонки, занимающих северо-восточную периферию видового ареала, при сохранении типичных для вида индивидуальных показателей плодовитости увеличивается доля размножающихся самок. Таким образом, повышенная смертность в экологически пессимальных условиях компенсируется увеличением репродуктивного выхода на уровне популяции.
Научная новизна: Анализ комплекса физиологических параметров позволил установить, что в формирование энергосберегающего бюджета подземных грызунов вовлечены практически все системы организма. Впервые на фактическом материале показано, что уровень достижения компромиссов между иммунитетом и другими жизненно-важными функциями организма, терморегуляторной и репродуктивной, определяется экологической специализацией вида. Впервые установлено наличие сопряженной изменчивости эндокринной и метаболической реакции на стресс на межвидовом уровне. Сформулировано представление о роли стресса в модификации видовых онтогенетических программ. Анализ географической изменчивости физиологических параметров рассматриваемых видов показал, что более высокая стоимость жизнедеятельности в экологически пессимальных условиях приводит к снижению резервных возможностей организма. Впервые на основе многомесячной регистрации температуры тела обыкновенной слепушонки, зимующей в естественных условиях, получены прямые доказательства того, что зимняя спячка и торпор, как способы ресурсной экономии, не свойственны данному виду животных.
Практическая значимость: Результаты работы демонстрируют возможность использования обыкновенной слепушонки в качестве удобного модельного объекта для биомедицинских исследований, в частности, как вида с крайне низкой реактивностью механизмов специфического иммунитета, стресс-реакции и термогенеза. Выявление физиологических механизмов, обеспечивающих высокую продолжительность жизни подземных грызунов, может оказаться ценным вкладом в геронтологические исследования.
Изучение уровня функциональной активности иммунной системы и ее связи с социальным и физиологическим статусом животного в естественном и антропогенном паразитарном окружении является важным шагом в понимании механизмов циркуляции возбудителей ряда заболеваний, в том числе и особо опасных трансмиссивных инфекций, которое необходимо для построения научно обоснованных прогнозов и планирования профилактических мероприятий.
Результаты работы используются при чтении курса лекций по экологической физиологии в Новосибирском, Томском и Кемеровском университетах.
Апробация работы: Материалы диссертационной работы были представлены на IV-й (Польша, 1993 г.), V-й (Марокко, 1995 г.), VI-й (Израиль, 1998 г.) и XI-й (Россия, 2008 г.) международных конференциях “Rodens & Spatium”, на XXV-й международной этологической конференциии (Австрия, 1997 г.), Международном симпозиуме по новым направлениям науки о животных в новом веке (Япония, 2001 г.), на IX международном териологическом конгрессе (Япония, 2005 г.), на 3 и 4 съездах физиологов Сибири и Дальнего Востока (Новосибирск, 1997, 2002 г.г.), на Сибирской зоологической конференции (Новосибирск, 2004 г.), на III Международной конференции "Актуальные проблемы экологии" (Караганда, 2004 г.), конференциях «Поведение и поведенческая экология млекопитающих» (Черноголовка 2005 г.), «Популяционная экология» (Томск, 2006 г.), VIII популяционном семинаре «Популяции в пространстве и во времени» (Нижний Новгород, 2005 г.). Основные результаты работы были представлены в виде пленарного доклада на VIII съезде Всероссийского териологического общества (Москва, 2007 г.) и пленарной лекции на Всероссийской конференции молодых ученых «От Арктики до Антарктики» (Екатеринбург, 2007 г.).
Публикации: По материалам исследований опубликовано 44 печатных работы, из них 7 статей в рецензируемых отечественных журналах, рекомендованных ВАК для опубликования основных научных результатов диссертаций; 7 статей в рецензируемых зарубежных журналах, в т.ч. 4 – в журналах, включенных в международную систему цитирования “Web of science”; глава в коллективной монографии, выпущенной в издательстве “Springer ”и 5 статей в отечественных и зарубежных сборниках.
Структура и объем работы: Диссертация состоит из введения, 7 глав, заключения, выводов и библиографического списка, содержит 50 рисунков и 20 таблиц, изложена на 393 страницах машинописного текста. Список цитируемой литературы состоит из 834 источников, в т.ч. 619 иностранных.
Благодарности. Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (гранты 96-04-50113; 99-04-49927; 00-04-49210; 02-04-49253; 03-04-48768; 07-04-00871), фонда Сороса (гранты RAH-000 и RAH-300), интеграционных проектов СО РАН, гранта НШ-1038.2003.4, программы президиума РАН «Происхождение и эволюция биосферы» (Гос. контракт № 151). Неоценимую помощь в сборе и анализе материалов для работы автору оказали друзья и коллеги - сотрудники лаборатории физиологических адаптаций позвоночных животных ИСиЭЖ СО РАН: Д.В. Петровский, И.Е. Колосова, Е.Ю. Кондратюк, В.В Мак, В.В. Панов, Е.А. Литвинова, А.Е. Гармс, А.Е. Акулов, а также - ныне покойный сотрудник ИСиЭЖ СО РАН А.К. Добротворский, сотрудники Института экологии животных и растений УРО РАН Н.Г. Евдокимов и Н.В. Синева, студенты Новосибирского, Кемеровского и Томского университетов, учащиеся МСОШ «Диалог». Всем им автор выражает свою горячую благодарность. Автор благодарит А.В. Сурова, А.Ю Телицину, О.В. Осипову – за любезно предоставленных лабораторных животных, И.Ю. Баклушинскую и Л.В. Якименко – за помощь в видовом определении слепушонок, А.В. Осадчука, В.И. Евсикова, В.В. Глупова, Ю.Н. Литвинова и А.А. Никольского за труд по ознакомлению с работой и полезные замечания. Автор выражает благодарность своим Учителям и, в первую очередь, научному консультанту работы - доктору биологических наук, профессору М.П. Мошкину а также – профессорам И.В. Стебаеву, В.И. Евсикову и ныне покойному Б.С. Юдину. Автор благодарит всех коллег, поименное перечисление которых, к сожалению, не возможно в формате автореферата, поддержавших его словом и делом при подготовке диссертации.
ФИЗИОЛОГИЧЕСКАЯ ЦЕНА АДАПТАЦИЙ
Энергия, поступающая в организм с пищей, расходуется на обеспечение жизнедеятельности (основной обмен и затраты на добывание пищи), поддержание гомеостаза (терморегуляция и иммунитет), соматическую и генеративную продукцию (Wade, Schneider, 1992; Klein, Nelson, 1999). В соответствии с современными эволюционно-экологическими представлениями, оптимальное распределение ресурсов между функциями, обеспечивающими выживание и репродуктивный успех особей, является основой видовых адаптационных стратегий (Stearns, 1992). Однако в природе поступление энергии в организм крайне редко находится в точном соответствии с его потребностями. В случае, если суммарные затраты организма превышают количество доступной энергии, имеет место компромиссное ограничение активности отдельных функций, направленное на снижение общей потребности в ресурсах. При незначительном снижении поступления энергии снижаются затраты на локомоцию, рост и размножение, т.е. функции, не оказывающие прямого влияния на выживаемость особи. При остром дефиците энергии снижается ресурсное обеспечение терморегуляции и иммунитета, что может привести к снижению жизнеспособности (Wade, Schneider, 1992; Klein, Nelson, 1999). Перераспределение внутренних ресурсов организма в соответствии с меняющейся иерархией потребностей обеспечивается комплексом физиологических механизмов, среди которых важное место занимает стресс-реакция (Меерсон, 1981; Sapolsky et al., 2000).
Анализируя патогенные эффекты стресса, Ф. Меерсон (1981) ввел понятие физиологической цены адаптации, которая определяется тем, что при адаптации к чрезмерным для данного организма нагрузкам все приспособительные реакции организма имеют лишь относительную целесообразность. Цена адаптации проявляется в прямом изнашивании функциональной системы, на которую падает главная нагрузка и в т.н. отрицательных перекрестных эффектах – нарушениях функционирования систем, непосредственно не связанных с нагрузкой (Меерсон, Пшенникова, 1988). Очевидно, что данная концепция применима и для более обычных в природе ситуаций, когда снижение функциональной активности какой либо системы как следствие перераспределения ресурсов в процессе формирования адаптации не носит характера явной патологии.
Поскольку перераспределение ресурсов между функциями имеет место и при освоении животными нового экологического пространства, сопутствующие изменения функциональной активности жизненно важных систем организма можно рассматривать как физиологическую цену адаптаций популяционного и видового уровня. Данная работа посвящена выявлению закономерностей, определяющих изменчивость функциональной активности систем организма, обусловленную экологической специализацией вида и пространственно-временной неоднородностью параметров среды его обитания. В качестве примера будут рассмотрены адаптации млекопитающих к подземному образу жизни.
ОСОБЕННОСТИ БИОЛОГИИ ПОДЗЕМНЫХ ГРЫЗУНОВ
Эволюционная тенденция перехода к подземному образу жизни, характерная для животных аридной и семиаридной зон, связана с обитанием в открытых биотопах, что требует от животных строительства сложных подземных убежищ (Наумов, 1948). Кроме того, по мере аридизации ландшафтов снижается доля надземной фитомассы и уменьшается ее доступность для фитофагов. Для большинства подземных млекопитающих характерны общие морфологические и физиологические адаптации, облегчающие передвижение в почве (Орлов, 1978; Ar, 1987), которое требует в 360-3400 раз больше энергии, чем перемещение на то же расстояние по поверхности (Vleck, 1979). Во время рытья животные потребляют количество кислорода в 2,8-7,2 превышающее базальный уровень метаболизма (Vleck 1979). Вместе с тем интенсивность аэробного обмена почвенных животных ограничивается низким парциальным давлением кислорода и повышенным содержанием углекислого газа (Ar, 1987). Адаптация к этим условиям достигается за счет снижения уровня основного обмена (в 1,5–2 раза по сравнению с надземными видами такого же размера) и затрат на активную терморегуляцию (Kleiber 1975; McNab, 1979; Withers, Jarvis, 1980).
Отличительной особенностью почвы как среды обитания являются ее хорошие защитные свойства. Специфика среды обитания исключает почвенных животных из большинства паразито-хозяинных систем, в которые вовлечены наземные виды, а высокая степень изоляции особей (или отдельных внутрипопуляционных группировок) ограничивает поток паразитов на уровне популяции (Loehlie, 1995). Относительно низкая смертность в сочетании с ограниченными возможностями расселения определяет низкую плодовитость большинства подземных животных (Наумов, 1948) и стимулирует переход к семейно-групповому и даже (у грызунов сем. землекоповых) к эусоциальному образу жизни (Lovergrove, 1991). Как у общественных перепончатокрылых, в колониях эусоциальных землекоповых в размножении участвует только одна самка и несколько самцов. Неполовозрелые особи (не являющиеся, в отличие от перепончатокрылых, стерильными) выполняют функции «рабочих», «солдат» и заботятся о потомстве (Jarvis, 1981; Jarvis, Bennet, 1993). Репродуктивно активные особи отличаются большими размерами (O’Riain et al., 2000) и большей продолжительностью жизни (Damman, Burda, 2006).
Семейно-групповой образ жизни характерен и для слепушонок (р. Ellobius), широко распространенных в аридных и семи-аридных ландшафтах Евразии (Громов, Ербаева, 1995). Различным сторонам их биологии посвящено достаточно большое количество работ (Наумов, 1948; Сластенина, 1963; Якименко, 1984; Якименко, Ляпунова, 1987; Евдокимов, 2001; Шевлюк, Елина, 2008 и др.). Результаты этих исследований, использованные нами для сравнительного анализа, будут рассмотрены в соответствующих разделах. Однако физиологические адаптации слепушонок, обусловленные обитанием в подземной среде, изучены достаточно слабо. До сих пор они рассматривались, главным образом, в связи с географической изменчивостью частот окрасочных фенотипов, выявленной у обыкновенной слепушонки в северной части ареала (Большаков и др., 1982; 1989; Мошкин и др., 1991).
МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗОВАННЫЕ В РАБОТЕ
В работе использованы материалы, собранные автором в период с 1993 по 2007 годы. Отловы обыкновенной слепушонки проводили в Искитимском и Сузунском районах Новосибирской области, в Угловском, Локтевском и Тюменцевском районах Алтайского края. Всего было отловлено и протестировано 1180 особей обыкновенной слепушонки. Для сравнительного анализа использованы данные, полученные автором и его коллегами в 1984-1989 и в 2001-2007 г.г. при изучении красной полевки из популяции, обитающей в Прителецкой тайге (респ. Алтай, Турочакский р-н), рыжей полевки и мохноногих хомячков из лабораторных популяций, поддерживаемых в ИПЭЭ РАН и любезно предоставленных нам О.В.Осиповой, А.В.Суровым и А.Ю.Телициной, а в ряде случаев – и других видов грызунов (восточно-азиатская лесная и полевая мыши, красно-серая и водяная полевки). Животных, используемых в тестах, содержали в стандартных лабораторных условиях при комфортной температуре и естественном режиме освещенности: Слепушонок - группами по 4 особи, животных остальных видов – поодиночке. Пищу, воду и гнездовой материал давали ad libitum.
Измерение стандартного и максимального обмена проводили методом непрямой калориметрии в установке закрытого типа (Калабухов, 1956). В качестве тестирующего охлаждения для оценки максимального потребления кислорода и эндокринно-метаболической реакции на холодовой стресс использовали 16 минутное охлаждение в гелиево-кислородной смеси при 7-8ºС (Rosenmann, Morisson, 1974). По отношению максимального обмена к стандартному рассчитывали метаболический индекс. Для оценки текущей интенсивности метаболизма измеряли потребление кислорода или выделение углекислого газа проточным методом с использованием газоанализаторов ПГА-12 и ПЭМ 2М. При использовании углекислотного датчика дыхательный коэффициент принимали равным 0,95. Показатели энергообмена выражали в общепринятых единицах. Двигательную активность оценивали с помощью установки, состоящей из инфракрасных датчиков охранной сигнализации и компьютерного интерфейса. Ректальную температуру тела измеряли с помощью термисторного датчика. Для изучения терморегуляторных адаптаций слепушонки в зимний период использовали сконструированные Д.В. Петровским накопительные термодатчики, способные в течение 22 месяцев с интервалом в 1 час фиксировать температуру тела особи. Осенью 2006 г. отловленным в природе зверькам имплантировали датчики и выпустили их в точке олова. Весной 2007 г. зверьков отловили повторно, извлекли датчики и сняли с них показания.
Уровень агрессивности и величину адренокортикальной реакции на социальный конфликт оценивали в тесте попарного ссаживания на нейтральной территории (Krebs, 1970; Colvin, 1973). Содержание белка в моче определяли по методике Бредфорда с красителем Кумасси (Досон и др., 1991). Величину гуморального иммунного ответа на эритроциты барана оценивали по числу антителообразующих клеток (АОК) в селезенке в пересчете на 1 г. массы тела (Сunningham 1965). Анализ концентраций стероидных гормонов в крови проводили флюориметрическим (Юденфренд, 1965) и радиоиммунным (Yalow, Berson, 1960) методами. Поскольку у грызунов, характеризующихся преобладанием кортикостерона, базальный уровень глюкокортикоидов был значительно выше, чем у видов с преимущественной секрецией кортизола, сравнение адренокортикального ответа на стрессирующие воздействия проводили на основе глюкокортикоидного индекса - отношения концентрации гормона в условиях стресса к его базальному уровню.
Видовую принадлежность слепушонок определяли по строению коренных зубов (Громов, Ербаева, 1995; Павлинов и др., 2002). Цитогенетические исследования отловленных нами животных из Новосибирской области, проведенные сотрудниками Института цитологии и генетики СО РАН, подтвердили их принадлежность к виду Ellobius talpinus c хромосомным набором 2N=NF=54 (Romanenko et al., 2007).
Обработку и анализ результатов проводили, в зависимости от свойств исследуемых выборок, методами параметрической или непараметрической статистики.
ЗАТРАТЫ НА ПОДДЕРЖАНИЕ ЖИЗНЕДЕЯТЕЛЬНОСТИ
Непрерывная регистрация параметров жизнедеятельности у джунгарских хомячков и обыкновенных слепушонок, содержащихся в стандартных условиях, показала, что при сопоставимом уровне активности и среднесуточного потребления кислорода слепушонка затрачивает значительно больше энергии на каждый двигательный акт (табл.1), что связано, очевидно, с видовыми особенностями локомоторного аппарата.
Таблица 1.
Показатели, отражающие энергетическую стоимость двигательной активности у обыкновенной слепушонки и джунгарского хомячка: уровень активности (секунд в час), среднесуточная интенсивность метаболизма (АDMR; млCO2* г-1*час-1), обмен покоя (мл CO2*г-1*час-1) и энергетическая цена активности (Дж*г-1*сек-1).
| | Обыкновенная слепушонка | Джунгарский хомячок | Достоверность различий (t-крит.) |
| Объем выборок | 8 | 12 | |
| Активность | 145,0±12,6 | 139,0±17,9 | н/д |
| ADMR | 4,35±0,09 | 4,18±0,05 | н/д |
| Обмен покоя | 2,46±0,04 | 3,53±0,02 | t=6,8; P<0,001 |
| Цена активности | 1,36±0,15 | 0,87±0,05 | t=2,7; P<0,01 |
Измерение метаболических и терморегуляторных характеристик мышевидных грызунов, проводившееся в летний период, показало, что среди протестированных нами видов слепушонка имеет минимальные значения стандартного и максимального обмена и обладает наименьшей способностью к поддержанию температурного гомеостаза (табл. 2.).
Таблица 2.
Величины стандартного и максимального обмена (мл*г-1*ч-1), метаболического индекса и падения температуры (ΔТ°С) после холодовой экспозиции у обыкновенной слепушонки и наземных видов грызунов.
| | Обыкновенная слепушонка | Рыжая полевка | Джунгарский хомячок |
| Объем выборок | 28 | 14 | 8 |
| Стандартный обмен | 1,6 0,1a | 3,9 0,3b | 3,0 0,2b |
| Максимальный обмен | 4,6 0,3 a | 12,0 0,5 b | 10,7 0,6 b |
| Метаболический индекс | 3,0 0,4 a | 3,2 0,3 a | 3,6 0,2 a |
| Падение температуры | -16,0 0,7 a | -13,3 1,0a | -6,7 1,3 b |
Примечания: Здесь и в табл.3 разными буквами отмечены значения одного и того же параметра, достоверно (критерий Тьюки; P<0,05) различающиеся между собой у разных видов.
Содержание свободных жирных кислот в крови, как до- так и после холодовой экспозиции у обыкновенной слепушонки оказалось достоверно меньшим, чем у наземных грызунов (критерий Тьюки; P<0,05). По содержанию глюкозы в крови протестированных животных такие различия отсутствовали (табл.3). Поскольку окисление жира дает меньший выход АТФ в пересчете на 1 моль потребленного кислорода, можно предполагать, что преимущественное использование в качестве субстрата углеводов является специфической адаптацией к условиям гипоксии.
Таблица 3.
Содержание основных биоэнергетических субстратов – глюкозы (ммоль*л-1) и свободных жирных кислот (СЖК; мкмоль*л-1) у слепушонки и наземных видов в покое и после холодовой экспозиции.
| | Обыкновенная слепушонка | Рыжая полевка | Джунгарский хомячок |
| Объем выборок | 17 | 10 | 8 |
| Глюкоза фон | 2,82±0,21 | 3,30±0,36 | 4,01±0,17 |
| Глюкоза холод | 6,97±0,44 | 5,98±0,66 | 7,80±0,80 |
| СЖК фон | 321,6±34,1а | 494,3±46,1b | 524,0±87,9 a,b |
| СЖК холод | 543,6±33,2 a | 918,5±102,1b | 1322,0±266 b |
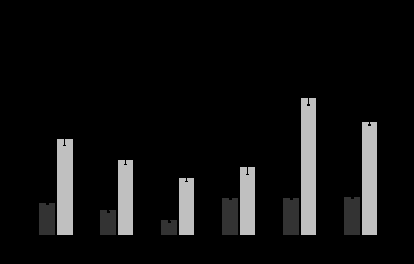 Рис. 1. Сезонные особенности метаболической и терморегуляторной реакции на острое охлаждение у обыкновенной слепушонки. Темные столбики – стандартный обмен, светлые столбики – максимальный обмен, Черные круги – падение температуры тела после холодовой экспозиции. Значения одного и того же параметра, достоверно (критерий Тьюки; P<0,05) различающиеся между собой в разные месяцы, помечены разными буквами.
Рис. 1. Сезонные особенности метаболической и терморегуляторной реакции на острое охлаждение у обыкновенной слепушонки. Темные столбики – стандартный обмен, светлые столбики – максимальный обмен, Черные круги – падение температуры тела после холодовой экспозиции. Значения одного и того же параметра, достоверно (критерий Тьюки; P<0,05) различающиеся между собой в разные месяцы, помечены разными буквами.Данные, полученные совместно с Д.В.Петровским, показывают, что в летний период у слепушонки снижается способность к поддержанию постоянной температуры тела и при умеренном охлаждении. Калоригенная реакция на норадреналин, отчетливо выраженная в холодное время года, летом практически отсутствует. Амплитуда спонтанных колебаний температуры тела составляет летом более 8°С, тогда как осенью – не превышает 4°С (Мошкин и др., 1999; Novikov et al., 1996; Moshkin et al., 2001).
Интенсивность метаболизма слепушонки в летние месяцы оказывается значительно ниже ожидаемой для грызуна такой массы тела исходя из линии регрессии, построенной нами на основании собственных результатов, полученных на разных видах с использованием единого методического подхода (рис.2). В холодное время года обыкновенная слепушонка по своим метаболическим и терморегуляторным возможностям не отличается (с учетом различий в массе тела) от наземных видов бореальных грызунов. В октябре величина максимального обмена у обыкновенной слепушонки оказывается даже выше, чем ожидаемая исходя из полученной нами эмпирической зависимости (рис.2).
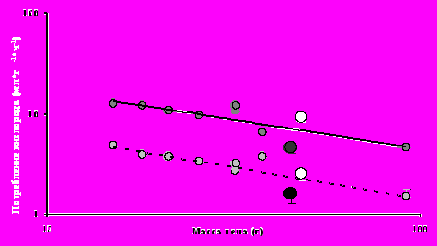
 Рис. 2. Зависимость средних для вида величин стандартного (светло-серые кружки, пунктирная линия) и максимального (темно-серые кружки, сплошная линия) обмена от массы тела для 9 видов мышевидных грызунов (мохноногие хомячки, лесные полевки, лесная и лабораторная мыши, обыкновенная слепушонка, водяная полевка), представленная в логарифмических координатах. Для обыкновенной слепушонки приведены летние (обозначены черным цветом) и осенние (белым цветом) значения показателей. За исключением отмеченных случаев стандартные ошибки средних значений не выходят за пределы точек-маркеров.
Рис. 2. Зависимость средних для вида величин стандартного (светло-серые кружки, пунктирная линия) и максимального (темно-серые кружки, сплошная линия) обмена от массы тела для 9 видов мышевидных грызунов (мохноногие хомячки, лесные полевки, лесная и лабораторная мыши, обыкновенная слепушонка, водяная полевка), представленная в логарифмических координатах. Для обыкновенной слепушонки приведены летние (обозначены черным цветом) и осенние (белым цветом) значения показателей. За исключением отмеченных случаев стандартные ошибки средних значений не выходят за пределы точек-маркеров.Вопрос о зимних терморегуляторных адаптациях вида до сих пор оставался открытым. Исходя из количества запасаемого на зиму корма и динамики роста корней зубов Н.Г. Евдокимов (2001) высказал предположение о том, что обыкновенная слепушонка является зимоспящим видом. Для проверки этого предположения в период с октября 2006 по май 2007 года у 5 особей обыкновенной слепушонки проводили непрерывную регистрацию температуры тела в естественных условиях. Ни в одном из 4739 измерений, полученных для каждой особи, температура тела животных не опускалась ниже 30°С, что означает отсутствие эпизодов спячки или торпора, для которых характерно падение температуры до 20°С и ниже.
Достоверный вклад в изменчивость стандартного обмена (F2,89=3,1; P<0,05) и метаболического индекса (F2,82=3,4; P<0,05) вносит окраска меха слепушонок. В Западно-Сибирских популяциях, полиморфных по окраске меха, бурые особи имеют достоверно более высокий уровень стандартного обмена, чем черные особи в этих же популяциях и бурые из южных популяций, где черная морфа отсутствует (1,89±0,13, 1,59±0,08, 1,46±0,38 мл*г-1*ч-1, соответственно (Критерий Тьюки; P<0,05)). Метаболический индекс у бурых особей в северной части зоны их обитания (в полиморфных популяциях) оказывается достоверно более низким, чем в южной (2,8±0,3 и 4,0±0,3, соответственно; критерий Тьюки, P<0,05). Снижение резервных возможностей метаболизма у бурых особей в северной части зоны их обитания, является, очевидно, одним из факторов, ограничивающих распространение данной окрасочной морфы на север (Novikov et al., 1996).