
Физиологическая цена адаптаций к подземному образу жизни: обыкновенная слепушонка ( ellobius talpinus pall.) В сравнении с наземными грызунами
| Вид материала | Автореферат диссертации |
| Затраты на соматическую и генеративную продукцию Затраты на защиту от паразитов Роль стресса в формировании адаптационной стратегии |
- Председатель, 417.14kb.
- Адаптации животных к нормальным и экстремальным факторам среды, 44.24kb.
- Рабочая программа дисциплины «Адаптивная изменчивость организмов» Код дисциплины, 90.26kb.
- Понятие адаптации, 125.13kb.
- Паразитология балашов Ю. С. Паразито-хозяинные отношения членистоногих с наземными, 282.93kb.
- Литература. Тема урока: «Очерк жизни и творчества И. А. Гончарова. Три романа- «Обыкновенная, 18.56kb.
- Программа «Формирование активной жизненной позиции и стремления к здоровому образу, 78.18kb.
- Сорта яблонь антоновка обыкновенная (зимний), 137.97kb.
- Задачи: дать понятие о роли здоровья в жизни человека; прививать детям стремление, 75.46kb.
- «Приветствие», 177.29kb.
ЗАТРАТЫ НА СОМАТИЧЕСКУЮ И ГЕНЕРАТИВНУЮ ПРОДУКЦИЮ
У самок млекопитающих основные затраты на репродукцию связаны с вынашиванием плодов и лактацией (Gittleman, Thompson, 1988). Так, у мышевидных грызунов увеличение потребления энергии беременными самками по сравнению с небеременными составляет 18-25% (Kaczmarski, 1966; Migula, 1969; Mattingly, McClure, 1982). Во время лактации количество ассимилируемой энергии увеличивается на 45-90% (Weyner, 1987). У самцов, приспособленность которых определяется, главным образом, успехом в конкуренции за репродуктивные ресурсы (Maynard Smith, 1977; Clutton-Brock, Harvey, 1978), прямые затраты на размножение (продукция гамет и спаривание) оказываются значительно меньшими, чем затраты на сопутствующие формы поведения (Gittleman, Thompson, 1988). В таблице 4 приведены наши данные, в ряде случаев дополненные литературными (Башенина, 1977; Юдин и др., 1979; Летицкая, 1984; Евдокимов, 2001; Gębczyński, 1983), которые позволяют сопоставить прямые и непрямые затраты на размножение, соматическую и генеративную продукцию у слепушонок, красной полевки и джунгарского хомячка.
Одним из ключевых факторов, определяющих не только репродуктивный успех особи, но и структуру ее энергобюджета, является уровень секреции половых гормонов (Kenagy, 1980; Beatty, 1992; Wade, Schneider, Li, 1996). Концентрации тестостерона в крови у половозрелых самцов обыкновенной слепушонки в сезон размножения были значительно ниже, чем у наземных мышевидных грызунов (табл. 4), причем только у 65,1% самцов уровень тестостерона был выше порога обнаружения. Вне сезона размножения тестостерон в крови самцов находился в следовых количествах. Обыкновенная слепушонка значительно уступает наземным видам грызунов по величине весового индекса семенников. Семенные пузырьки и препуциальные железы у самцов слепушонок не развиты.
При попарном ссаживании на нейтральной территории только что отловленные особи обыкновенной слепушонки практически не проявляли агрессии (Большаков и др., 1989; Мошкин и др., 1991; Мошкин и др., 1999). Однако в тестах, проведенных в апреле на самцах (8 пар), отловленных в октябре предыдущего года (т.е. содержавшихся в виварии в течение всей зимы), среднее количество драк превышает аналогичный показатель у полевок и сопоставимо с таковым у джунгарских хомячков (табл.4).
Таблица 4.
Показатели соматической и генеративной продукции у сравниваемых видов.
| Показатель | Красная полевка | Джунгарский хомячок | Обыкновенная слепушонка |
| Репродуктивные характеристики самцов | |||
| Концентрация тестостерона (нг*мл-1) | 22 0,640,12 | 24 0,670,15 | 20 0,080,02 |
| Агрессивность в тесте парного ссаживания | 24 3,3±0,9 | 22 14,3±2,9 | 8 13,04,1 |
| Содержание белка в моче (мг*мл-1) | 31 1,470,32 | - | 38 0,370,08 |
| Весовой индекс семенников | 111 2,02±0,49a | 12 1,70±0,25a | 64 0,16±0,01b |
| Репродуктивные характеристики самок | |||
| Фактическая плодовитость | 136 5,70,1 | 17 5,50,4 | 6 4,3±0,4 |
| Число приносимых пометов (в год) | 2-4 | 3-5 | 2 |
| Интервал между пометами (дн.) | 20 | 20 | >30 |
| Сезон размножения | Май-сентябрь | Май-сентябрь* | Февраль-май |
| Общие энергозатраты за сезон (кДж) | 4300 | 3000 | 1500-2000 |
| Характеристики соматической продукции | |||
| Масса новорожденных (г) | 12 1,5-1,8 | 14 1,4±0,03 | 10 3,10,1 |
| Процент от веса матери | 5,3 | 5,5 | 7,5 |
| Константа весового роста (1-30 день) | 0,472 (0,581*) | 0,588 (0.665*) | 0,438 (0,469*) |
| Вес к моменту окончания лактации (% от веса взрослого) | 60* | 60 | 48 |
| Продолжительность лактации (дн.) | 18-20 | 15 | 25 |
| Возраст вступления в размножение (мес.) | 1-1,5 | 1,5-2 | >12* |
| Продолжительность жизни | В природе - до 1 года | В неволе – до 3-х лет | В природе – до 6 лет* |
Примечания: В верхней строчке ячеек приведен объем выборок
* - литературные данные
Эффективность химической коммуникации грызунов во многом определяется содержанием в моче белков, связывающих летучие молекулы (Vandenbergh et al., 1975; Lombardi et al., 1976). Продукция основного белка мочи определяется уровнем секреции андрогенов (Finlayson, et al., 1965) и составляет у самцов мышей 5-10 мг в сутки - до 10% всего азотного обмена особи (Berger, Szoka, 1981; Cavaggioni, Mucignat-Caretta, 2000). У красной полевки достоверное влияние на содержание белка в моче оказывали месяц наблюдений (F1,53=13,1; P<0,001) и пол животных (F1,53=7,4; P<0,01). В июне содержание белка в моче было выше, чем в августе, самцы имели более высокое его значение (табл.4), чем самки (t53=2,6; P<0,01). У самцов концентрация белка в моче коррелировала с массой репродуктивных органов (r=0,47; P<0,01 для семенников и r=0,71; P<0,001; N=31, для препуциальных желез). Половозрелые самки по содержанию белка в моче не отличались от неполовозрелых. Обыкновенная слепушонка имеет, в целом, более низкие, чем красная полевка, концентрации основного белка мочи (табл.4). В отличие от последней, концентрации белка в моче у слепушонки не подвержены сезонной изменчивости, не различаются у самцов и самок и не зависят от репродуктивного статуса особей.
Наблюдение за размножением восточной (Летицкая, 1984; Якименко, Ляпунова, 1987) и обыкновенной (наши данные) слепушонок в неволе показало, что эти виды характеризуются значительно более продолжительной, чем у наземных грызунов сопоставимого размера беременностью (30 дней) и столь же продолжительной лактацией (не менее 25 дней). В северной части ареала обыкновенной слепушонки самки приносят в год не более двух пометов (Евдокимов, Позмогова, 1993, Евдокимов, 2001). Н.Г. Евдокимов и В.П. Позмогова (1993), исследовав несколько сотен семей в разных частях ареала, пришли к выводу, что в каждой из семей размножается только одна самка, вследствие чего многие особи в течение нескольких лет жизни остаются яловыми. По данным Н.Г.Евдокимова (2001), сеголетки вообще не участвуют в размножении, а основной пик репродуктивной активности самок приходится на возраст 2-4 года. Самки могут размножаться до 6 сезонов и приносить, в общей сложности, до 57 детенышей, однако только небольшой (15,2%) части их удается в течение жизни принять участие в размножении (Евдокимов, 2001).
Отловы обыкновенной слепушонки, проводившиеся в Новосибирской области в период со второй половины апреля по средину ноября, показали, что размножение на северо-восточной периферии ареала вида полностью прекращается к началу лета (табл. 4). Средняя плодовитость обыкновенной слепушонки соответствует типичным для вида (Евдокимов, 2001) значениям. В весенних отловах доля самок, участвующих в размножении, колеблется в разные годы от 13 до почти 40%, в целом составляя 24,2±3,3% (10,9±1,6% от общего числа особей в популяции), что несколько превышает показатели, приведенные в работе Н.Г.Евдокимова (Евдокимов, 2001). В изучаемой нами популяции из одного гнезда удавалось одновременно отловить до 3-х участвующих в размножении самок. Среднее число размножающихся самок в 36 обследованных семьях составило 1,40,1 самки на семью.
У слепушонок, отловленных в октябре и содержащихся в виварии в течение зимы, размножение начиналось в конце января и заканчивалось в конце апреля. Размножающиеся самки приносили по 1-2 помета, состоящих, в среднем, из 2,80,4 детенышей. При содержании группами по 4 особи (2 самца и 2 самки) потомство приносили обе самки. Таким образом, как в изучаемой нами популяции, так и в популяциях Урала и Зауралья, ежегодный репродуктивный выход самки составляет, в среднем, 6-8 детенышей.
В популяциях обыкновенной слепушонки, обитающих в Уральском регионе, отчетливо выражен сдвиг соотношения полов в сторону самцов, связанный очевидно с большей смертностью и большей миграционной активностью самок (Евдокимов, 2001). В популяциях, обитающих в Новосибирской области, соотношение полов у взрослых особей достоверно не отличалось от 1:1 (χ2=3,6; df=1; P=0,06).
Несмотря на то, что детеныши обыкновенной слепушонки при рождении имеют более крупные, чем у наземных грызунов размеры, их удельная скорость роста, а следовательно, и удельные затраты энергии матери на выкармливание, меньше, чем у наземных грызунов (табл. 4).
Непосредственные наблюдения за меченными животными, проведенные в Уральском регионе, показывают, что в природе продолжительность жизни обыкновенной слепушонки может достигать 6 лет (Евдокимов, 1997; 2001). Согласно оценкам, сделанным Н.В.Синевой на основании возрастной шкалы, разработанной Н.Г.Евдокимовым (2001), в Новосибирской области особи старше 2-х лет в отловах отсутствовали.
ЗАТРАТЫ НА ЗАЩИТУ ОТ ПАРАЗИТОВ
Помимо отклоняющих воздействий физической природы практически любой организм в процессе своей жизнедеятельности неизбежно сталкивается с риском внедрения в него паразитов (в широком смысле этого слова, т.е. организмов, существующих за счет хозяина). Соответствующая защитная функция включает в себя широкий спектр конституциональных (структурных) адаптаций, неспецифические поведенческие и физиологические реакции и собственно иммунитет (Ройт и др. 2000). Среди врожденных защитных реакций наиболее ощутимым для энергобюджета является, очевидно, синдром болезни, который выражается в снижении количества ассимилируемой энергии, повышении температуры тела (Kruk et al., 1998; Akins, et al., 1996) и снижении двигательной активности (Van Damm et al., 1985; Stettler et al., 1992; Dantzer, 2001). Приобретенный иммунитет, связанный с определенными затратами энергетических и пластических ресурсов на пролиферативные процессы и синтез белковых антител (Buttgereit et al., 2000), также оказывает значимое влияние на энергобаланс организма (Behnke, et al., 1992; Shudo, Iwasa, 2001; Ots et al., 2003). Однако реальная физиологическая цена иммунного ответа до сих пор является предметом дискуссий (Lochmiller, Deerenberg, 2000; Raberg et al., 2002; Derting, Compton, 2003; Schmid-Hempel, 2003). Видоспецифичность патогенного действия паразитов и неодинаковая устойчивость к ним хозяев делает проблематичным использование экспериментального заражения животных в качестве инструмента для сравнительной оценки затрат на поддержание и активацию иммунитета. Поэтому в качестве показателя этих затрат мы использовали величину физиологического ответа животного на стандартные иммуногенные факторы – бактериальные липополисахариды (ЛПС), активирующие неспецифическую защитную реакцию (синдром болезни), и эритроциты барана, стимулирующие выработку специфических антител.
Введение бактериального липополисахарида (использовали промышленный препарат липополисахарида E. сoli L-4005, серотип 055:B5, «Сигма») джунгарским хомячкам (16 особей) и обыкновенной слепушонке (8 особей) вызывало у обоих видов типичные проявления синдрома болезни. В первые сутки после инъекции у животных снижались двигательная активность и среднесуточный уровень метаболизма. Удельные энергозатраты на один акт двигательной активности (которые можно рассматривать как энергетическую цену локомоции) в 1-2 сутки после инъекции ЛПС у обоих видов были достоверно выше, чем до инъекции (рис.3). Температура тела, оцененная у ЛПС-стимулированных животных обоих видов через 3 часа после инъекции, была достоверно выше, чем у контрольных, которым вводили физиологический раствор. У ЛПС-стимулированных хомячков она равнялись 37,3±0,1°С у контрольных - 36,2±0,2°С (t14=5,7; P<0,001). У слепушонки соответствующие значения показателей составляли 37,0±0,3°С и 36,2±0,04°С (t6=2,7; P<0,05). Введение подопытным животным обоих видов эритроцитов барана не вызывало у них достоверных изменений метаболической и двигательной активности.
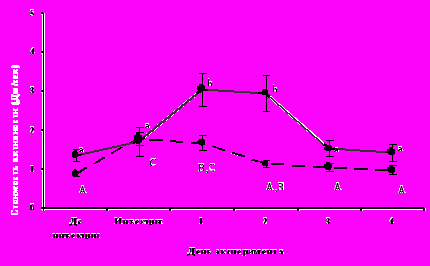 Рис.3. Удельная стоимость двигательной активности у обыкновенной слепушонки (сплошная линия) и джунгарского хомячка (пунктирная линия) до- и после введения бактериального липополисахарида. Для каждого вида значения, достоверно (критерий Тьюки; P<0,05) различающиеся между собой, помечены разными буквами.
Рис.3. Удельная стоимость двигательной активности у обыкновенной слепушонки (сплошная линия) и джунгарского хомячка (пунктирная линия) до- и после введения бактериального липополисахарида. Для каждого вида значения, достоверно (критерий Тьюки; P<0,05) различающиеся между собой, помечены разными буквами.Анализ величины гуморального иммунного ответа на введение эритроцитов барана у грызунов, отловленных в природе, показывает, что слепушонка имеет минимальные в ряду рассмотренных видов значения этого показателя (рис.4). В отличие от наземных грызунов, для которых характерно снижение величины гуморального иммунного ответа при достижении половозрелости (Мак и др., 2002; Moshkin et al., 1998; Saino et al., 2000), у обыкновенной слепушонки в сезон размножения иммунный ответ размножавшихся самок практически не отличался от такового у не размножавшихся (10,±1,6 и 12,1±1,0 АОК*г-1, соответственно). Корреляция между весенней массой семенников самцов и величиной гуморального иммунного ответа также отсутствовала (rs= -0,11; N=72; P=0,40).
Таким образом, величина гуморального иммунного ответа на незнакомый антиген у обыкновенной слепушонки оказывается значительно ниже, чем у наземных грызунов. Однако в отличие от последних, иммунореактивность у слепушонки не зависит от функционального состояния организма и видимо не конкурирует за ресурсы с репродуктивной функцией.
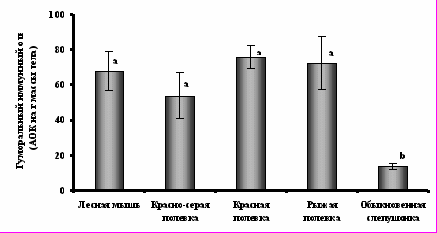
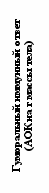 Рис. 4. Величина гуморального иммунного ответа у обыкновенной слепушонки в сравнении с наземными видами грызунов. Значения, достоверно (критерий Тьюки; P<0,05) различающиеся между собой, помечены разными буквами.
Рис. 4. Величина гуморального иммунного ответа у обыкновенной слепушонки в сравнении с наземными видами грызунов. Значения, достоверно (критерий Тьюки; P<0,05) различающиеся между собой, помечены разными буквами.РОЛЬ СТРЕССА В ФОРМИРОВАНИИ АДАПТАЦИОННОЙ СТРАТЕГИИ
Стресс-реакция является основным неспецифическим механизмом мобилизации энергетических и пластических ресурсов организма и их перераспределения в пользу приоритетной функции для компенсации внешних воздействий, нарушающих гомеостаз (Горизонтов, 1976; Меерсон, 1981). Универсальный характер организменного ответа на стрессирующие воздействия и многообразие физиологических эффектов гипоталамо-гипофизарно-надпочечниковой системы (ГГНС) позволяют ей эффективно регулировать распределение ресурсов между различными функциями: локомоторной, терморегуляторной, репродуктивной и иммунной (Селье, 1960; Хаскин, 1975; Коренева, Шхинек, 1988; Weber et al., 2002).
В качестве показателя мобилизационных возможностей организма обычно используется величина адренокортикальной реакции на стандартные стрессирующие воздействия - отношение концентрации глюкокортикоидных гормонов после воздействия к их фоновой концентрации, которая, в свою очередь, отражает кумулятивный эффект предшествующих стрессов и, следовательно, может рассматриваться как индикатор «благополучия» животного (Boonstra, 2004). Экспериментальное моделирование стрессовых ситуаций различной природы (глубокое охлаждение, иммобилизация, социальный конфликт, антигеная стимуляция) показало, что по сравнению с наземными грызунами обыкновенная слепушонка имеет пониженную адренокортикальную реактивность на все типы стрессоров (рис. 5).
Рис. 5. Величина адренокортикальной реакции на стандартные стресс-стимулы у обыкновенной слепушонки и наземных грызунов. Знаком «*» отмечены случаи достоверных (критерий Тьюки; P<0,05) различий соответствующих показателей у слепушонки и наземных видов грызунов.
Многочисленные данные свидетельствуют о том, что на определенных этапах онтогенеза уровень функциональной активности ГГНС может оказывать влияние на темпы полового созревания особи (Novikov, Moshkin, 1998; Marchlewska-Koj et al., 2003), ее функциональное место в пространственно-этологической структуре популяции (Sapolsky, 1982; Сreel, 2001) и выживаемость в экстремальных условиях (Romero, Wikelski, 2001; Rogovin et al., 2003). Контролируя уровень ресурсных «инвестиций» в репродуктивную функцию, стресс, тем самым, определяет продолжительность различных этапов онтогенеза животного и, следовательно, может рассматриваться как универсальный механизм модификации онтогенетических программ в рамках видовой адаптационной стратегии.
Одним из наиболее известных примеров такой модификации является индивидуальная изменчивость мышевидных грызунов по срокам вступления в размножение и общей продолжительности жизни (Оленев, 2002). Наши данные показывают, что одной из возможных причин задержки полового созревания сеголеток красной полевки при высокой плотности популяции является их повышенная стрессированность (Novikov, Moshkin, 1998). Данные, полученные при изучении голого землекопа, свидетельствуют о том, что наименее стрессированной особью в семейной группе является размножающаяся самка (Faulkes, Abbot, 1997). Однако у обыкновенной слепушонки в сезон размножения половозрастные различия между членами одной и той же семьи по уровню стрессированности отсутствовали. В то же время, в семьях, где осенью животные имели базальные концентрации глюкокортикоидов ниже 200 нг*мл-1, к следующей весне сохранилось 85,7±9,7% самцов, тогда как в семьях с концентрациями гормонов свыше 400 нг*мл-1 - только 33,3±11,4% (различия достоверны, t30=3,3; P<0,01). Следовательно, не влияя непосредственно на участие в размножении, активация ГГНС у слепушонки стимулирует миграционную активность либо гибель части особей, регулируя тем самым, численность семейных групп (Новиков и др., 2008).
В целом, базальные концентрации глюкокортикоидов в изучаемой популяции (347,2±38.5 нг*мл-1) оказываются достоверно выше (критерий Тьюки; P<0.05), чем у особей, отловленных в Северном Казахстане (100,3±6,2 нг*мл-1), и на Южном Урале (196,1±8.9 нг*мл-1) (Большаков и др., 1989; Мошкин и др., 1991). В отличие от популяций Уральского региона, животные из которых реагировали на получасовую иммобилизацию достоверным (t136=5,2; P<0.001) увеличением концентраций глюкокортикоидов, у слепушонок из Новосибирской области концентрации глюкокортикоидов после иммобилизации (368.2±35.1 нг*мл-1) достоверно не отличались от базальных (t65=0,4; P>0,05). Таким образом, как и у красной полевки (Мошкин и др., 1993; Novikov et al., 1993), у обыкновенной слепушонки уровень стрессированности увеличивается в экологически пессимальных условиях.
ЗАКЛЮЧЕНИЕ
Проведенный нами сравнительный анализ основных параметров жизнедеятельности организма – интенсивности метаболизма, соматической и генеративной продукции, терморегуляции, затрат на защиту от паразитов и величины адренокортикальной реакции на стресс у мышевидных грызунов различной экологической специализации показал наличие значительной межвидовой изменчивости по большинству рассмотренных показателей.
Обыкновенная слепушонка – семисоциальный вид, ведущий подземный образ жизни, среди рассмотренных нами грызунов занимает исключительное положение. Специфика среды обитания определяет низкую интенсивность обменных процессов социальных подземных грызунов и редуцированную способность к активному поддержанию постоянной температуры тела в покое (McNab, 1979; Withers, Jarvis, 1980). Однако, в отличие от тропических подземных видов, низкий уровень обмена и подавление активной терморегуляции у слепушонки отмечаются только в теплое время года. В холодное время года по своим терморегуляторным и метаболическим характеристикам этот вид не отличается от симпатричных наземных грызунов. Можно предполагать, что летнее снижение затрат на поддержание температурного гомеостаза связано не только с проблемами его ресурсного обеспечения, но и с подавлением механизмов несократительного термогенеза, высокая реактивность которых сочетается со снижением КПД мышечной работы (Якименко и др., 1971). Последнее обстоятельство имеет большое значение в теплое время года - период наиболее интенсивной роющей деятельности.
Слабая метаболическая и терморегуляторная реакция на охлаждение сочетается у обыкновенной слепушонки с низкой адренокортикальной реактивностью как на физические так и на эмоциональные стрессоры. Это хорошо согласуется с представлением об адаптивном снижении стресс-реактивности в условиях, когда эмоциогенные воздействия преобладают над физическими (Беляев и др., 1977; Ganem, 1991; Moshkin et al., 2002).
В отличие от наземных мышевидных грызунов, обыкновенная слепушонка имеет значительно меньшую скорость пре- и постнатального развития, более поздние сроки вступления в размножение и значительно более низкую репродуктивную активность. Даже на периферии видового ареала, где высокая смертность приводит к увеличению числа размножающихся самок, размер и количество приносимых за сезон выводков у слепушонки оказываются меньше, чем у наземных грызунов. Половозрелые самцы в сезон размножения у слепушонки имеют значительно меньшую, чем у наземных видов массу репродуктивных органов и значительно более низкую концентрацию тестостерона в крови. Специфика образа жизни и пространственно-этологической структуры популяций позволяет обыкновенной слепушонке снижать затраты на маркировочное поведение, обусловленные экскрецией низкомолекулярных белков, экономя тем самым пластические ресурсы организма,.
Относительная изоляция семейных группировок в сочетании со спецификой паразитарного окружения определяют низкую функциональную активность адаптивного иммунитета. В отличие от наземных грызунов с высокой интенсивностью размножения, у обыкновенной слепушонки отсутствует репродуктивная иммуносупрессия. В то же время, по характеру физиологического ответа на введение бактериальных липополисахаридов слепушонка не отличается от джунгарского хомячка. Можно предполагать, что данная неспецифическая реакция, представляющая собой первую линию защиты от паразитов, при семейно-групповом образе жизни играет решающую роль в предотвращении распространения инфекций.
Таким образом, помимо морфологических адаптаций к роющему образу жизни, обыкновенная слепушонка характеризуется рядом уникальных физиологических особенностей, соответствующих единой адаптационной стратегии, которая состоит в минимизации затрат на все стороны жизнедеятельности. «Экономное» расходование ресурсов организма в сочетании с хорошими защитными свойствами среды позволяет обыкновенной слепушонке достигать высокой для грызуна такой массы тела продолжительности жизни (Евдокимов, 1997; 2001) и обеспечивает достаточное количество ресурсов для высокозатратной локомоторной активности. Прямой оценки энергозатрат на роющую деятельность у слепушонок не проводили, но вряд ли они принципиально отличаются от таковых, измеренных у других видов подземных грызунов (Vleck, 1979). Кроме того, морфологические особенности локомоторного аппарата слепушонки увеличивают и стоимость перемещения по твердому субстрату.
В северной части видового ареала размножение обыкновенной слепушонки начинается еще под снегом и заканчивается в конце весны. Функциональная активность терморегуляторной и иммунной систем в сезон размножения оказываются даже выше, чем после его окончания. Можно предполагать, что в условиях обилия и высокой доступности пищи в период предшествующий началу вегетации сберегающая стратегия обыкновенной слепушонки позволяет ей обеспечивать ресурсами как репродуктивную, так и защитные функции организма. Однако возможности их ресурсного обеспечения снижаются по мере уменьшения доступности корма после начала вегетации, когда снижается кормовая ценность подземных частей растений. В частности, на модельном участке разнотравно-злакового луга, представляющего собой типичное местообитание обыковенной слепушонки, в период с 3 мая по 3 июля 2007 г. общая подземная фитомасса уменьшилась с 1130 до 710 г сухого вещества на м2, а сырая масса корней зопника клубненосного – основного кормового растения слепушонки в зимний период – с 185 до 120 г на м2. В соответствии с этим летом у слепушонки практически прекращается размножение, подавляется активная терморегуляция и снижается иммунореактивность.
Популяции обыкновенной слепушонки, которые рассматривали в данной работе, обитают на Северо-Восточной периферии видового ареала. Сопоставление наших результатов с данными, полученными при изучении других популяций вида (Большаков и др., 1989; Мошкин и др., 1991), показывает, что в условиях «географического пессимума» повышается стрессированность животных и уменьшается величина их адренокортикальной и метаболической реакции на модельные стрессоры. Повышенная смертность компенсируется, очевидно, увеличением числа размножающихся самок. Поэтому средняя численность семейных группировок и локальная плотность в предпочитаемых местообитаниях в популяциях Уральского региона и Западной Сибири не различаются (Новиков и др., 2007).
Таким образом, физиологической ценой адаптаций к подземному образу жизни у обыкновенной слепушонки является ограничение затрат на поддержание жизнедеятельности, размножение, терморегуляцию и защиту от паразитов, что позволяет обеспечивать ресурсами энергоемкую роющую деятельность. Данная адаптационная стратегия, оставаясь неизменной в своей сути, обладает, тем не мене, широкой нормой реакции, которая дает возможность эффективно перераспределять ресурсы как в эко-географических градиентах, так и в онтогенезе животных.