
Метаболизм белков у растущих бычков и свиней и факторы его регуляции 03. 00. 04 биохимия 03. 00. 13 физиология
| Вид материала | Автореферат |
| 3.3. Регуляция обмена белков и качества продукции у бычков и |
- Глицеральдегид-3-фосфатдегидрогеназа: роль в регуляции энергетического обмена, индукции, 699.32kb.
- Влияние пренатального стресса на процессы окислительной модификации белков и активность, 202.57kb.
- Рабочая программа дисциплины «биология клетки» (молекулярная биология) Код дисциплины, 225.32kb.
- «Молекулярные механизмы гормональной регуляции» Общая трудоемкость изучения дисциплины, 11.18kb.
- Тема строение белков и ферментативный катализ вопросы лекции, 137.02kb.
- Алиевич биологическое обоснование, разработка и использование полнорационных комбикормов, 942.5kb.
- Пояснительная записка Нормальная физиология наука и учебная дисциплина о жизненных, 578.98kb.
- Тема обмен белков. Вопросы лекции, 90.92kb.
- Биосинтез белков Интегрированный урок в 10-м классе(химия и биология). Цель урока, 27.63kb.
- Метаболизм соединений свинца и меди в организме собак г. Красноярска чужакин Н. Л.,, 59.93kb.
3.3. Регуляция обмена белков и качества продукции у бычков и
свиней в период интенсивного доращивания
и откорма факторами питания
Для изучения возможного регулирования метаболизма белков и качества продукции у растущих животных путем изменения уровня обеспеченности их организма аминокислотами и энергией провели серию опытов на бычках холмогорской породы и свиньях двух- и трехпородных помесей на разных стадиях их выращивания и откорма.
В двух экспериментах, проведенных в виварии института на 20 бычках методом групп (по 5 животных в группе) в период с 10- до 15- месячного возраста, исследовали обмен азотистых веществ и количественные аспекты синтеза и распада белков скелетных мышц при разной обеспеченности метаболических процессов обменным белком и лимитирующими аминокислотами. Разный уровень обеспеченности организма бычков аминокислотами методически моделировался путем применения разных по составу комбикормов. Для животных контрольных групп применяли типовой комбикорм на ячменной основе с подсолнечниковым шротом в качестве источника протеина с высокой распадаемостью в рубце. Для бычков опытных групп из состава комбикорма исключали подсолнечниковый шрот, но вводили кукурузный глютен и соевый шрот в качестве источников протеина с низкой распадаемостью и для балансирования содержания некоторых незаменимых аминокислот (лизин, метионин, лейцин). Кроме того во втором эксперименте к вышеуказанным компонентам добавляли пивную дробину, диаммонийфосфат, серу и органические кислоты (яблочная, лимонная и фумаровая) для поддержания микробиологических процессов в рубце на высоком уровне. Во всех экспериментах рационы содержали одинаковые количества сырого протеина и обменной энергии. Распадаемость протеина корма в контрольных группах составляла 71 %, а в опытных в первом опыте – 63 %, во втором – 64 %. Уровень обеспеченности организма растущих бычков обменным белком и доступными аминокислотами проводили расчетным путем. Расчеты проводили совместно с доктором биологических наук Харитоновым Е.Л. В основу расчета положены принципы, изложенные в «Методах исследований питания сельскохозяйственных животных»(1998) (ред. академика РАСХН Б.Д. Кальницкого), а также данные, полученные в эксперименте.
В наших исследованиях в результате снижения уровня распадаемого протеина потребляемого корма с 71 до 63 % возрастало количество поступающих в метаболический пул обменного белка на 10,9 % с 624 до 692 г/сутки и лизина на 4,6 %, лейцина на 19,2 %, гистидина на 8,3 %, метионина на 4,8 %.
В результате улучшения обеспеченности процессов метаболизма аминокислотами в условиях изопротеиновых и изокалорийных рационов в период интенсивного доращивания и откорма позволило на 8,7 % увеличить среднесуточный прирост живой массы животных (у бычков опытной группы он составил 138424, у контрольных 127325 г) (р<0,01). В целом за опыт прирост живой массы у бычков контрольной группы составил 90,41,76 кг, а у животных опытной группы – 98,31,68 кг, что на 8,7% больше (р<0,01). При этом у бычков опытной группы за весь период эксперимента повышалась не только интенсивность роста, но и эффективность использования корма.
Результаты балансовых опытов показали, что при практически одинаковой переваримости сырого протеина в опытной группе, отложение азота было больше за счет меньшего выделения его с мочой, что свидетельствует о более высокой эффективности использования у них азота в обменных процессах (табл. 6). У животных опытной группы по сравнению с контролем обнаружено снижение уровня свободных аминокислот в плазме крови. При этом заметное снижение отмечалось по незаменимым аминокислотам. Кроме того в плазме крови у животных этой группы обнаружена достоверно более низкая концентрация мочевины, которая сопровождалась снижением отношения азота, выделенного с мочой, к азоту, принятому с кормом и переваренному (табл.6).
Таблица 6
Использование азота корма и концентрация свободных аминокислот и мочевины в плазме крови у бычков с живой массой 300 кг
| Показатели | Группы | |
| контроль | опыт | |
| Принято азота с кормом, г/сутки | 162,4 4,0 | 164,6 1,6 |
| Выделено с калом, г/сутки | 54,3 1,6 | 55,4 1,2 |
| Переварено азота, г/сутки | 109,7 4,0 | 109,2 2,5 |
| % | 66,8 0,9 | 66,3 1,0 |
| Отношение азота мочи/ азоту принятому с кормом | 0,41 | 0,36 |
| Отношение азота мочи/ азоту переваренному | 0,61 | 0,55 |
| Мочевина до кормления, ммоль/л | 4,76 ± 0,15 | 4,20 ± 0,20 |
| Мочевина через 3 часа после кормления, мкмоль/л | 5,42 ± 0,16 | 4,71 ± 0,21* |
| Сумма свободных аминокислот, мкмоль/л | 1446,5 ± 75,4 | 1241,8± 68,3 |
| в т. ч. незаменимые, мкмоль/л | 786,6 ± 32,0 | 682,4 ± 28,4* |
| заменимые, мкмоль/л | 659,9 ± 0,38 | 559,4 ± 0,34 |
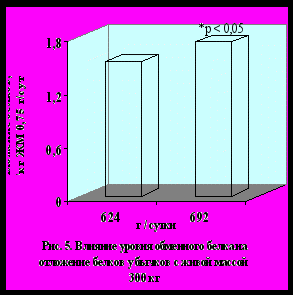
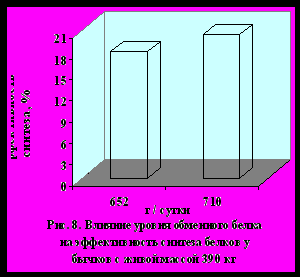
Уровень обеспеченности метаболических процессов обменным белком и лимитирующими аминокислотами оказывает существенное влияние на обмен белков в мышечной ткани бычков и, соответственно, на показатели мясной продуктивности и качество мяса. Бычки опытной группы по интенсивности наращивания скелетной мускулатуры и по отложению белков (на 17,4 %, р < 0,05) в мышцах превосходили контрольных животных. Более высокое отложение мышечных белков у бычков опытной группы обусловлено интенсификацией процессов синтеза белков, которая превалировала над его распадом. В целом эффективность синтеза белков в опытной группе возросла с 18,5 до 20,6 %.
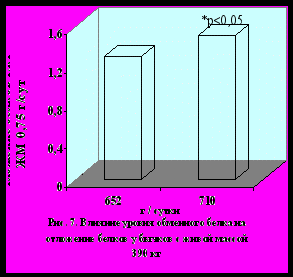
Во втором эксперименте изменения метаболизма белков у бычков с живой массой 390 кг под влиянием доступных для усвоения обменного белка и аминокислот имели ту же направленность, что и в первом опыте. При обобщении результатов всех опытов установили, что интенсивность и направленность метаболизма белков в скелетных мышцах существенно зависела от уровня обеспеченности организма бычков обменным белком и лимитирующими аминокислотами в условиях интенсивного выращивания и откорма (рис. 5, 6, 7, 8).
Корреляционный анализ данных экспериментов показал, что интенсивность отложения и эффективность синтеза мышечных белков положи-
| Влияние уровня доступных для усвоения аминокислот (г/сутки) на метаболизм мышечных белков у бычков | |
 |  |
 |  |
тельно и тесно коррелирует с уровнем доступного для усвоения белка (r = + 0,95; p < 0,04 и r = + 0,97; p < 0,01) и лизина (r = + 0,95; p < 0,05 и r = + 0,88; p < 0,10).
Следовательно, повышение обеспеченности организма бычков доступным для обмена белком и лимитирующими аминокислотами в результате применения кормов с пониженной распадаемостью протеина и крахмала в рубце в период интенсивного доращивания и откорма способствует интенсификации биосинтетических процессов в мышечной ткани, увеличению отложения и повышению эффективности синтеза белков в скелетных мышцах, позволяющего в более полной мере реализовать генетический потенциал мясной продуктивности.
В проведенных нами исследованиях установлено, что использование в кормлении бычков рационов с низкой распадаемостью протеина в преджелудках улучшает обеспеченность процессов метаболизма незаменимыми аминокислотами и повышает мясную продуктивность и качество мяса. Улучшение обеспеченности обменным белком и лимитирующими аминокислотами метаболических процессов в мышечной ткани растущих бычков повышает интенсивность синтеза белков (на фоне отсутствия существенных изменений по скорости обновления), массу скелетных мышц и сопровождается снижением катаболизма аминокислот в тканях и процессов мочевинообразования. В конечном итоге эти изменения в интенсивности биосинтетических процессов в организме бычков способствуют повышению эффективности использования азотистых веществ корма и в целом улучшению конверсии протеина корма в продукцию.
Как известно, поток глюкозы из кишечника жвачных в метаболический пул в определенной мере связан с распадаемостью в рубце крахмала корма. В опыте на бычках, выращиваемых на мясо, введение в рацион кукурузы, как источника крахмала с пониженной расщепляемостью в рубце, обеспечило дополнительное поступление глюкозы в кровь. Данный факт подтверждается увеличением в крови бычков концентрации глюкозы на 6,5; 11,4 и 9,4 % до приема корма и через 1 и 3 часа после него, соответственно (по данным Матвеева В.А. и др., 2003). В данной ситуации улучшение обеспеченности метаболических процессов глюкозой при интенсивном выращивании и откорме бычков способствует реутилизации аминокислот, высвобождаемых при разрушении белков. Более эффективная реутилизация аминокислот, в свою очередь, сопровождается снижением интенсивности их окисления. Об этом свидетельствуют низкие уровни свободных аминокислот и мочевины в крови, а также меньшее выделение азота с мочой у подопытных животных. Следовательно, дополнительное поступления глюкозы в кровь улучшает обеспеченность процессов метаболизма аминокислотами за счет меньшего их расходования на глюконеогенез, что в свою очередь способствует повышению эффективности синтеза белков в скелетных мышцах и увеличению массы скелетной мускулатуры.
Таким образом, количество аминокислот и глюкозы, поступающее из желудочно-кишечного тракта в метаболический пул организма, является фактором, лимитирующим интенсивность процессов биосинтеза компонентов мяса.
С целью определения оптимального уровня и соотношения незаменимых аминокислот в низкопротеиновых рационах и изучение их влияния на интенсивность роста, параметры отложения белков и липидов в теле, а также метаболизм и эффективность использования азотистых веществ корма провели эксперимент на помесных поросятах (ландрас крупная белая; Pic-402 крупная белая) в период доращивания. По принципу парных аналогов в возрасте 65 суток (живая масса 20 кг) были сформированы три группы по 7 голов в каждой. Содержание групповое в клетке, поение из автопоилок. Опыт продолжался до достижения живой массы 42-52 кг.
Животные 1-й (отрицательный контроль) группы получали основной рацион с содержанием сырого протеина 11,91 % без добавления синтетических аминокислот. Поросятам 2-й (опытной) группы к вышеуказанному основному рациону были добавлены синтетические аминокислоты (треонин, лизин, метионин). Так, уровень лизина был доведен до 8,1 г в кг корма.
У поросят 3-й группы содержание сырого протеина в рационе было доведено до 15,37 % путем дополнительного введения в состав комбикорма соевого шрота, что на 10 % ниже уровня существующих норм (Калашников А.П. и др., 2003). Комбикорма для свиней 2-й и 3-й опытных групп содержали одинаковые количества лимитирующих аминокислот – лизина, метионина и треонина. При этом, были определены относительные уровни незаменимых аминокислот по отношению к лизину и соотношение лизина к обменной энергии в рационах поросят в период доращивания (табл. 7).
Таблица 7
Соотношение аминокислот в кормах
поросят в период доращивания (по отношению к лизину)
| Аминокислоты | Группы | ||
| 1-я (контроль) | 2-я (опыт) | 3-я (опыт) | |
| Лизин | 100 | 100 | 100 |
| Метионин + Цистин | 82,6 | 59,9 | 60,0 |
| Треонин | 82,1 | 62,3 | 62,5 |
| Триптофан | 31,0 | 18,2 | 25,2 |
| Изолейцин | 96,4 | 56,7 | 77,4 |
| Валин | 116,6 | 68,5 | 90,2 |
| Лейцин | 170,4 | 100,1 | 131,5 |
| Гистидин | 55,0 | 32,3 | 44,1 |
| Аргинин | 121,1 | 71,2 | 103,9 |
| Фенилаланин + Тирозин | 170,6 | 100,3 | 138,4 |
| Лизин, в г на МДж обменной энергии | 0,38 | 0,65 | 0,65 |
| Уровень доступного лизина, г/сутки | 8,20 | 16,17 | 16,20 |
Аминокислотную обеспеченность биосинтетических процессов у поросят при интенсивном выращивании на низкопротеиновых рационах, обогащенных незаменимыми аминокислотами, в описываемом эксперименте определяли расчетным путем. В основу расчета положены принципы, изложенные в работах Stein H.H. et al., 2001, 2005, а также данные, полученные в эксперименте. Расчет аминокислотной доступности проводили по всем компонентам комбикорма. При этом мы допускали, что синтетические аминокислоты усваиваются в организме животных на 100%. Повышение уровня доступности лимитирующих аминокислот, а также доведение соотношения этих аминокислот до уровня «идеального» позволило на 40,7 % достоверно повысить среднесуточный прирост живой массы поросят. В конце опыта живая масса у поросят опытных групп составляла 51,50 кг, или была выше на 22,2 %, чем у животных 1-й группы. При этом затраты сырого протеина на 1кг прироста у поросят 2-й и 3-й групп были на 28,1 и 8,2 %, соответственно, ниже по сравнению с контролем. Сходная тенденция отмечалась и по затратам кормов на единицу продукции. Увеличение прироста живой массы свиней опытных групп по сравнению с контролем обусловлено большей активностью биосинтетических процессов в их организме (табл. 8). Интенсивность синтеза белков тела была выше у поросят, получавших рационы с оптимальными соотношениями лимитирующих аминокислот (по отношению к лизину) с учетом их доступности, по сравнению с животными, содержавшимися на низкопротеиновых рационах с неоптимальным их соотношением. Характерно, что наряду с повышением интенсивности синтеза белков имело место и некоторое усиление скорости их распада, а, следовательно, и обновления при относительном преобладании биосинтетических процессов. Такое состояние обмена белков приводило к повышению их уровня в организме (белкового фонда) у опытных поросят. В конечном итоге эффективность белкового синтеза, рассчитанная как отношение отложенного к синтезированному белку, заметно возросла - с 28,5 (в контроле) до 32 % (в опытных группах).
Таблица 8
Метаболизм белков тела у поросят в период доращивания, г/сут
| Показатели | Группы | ||
| 1-я (контроль) | 2-я (опыт) | 3-я (опыт) | |
| Синтез | 288 13,2 | 355 10,9 3) | 356 7,6 4) |
| Распад | 206 6,6 | 240 5,5 | 242 3,8 |
| Отложение | 82 4,97 | 113 3,93 1) | 114 9,50 3) |
| Эффективность синтеза, % | 28,5 | 31,8 | 32,0 |
| Отношение синтез/распад | 1,40 | 1,48 | 1,47 |
Изменения метаболизма белков тела у поросят опытных групп сопровождались заметным снижением концентрации мочевины, что свидетельствуют о более эффективном использовании аминокислот в биосинтетических процессах и уменьшении их дезаминирования в тканях (табл. 9). В нашем опыте подтверждением этому является повышение интенсивности синтеза белков в организме поросят (табл. 8). При этом наименьшая концентрация мочевины отмечалось у поросят, обеспеченных «идеальным» (оптимальным) соотношением незаменимых аминокислот. Более высокие величины содержания мочевины в плазме крови у поросят 3-й группы в отличие от животных 2-й группы обусловлено главным образом избыточным поступлением в организм аминокислот, не относящихся к лимитирующим аминокислотам. Аналогичная картина отмечена и по уровню свободных аминокислот в плазме крови, что вполне естественно, поскольку азот мочевины крови является продуктом катаболизма аминокислот (табл. 9). Более интенсивное расходование аминокислот в синтезе белков (главным образом – белков мышц) у поросят опытных групп способствует отвлечению из общего метаболического пула значительной части аминокислот, вследствие чего меньшая их доля используется на окисление с образованием мочевины. В этой ситуации в плазме крови опытных животных обнаружена достоверно высокая концентрация креатинина, положительно коррелирующая с массой мышц (табл. 9).
Таблица 9
Концентрация метаболитов азотистого обмена в плазме крови поросят
-
р < 0,001; 2) р < 0,02; 3) р < 0,03; 4) р < 0,04; 5) р < 0,05
Показатели
Группы
1-я (контроль)
2-я (опыт)
3-я (опыт)
Мочевина, ммоль/л
5,01 0,10
1,88 0,31 1)
4,40 0,50 2)
Креатинин, мкмоль/л
40,80 2,46
50,71 2,13 2)
49,98 1,99 3)
Гистидин, мкмоль/л
95,3 5,08
64,7 2,58 1)
104,3 10,5 2)
Фенилаланин, мкмоль/л
71,4 6,05
58,1 7,26
61,7 8,71
Треонин, мкмоль/л
195,7 5,88
134,4 11,7 5)
133,6 7,56 4)
Тирозин, мкмоль/л
61,3 5,52
49,7 9,4
64,0 3,31
Лейцин, мкмоль/л
192,0 7,62 6)
185,9 8,1 3)
275,8 8,2
Изолейцин, мкмоль/л
126,5 4,10 6)
120,4 6,57 2)
185,9 9,91
Валин, мкмоль/л
267,3 12,7 3)
215,2 11,8 2)
347,6 18,6 4)
В конечном итоге эти изменения в интенсивности и направленности метаболических процессов, вызванные лучшей аминокислотной обеспеченностью организма свиней опытных групп «идеальным» (оптимальным) соотношением лимитирующих аминокислот, способствовали повышению интенсивности роста мышечной ткани, увеличению отложения белков в скелетных мышцах (табл. 10). Отмеченные нами изменения в отложении белков в скелетных мышцах при разной обеспеченности организма свиней аминокислотами сопровождались определенными качественными изменениями в соотношении белковых фракций в мышечной ткани. Так, улучшение аминокислотной обеспеченности организма свиней опытных групп оптимальным соотношением лимитирующих аминокислот вызывает относительное и абсолютное увеличение содержания в белках мышц доли биологически полноценных белков саркоплазмы и миофибрилл при соответствующем снижении менее ценных в пищевом отношении белков стромы (табл. 10).
Исследование состава туши при убое в конце периода доращивания показало, что у свиней опытных групп по сравнению с контролем больше содержание мякоти в туше, выше убойный выход, масса туши и меньше относительное содержание сала. Повышение отношения количества мякоти к выходу жира у животных 2-й группы сопровождалось заметным снижением внутреннего жира, нежели в других группах. Эти данные свидетельствуют о лучших мясных и убойных качествах свиней опытных групп.
Таблица 10
Отложение мышечных белков и мышечной массы у свиней
| Показатели | Группы | ||
| 1-я (контроль) | 2-я (опыт) | 3-я (опыт) | |
| Мышечная масса, кг | 11,99 0,75 | 18,70 1,95 1) | 18,25 1,85 1) |
| Мышечный белок, кг | 2,02 0,13 | 3,45 0,36 2) | 3,43 0,35 2) |
| Интенсивность прироста мышечной массы, г/сутки | 92,5 12,6 | 204,3 32,6 1) | 197,8 30,8 1) |
| Интенсивность роста мышечной ткани, % / сутки | 0,77 0,07 | 1,09 0,11 3) | 1,08 0,10 3) |
| Интенсивность отложения белков в мышцах, % / сутки | 0,77 0,05 | 1,08 0,08 2) | 1,08 0,07 2) |
| Содержание белков, г%: саркоплазматических миофибриллярных стромальных | 6,23 0,34 8,01 0,30 3,72 0,21 | 7,86 0,43 3) 9,25 0,36 3) 2,88 0,27 3) | 7,90 0,39 3) 9,20 0,38 3) 2,96 0,25 3) |
Не менее важной качественной характеристикой мясной продуктивности является химический состав мяса и, в частности, содержание липидов, белков, сухого вещества. Наши результаты показали, прежде всего, высокое содержание белков, как в длиннейшей мышце спины, так и гомогенате мышц у поросят, получавших рацион с высоким уровнем доступного лизина, метионина и треонина по сравнению с контролем. Напротив, у свиней, выращиваемых на низкопротеиновых рационах с низким уровнем доступных лимитирующих аминокислот, имело место повышенное отложение липидов в мясе. Высокий уровень отложения липидов в мышцах у поросят 1-й группы можно отнести на счет усиленного использования углеродного скелета аминокислот для синтеза липидов в связи с несбалансированностью аминокислотного питания.
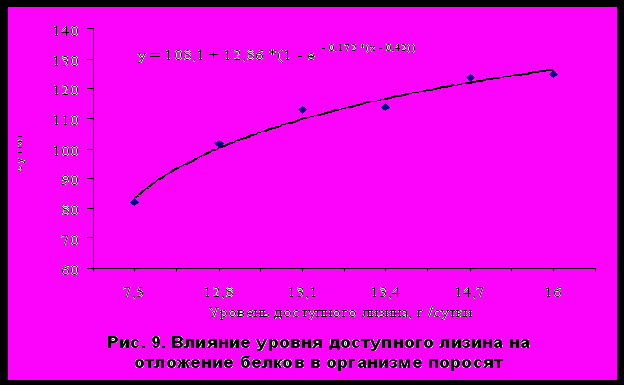
На основании обобщенных данных (с использованием метода регрессионного анализа) у поросят с живой массой от 20 до 60 кг доказана положительная зависимость отложения белков в организме от уровня доступного лизина и отношения доступного лизина к обменной энергии. С учетом выявленных взаимосвязей предложено уравнение для прогнозирования интенсивности отложения белков в теле растущих свиней (г/сутки) при разном уровне доступного для усвоения лизина (от 7,5 до 16 г/сутки) (рис. 9).
Полученные нами данные свидетельствуют о перспективности использования низкопротеиновых рационов для свиней при условии балансирования синтетическими аминокислотами состава протеина корма до уровня «идеального» протеина. Увеличение уровня доступного лизина, метионина и треонина при одновременном снижении содержания протеина до 12,06 % в рационе поросят в период доращивания обеспечили лучшие характеристики роста, метаболизма и биосинтеза компонентов мяса, а также позволили снизить избыточное выделение животными азота с мочой и
повысить эффективность использования в кормлении свиней зерна злаковых культур, содержащих низкий уровень протеина, что подтверждает необходимость совершенствования условий аминокислотного питания помесных свиней. В наших экспериментах оптимальным содержанием протеина и лимитирующих аминокислот в рационе поросят-помесей в период доращивания является: 120,6 г/кг комбикорма сырого протеина; 8,1 г/кг лизина; 4,84 г/кг метионина+цистина; 5,04 г/кг треонина. При этом отношение лизина к обменной энергии (в г на МДж) должно составлять 0,65.
Таким образом, количество аминокислот (при оптимальном их соотношении), поступающее из желудочно-кишечного тракта в метаболический пул организма, является фактором, лимитирующим интенсивность процессов биосинтеза компонентов мяса.
В другом эксперименте изучали влияние разного уровня энергии в рационе на метаболизм белков тела у трехпородных помесей (дюрок × крупная белая × крупная черная) в период доращивания с целью установления их оптимального уровня. Из поросят, отнятых в 40-дневном возрасте, было сформировано 3 группы по 10 голов в каждой. Поросята 1 группы получали комбикорм с уровнем обменной энергии 12,45 МДж/кг корма, животные 2-й группы – 13,07 МДж/кг, животные 3-й группы – 13,69 МДж/ кг при изопротеиновом рационе. Содержание животных групповое, поение из автопоилок.
Удовлетворение энергетических потребностей организма животного за счет основных энергетических источников (углеводы, жиры) способствует снижению расходования белков на эти нужды, и тем в большей мере последние смогут использоваться для анаболических целей. Увеличение уровня обменной энергии в рационе на 5 % (2-я группа) против существующих норм (1-я группа) сопровождалось повышением эффективности синтеза и отложения белков тела поросят в период доращивания. Дальнейшее повышение содержания энергии (на 10 % - 3-я группа) привело к снижению вышеуказанных показателей (табл. 11).
Таблица 11
Метаболизм белков тела поросят 105-дневного возраста, г/сутки
| Показатели | Уровень энергии в рационе, МДж/кг корма | ||
| группы | |||
| 1-я – 12,45 | 2-я – 13,07 | 3-я – 13,69 | |
| Синтез | 258,5 8,7 | 263,0 11,5 | 238,3 13,3 |
| Распад | 149,6 6,9 | 142,1 8,6 | 140,0 9,1 |
| Отложение | 108,9 8,8 | 120,9 7,9 | 98,3 8,4 |
| Эффективность синтеза, % | 42,1 | 46,0 | 41,3 |
Увеличение уровня энергии в рационе на 10 % в 3 группе относительно контроля за счет дополнительного введения жиров уже выступает как ингибирующий фактор в использовании азота. Различия в эффективности синтеза белка поросятами сопровождались неодинаковым содержанием мочевины в плазме крови и разным уровнем свободных аминокислот в мышцах. Так, у поросят 2-й группы, получавших рацион с уровнем обменной энергии 13,07 МДж/кг корма, концентрация мочевины в плазме крови и свободных аминокислот в мышцах была ниже по сравнению с животными 1-й и 2-й групп. Азотсберегающий эффект энергетической ценности рациона в проведенном опыте прежде всего проявляется в более высоких убойных показателях туши и приростах живой массы поросят. На фоне различий по обменной энергии поросята с уровнем энергии 13,07 МДж росли более интенсивно, чем в других группах (490 59 – 2-я группа, 397 53 – 3-я группа против 427 47 – 1-я группа). Исследование состава туши при убое в 105-дневном возрасте показало, что у поросят 3-й группы наряду со снижением эффективности синтеза белков тела животных происходило уменьшение выхода мяса, тогда как увеличение обменной энергии на 5 % (2-я группа) способствовало повышению доли мышечной ткани.
Таким образом, удовлетворение энергетических потребностей поросят является важным условием в повышении эффективности использования протеина корма и превращении его в белок тела. На основании полученных данных по метаболизму белков тела и направленности азотистого обмена можно заключить, что для помесных поросят (ввиду лучшей эффективности синтеза белка) является оптимальным полнорационный комбикорм с уровнем концентрации обменной энергии 13,07 МДж/кг на протяжении всего периода доращивания.
В целом проведенные исследования свидетельствуют об актуальности решения проблемы по изысканию путей повышения биоконверсии протеина корма в компоненты мяса и улучшения его качества за счет направленной регуляции метаболизма и процессов биосинтеза белка.