
Литература введение
| Вид материала | Литература |
Содержание1.3.2.Филогенетическая систематика 1.3.3.Фенетическая систематика |
- Тематическое планирование. Литература. 8 класс, 103.74kb.
- Основное содержание Введение (1ч.), 1599.38kb.
- Литература Основной учебник Введение в философию. Учебное пособие для вузов. М.: Республика,, 21.49kb.
- Литература 70, 370.34kb.
- Литература, 147.45kb.
- Литература, 195.44kb.
- Литература, 824.29kb.
- Литература, 209.32kb.
- Литература, 51.41kb.
- Литература, 1672.56kb.
1.3.2.Филогенетическая систематика Далее систематика развивалась по двум направлениям, в зависимости от положенного в основу классификации критерия - филогенетическому и фенотипическому. Филогенетичекая классификация (кладизм), отражает эволюционные связи, в основе которых лежит происхождение организмов и наследование ими определенных признаков. При построении такой системы предполагается что организмы, относящиеся к одному таксону, имели общих предков. Согласно кладизму организмы получают ранг и классифицируются исключительно в зависимости от "давности происхождения от общего предка" (9). Основные идеи кладизма восходят к выдающемуся немецкому энтомологу и систематику В. Хеннигу, который, видя недостатки отдельных систематических школ, предложил свою стройную систему логических построений в биологической систематике. Кладизм с его методами стал одним из мощных инструментов познания в систематике и филогенетике. Принципы Хеннига сводятся в основном к следующему: 1. Классификация последовательно отождествляется с филогенией, точнее даже, с генеалогией; 2. Система представляется как иерархия дихотомического (надвое) деления групп, которые должны быть строго монофилетическими; 3. Порядок соподчинения групп (относительный ранг), определяется только временем их возникновения. Статус категории, согласно теории Хеннига, зависит от положения точки ветвления на филогенетическом древе. Кладизм, учитывая ветвление, не принимает во внимание степень дивергенции, то есть темпы эволюционных преобразований. Эволюционное же родство обязательно должно включать оба эти момента. Следуя кладистам, нужно объединить в один таксон крокодилов, птиц, ящеротазых динозавров и еще две вымершие группы, а пресмыкающихся противопоставить им как другой монофилетический таксон - таксон, происходящий от одной общей предковой группы. Хотя генеалогически крокодилы более близки к птицам, их все-таки необходимо объединять с рептилиями. Птицы настолько бурно эволюционировали, что их генотипы намного сильнее отличаются как от крокодилов, так и от других рептилий. Крокодилы и ящерицы, несомненно, сохранили гораздо больше общего. Именно генетическое родство указывает на существование двух четких таксонов: Aves и Reptilia (6). Если же формально придерживаться генеалогического родства, то совершенно различные группы окажутся в одном таксоне и прогностическая ценность классификации в значительной степени пострадает. Кладисты считают, что дивергенция представляет собой единственный исторический процесс, имеющий место в эволюции надындивидуальных групп организмов в природе, исключая различие в темпах эволюции. Приемы выяснения филогенетических отношений и выработка соответствующей терминологии кладизма представляются весьма ценными, принципы же классификации, напротив, вызывают лишь недоумение. Так в соответствии с принципами, сформулированными Хённигом, пользоваться старой классификацией нельзя, так как и таксоны, и их ранги он понимает совсем иначе. Только вид остается в качестве элементарной единицы систематики (19). Одним из центральных понятий традиционной систематики является монофилия. Монофилетическим обычно именуется таксон, возникший единым корнем от своего предка, т. е. такой, нижнюю границу которого пересекает единственная различимая линия предка. При этом вид, к которому сходятся все филогенетические линии в пределах монофилетического таксона, по своим признакам также является его членом. Хённиг же сузил это понятие, связав его с определенными особенностями не только происхождения таксона, но и с его дальнейшей судьбой: его монофилетический таксон также возникает единым корнем, но сверх того включает всех потомков предкового вида (22). Группы, монофилетические в обычном смысле, но имеющие потомков, не включаемых в их состав, Хённиг назвал парафилетическими и отказал им в статусе таксона. Например, пресмыкающиеся вместе с птицами и млекопитающими представляют собой монофилетическую группу, но и те и другие произошли от пресмыкающихся, следовательно, по Хённигу, класс пресмыкающихся – парафилетический и не может быть таксоном в кладистической системе. Таксоном в ней может быть только группа без потомков: амниоты как целое, динозавры вместе с птицами, тероморфы с млекопитающими и т. д. Чтобы сохранить до тех пор общепринятое исходное содержание термина «монофилия», сторонники традиционного направления хённиговскую монофилию называют голофилией. Метод синапоморфий служит для выяснения филогенетических отношений. Метод этот заключается в восстановлении филогении на уровне кладограммы (древовидной схемы) с помощью анализа судеб эволюционных новообразований – апоморфий. Если новообразование возникло перед каким-либо расщеплением эволюционной линии, но после предыдущего, в идеале его можно обнаружить у всех потомков того вида, у которого оно появилось. Его можно использовать как характеристику группы, объединяющей предка со всеми его потомками и отличающую этим от других групп. Такая характеристика квалифицируется кладистами как аутапоморфия. Однако если какое-либо свойство объединяет несколько групп, его и называют синапоморфией. Например, втяжные когти– синапоморфия представителей семейства кошачьих, одновременно это и его аутапоморфия, так как отличает семейство от других хищников. Филогения по Хённигу описывается как судьба линий признаков: в определенный момент каждая линия переходит из исходного состояния – плезиоморфного – в производное, или апоморфное; скачок совершается в точке дивергенции – узле кладограммы, в следующем узле хотя бы одна линия должна претерпеть еще одно такое же превращение. Между узлами ничего не происходит, лишь определяется интервал между событиями в узлах. Стягивание всех эволюционных событий в узел кладограммы Хённиг допускает только как формальный прием анализа при ее построении. Общие признаки, наблюдаемые у ряда представителей, по Хённигу, могут быть синапоморфиями, возникшими у их ближайшего общего предка, и тогда это монофилетическая группа. Если же признаки достались от более далеких предков – это симплезиоморфии, они не свидетельствуют о непосредственном родстве связанных ими групп. Действительно, наличие двух пар ног у разных ящериц не доказывает их филогенетического единства как особой группы, противопоставленной змеям. Такие признаки ничего кладистике не дают (16). Одним из постулатов Хённиг является то, что в каждой точке расщепления возникают две разные апоморфии, т, е. обязательным считается наличие своей апоморфии (следа расщепления) в образовавшихся производных линиях. Исходя из этого, вид и любой другой таксон, разделяясь на два, прекращает свое существование, обе сестринские ветви от него должны отличаться, вид-предок не может сосуществовать с видом-потомком, и любые две разошедшиеся ветви маркированы апоморфиями с момента возникновения. С этим утверждением также нельзя согласиться, так как неизвестен ни один биологический механизм, который запрещал бы изменение только части популяций вида. Этот постулат нужен кладистам, чтобы допустить возможность однозначно определять все узлы кладограммы (25). Легко представить широко распространенный континентальный вид, который в разное время проник на некоторые острова и там обособился в ряд новых видов, но остался стабильным на континенте, т. е. сохранил свои признаки в плезиоморфном состоянии. Такой исход дивергенции скорее правило, чем исключение. Построение кладограмм (сборка) идет сверху вниз – от вершин к корням: сначала минимальные группы (виды, роды) объединяются в сестринские пары по самым узким синапоморфиям, затем две пары или пара и одиночка – по более широким и т. д. до завершения операции. Сестринской парой считается та, в которой найдена хотя бы одна синапоморфия. Поскольку реконструкция порядка ветвления филогенетических линий производится по апоморфиям (производным состояниям отдельных меняющихся признаков), определить, какое состояние признака было сначала и какое потом, становится важной проблемой, ибо признаки могут развиваться как в прямом, так и в обратном направлении. Если в группе признак выражен более чем в двух вариантах, их располагают в ряд так, чтобы образовался наиболее постепенный переход между крайностями. Эти ряды называют морфологическими, а также трансформационными сериями или морфоклинами. В многочисленных рядах, как матрешки, одна в другой содержатся апоморфии, последовательно маркирующие узлы кладограммы все более высокого порядка; чтобы выявить каждый отдельный узел, ряд последовательно делят на две части. Направление эволюции в таких рядах, или полярность ряда, может определяться по-разному. Кладисты считают наиболее строгим метод внешней группы, под которой подразумевается ближайшая к изучаемой родственная группа, т. е. сестринская. Так как из двух состояний признака то, которое встречается в обеих группах (если внешняя определена правильно), должно было возникнуть у их общего предка, оно в изучаемой группе считается исходным, плезиоморфным, а то, которое обнаруживается только у части этой группы, возникло в ее пределах, стало быть, позже,– апоморфным. Другие приемы распознавания полярности трансформационных серий основываются на палеонтологических свидетельствах – более древние находки считаются носителями плезиоморфных признаков (геологическая последовательность); на выводах из биогенетического закона – плезиоморфные состояния раньше возникают в онтогенезе (онтогенетическая последовательность); на географическом распределении носителей признаков – плезиоморфны те их состояния, которые встречаются только в центре происхождения группы (хорологическая последовательность). Кроме того, если в некоторой группе полярность каких-то морфоклин считается хорошо обоснованной, в других рядах можно с известной осторожностью принять ту же полярность. Надежность всех этих приемов не бесспорна, а некоторые из них даже весьма шатки. Строящаяся с их помощью филогенетическая схема всегда предварительна и часто неполна, поскольку отношения не всех анализируемых групп выяснены с приемлемой надежностью. Об этом нельзя забывать и тем более скрывать. Но у кладистов филогения – в основе системы, фактически сама система, а она должна быть максимально полной. Это понуждает их детализировать кладограмму даже там, где нет достаточных оснований, и порой удовлетворяться указанием единственной, пусть даже сомнительной синапоморфии на узел кладограммы, что означает столь же слабое обоснование таксона. Конечно, поспешностью в детализации кладограммы грешат отнюдь не только кладисты, но только у них это прямо сказывается на системе, обрекая ее на обилие слабо обоснованных таксонов и, соответственно, на значительную нестабильность. Для «традиционалистов» основа системы – отношения сходства между таксонами, филогения же используется для контроля за его надежностью. Если сходство очерчивает полифилетическую группу, скорее всего, либо оно поверхностно, либо полифилия кажущаяся – в любом случае это сигнал о необходимости дополнительного анализа системы и филогенеза группы. В остальном филогения для обычных систематиков имеет самостоятельное значение, а кладограмма выступает как канва и иллюстративный материал, наглядное отображение процесса. Главную же познавательную ценность имеет словесное описание эволюционного процесса с анализом его времени, условий, обстоятельств, движущих сил, модифицирующих факторов, закономерностей, последовательности крупных и мелких событий – словом, всего того, что с легкой руки кладистов уже привычно обозначается словом «сценарий». Кладистический подход к системе, в отличие от традиционного, предполагает, как уже говорилось, ее вторичность по отношению к филогении и конкретно к кладограмме. Это значит, что только дивергенция, отмеченная синапоморфиями, служит законным основанием для выделения таксона. Такой таксон будет голофилетичным, т. е. будет иметь единственную линию предков, как в традиционной системе, но в отличие от ее таксона кладистический не может иметь потомков – иначе он окажется парафилетическим, а этого кладисты не признают. Поэтому человека нельзя выделить из человекообразных обезьян в особое семейство, ибо тогда последние станут парафилетической группой. Точно так же птиц нельзя выделить из динозавров, млекопитающих – из тероморфных рептилий, амниот – из амфибий, амфибий – из рыб и т. д. Кладистическая система точно описывает родственные отношения, но не отражает баланс сходств и различий, вернее, делает это, исходя из равномерности эволюции, т. е. равномерного накопления различий. Своеобразием отличается также кладистический подход к установлению ранга таксонов. В традиционной системе ранг таксона (его соответствие уровню, скажем, вида, семейства или класса) определяется масштабом его отличий или, точнее, степенью обособленности как некой интегральной меры его сходства и родства с другими таксонами. Кладисты строят системы в конечном счете только по дивергенциям, которые все оказываются равноценными, так что единственным собственным критерием ранжирования таксонов остается последовательность дивергенции. Но если каждая следующая дивергенция дает таксон более низкого ранга, чем предыдущая, то число рангов будет огромным, система названий этих рангов (вид, род и т. д.) потеряет смысл и таксоны придется просто нумеровать, как это и делал сам Хенниг. Непрактичность подобной системы рангов столь очевидна, что кладисты сами от нее отказались и используют все ту же традиционную, совершенно не связанную с их принципами. Добавим к сказанному то, что уже обсуждалось: кладисты строят систему, опираясь только на родственные отношения, которые всегда известны хуже, чем отношения по сходству. Действительно, в обоих случаях важно, как распределены признаки сходства и различия, но кладисты могут использовать только те признаки, о которых известно, исходные они или производные. Это сужает эмпирическую базу кладистической системы и, значит, ее обоснованность. Кроме того, кладисты считают важной лишь последовательность дивергенции, все остальное, весь сценарий, хотя и полезен для их построений, но необязателен. Напомним также, что кладистика, именуемая первоначально филогенетической систематикой, менее всего приспособлена для анализа самого важного филогенетического материала – палеонтологического, поскольку, во-первых, не признает предковых таксонов, во-вторых, из-за их меньшей, чем современные родичи, изученности редко может включить их в общую кладограмму на равных основаниях. Таким недостаточно изученным палеонтологическим группам кладисты придумали даже специальную категорию неполноценного (с их точки зрения) таксона – плезион. В филогенетике наиболее удобный путь визуального представления эволюционных взаимоотношений среди групп организмов осуществляется посредством графиков, которые называются филогенетичними древом (рис.3): 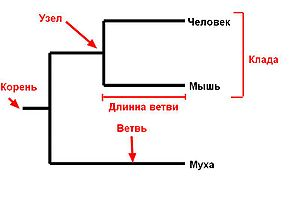 Рис.3. Филогенетическое древо.
Популярность кладистики основывается на привлекательной для научного сообщества возможности в явном виде, шаг за шагом описать в воспроизводимой форме процедуру построения кладограммы, а с ней и системы группы организмов, что недостатки кладистического подхода не хочется замечать. Забывается, что важнейшая задача системы – организация органического разнообразия в группы (таксоны), максимально однородные внутри себя и максимально различающиеся между собой (7). Нам представляется, что несомненные достоинства кладистики все же далеко не уравновешивают ее не менее очевидные недостатки. Классификацию, видимо, нельзя сводить к филогении. Классификация основывается на филогении, но не является ее простой и слепой проекцией. Это не освобождает, конечно, от необходимости освоить все полезное в кладистике, прежде всего, систему методов реконструкции филогенеза. 1.3.3.Фенетическая систематика В современной таксономической науке есть еще одно направление, классифицирующее животных - фенетическая систематика, идеологами которой были англичанин А.Кэйн и американцы Р.Сокэл и П.Снит. Это направление биологической систематики сформировалось в первой половине ХХ в., когда биологией на какое-то время завладела позитивистская философия познания (27). Будучи в теоретическом плане наследницей номинализма, она исходит из того, что объектами научного исследования могут быть только так или иначе наблюдаемые и измеряемые объекты. В случае систематики таковы, очевидно, сами организмы. Соответственно, всякие теоретизирования о том, как организовано и почему возникло биологическое разнообразие, избыточны и относятся к сфере метафизики. Последний тезис равносилен отказу от идеи естественной системы в ее эссенциалистском понимании. Вместо этого для оценки качества классификаций используются критерии, разработанные позитивистами в рамках своего понимания того, что такое научное знание. В фенетическом подходе такие классификации трактуются как особого рода справочные системы, свободные от какого-либо теоретического груза. С этой точки зрения, классификации должны быть: – устойчивы, т.е. минимально меняться при включении новых данных; – максимально прогностичны, т.е. позволять предсказывать максимально возможное число свойств организма, исходя из его положения в системе; – максимально прагматичны, т.е. обеспечивать эффективное решение возложенных на них практических задач, прежде всего хранения и извлечения информации (4). Следует заметить, что перечисленные критерии (иногда к ним добавляют еще два-три более частных) используются не только фенетической систематикой. К ним, как это ни странно, прибегают и теоретики современной типологии, для которой по-прежнему актуальна идея естественной системы. В методологическом плане фенетическая систематика базируется на «фенетической идее», которая означает, что организмы могут быть представлены суммой неких элементарных признаков, которым приписывается равный вес (14) . Достаточным основанием для классифицирования служит так называемое общее сходство – сумма всех «единичных сходств», получаемых по этим элементарным признакам. «Достаточность» в данном случае означает, что это сходство никак далее не интерпретируется – ни с точки зрения отражения сущностей, как в эссенциализме, ни с точки зрения родства, как в далее рассматриваемом эволюционном направлении. Соответственно, в данном случае таксон – это фенон, т.е. просто группа наиболее сходных между собой организмов. Методически фенетическая систематика ориентирована на разного рода количественные меры оценки сходства и количественные классификационные алгоритмы. Поэтому данную школу часто называют нумерической таксономией. Лежащая в ее основе идея с очевидностью вытекает из фенетической: коль скоро общее сходство, есть сумма единичных признаков, нужно просто уметь измерять и суммировать последние. Для этого разработана геометрическая интерпретация сходства. Она основана на концепции фенетического гиперпространства, осями которого являются признаки, причем как количественные, так и качественные (20). Сравниваемые объекты рассматриваются как точечные тела, распределенные в этом пространстве, а сходственные отношения – как непрерывные расстояния в нем, которые могут быть так или иначе измерены. Названная концепция находит широчайшее применение в самых разных биологических дисциплинах, где можно количественно оценивать сходство, – от биогеографии (районирование) до экологии (перекрывание ниш). В нумерической таксономии основным критерием пригодности того или иного метода нередко считается его математическая обоснованность, а не биологическая состоятельность. Исходя из этого, основное внимание уделяется разработке техники, а не биологической теории классифицирования (отчасти это продолжение схоластической традиции). Идеологами этого направления систематики являются уже упоминавшиеся выше Сокэл и Снит; в нашей стране его ярким представителем был энтомолог Е.С. Смирнов. Фенетическая систематика уделяет основное внимание разнообразию на низших таксономических уровнях – видовом и внутривидовом, обеспечивая техническими средствами «новую систематику». Немногие попытки применения этого подхода на более высоких уровнях чаще всего приводили к обескураживающим результатам. Причина в том, что при работе с морфологическим материалом на этих уровнях практически невозможно достичь идеала фенетического подхода – представить организм как совокупность равноценных элементарных признаков (22). Однако при обращении к макромолекулам фенетическая идея работает безотказно. Она позволяет сравнивать самые морфологически несходные организмы (например, прокариот, высших позвоночных животных) по нуклеотидным последовательностям, в которых каждый сайт (положение нуклеотида с определенным номером) считается отдельным элементарным признаком. В этом смысле современная геносистематика, работающая с такого рода данными, по своим методическим принципам является чисто фенетической (17). Впрочем, идеологически она ориентирована на филогенетику и рассматривает численную оценку суммарного сходства как показатель времени эволюционной дивергенции организмов. Обращение систематики к молекулярно-генетическим данным в конечном итоге вылилось в формирование геносистематики – одного из последних ответвлений эволюционного направления. В ней родственные отношения реконструируются по строению белка, ДНК, РНК, которым приписывается больший вес, чем данным классической морфологии. Этот подход базируется на теории нейтральной эволюции: разработанная ею концепция молекулярных часов позволяет выделять и ранжировать таксоны по количеству различающих их геномных перестроек, которые трактуются как мера времени их эволюционного расхождения. Геносистематика, имея дело с тысячами аминокислотных остатков или нуклеотидных оснований, целиком опирается на количественные методы оценки сходства и построения молекулярно-генетических деревьев (10). 1.3.4. Кладистическая геносистематика В настоящее время в систематике доминирует подход, который можно назвать «кладистической геносистематикой»: классификации разрабатываются на основе кладистических принципов, примененных к молекулярно-генетическим данным. Названную школу, в сравнении с традиционными, отличает высокая формализованность как в представлении исходных данных, так и в методах получения результатов. Это делает всю процедуру классифицирования достаточно прозрачной и в принципе воспроизводимой. Несомненным достоинством является возможность объединять в общей системе таксонов такие отдаленные организмы, как прокариоты, цветковые растение и позвоночные, на основании достаточно обоснованных суждений о генетическом родстве между ними (16). Серьезным недостатком рассматриваемого подхода является его редукционный характер (отчасти служащий необходимым условием его формализованности). Филогенез редуцируется до последовательности кладистических событий, филогенетическое единство – до генетического родства, организм – до нуклеотидных последовательностей, филогенетическое дерево редуцируется до кладограммы, а из естественной системы исключается вся информация, относящаяся к эволюционной (в самом широком смысле) специфике групп. |