
Isbn 978-5-7262-1226 нейроинформатика 2010
| Вид материала | Документы |
СодержаниеПостановка задачи и описание алгоритма |
- Isbn 978-5-7262-1226 нейроинформатика 2010, 142.85kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 136.25kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 9.21kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 89.17kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 124.83kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 86.68kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 101.88kb.
- Isbn 978-5-7262-1377 нейроинформатика 2011, 107.92kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 127.94kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 25.66kb.
ISBN 978-5-7262-1226-5. НЕЙРОИНФОРМАТИКА – 2010. Часть 1
И.В. КАБАНОВА, Р.Р. АЛИЕВ
Институт теоретической и экспериментальной биофизики РАН, Пущино
Московский физико-технический институт (государственный университет)
rubin@iteb.ru
АНАЛИЗ ТЕТА-РИТМА ЭЭГ В НОРМЕ И ПРИ ЭПИЛЕПСИИ
ПРИ ПОМОЩИ ВЕЙВЛЕТ-ПРЕОБРАЗОВАНИЙ
Рассматривается задача анализа электроэнцефалограмм при помощи вейвлет-преобразований. В частности, проведен анализ фазовых соотношений между сигналами тета-ритма гиппокампа и медиальной септальной области мозга в норме и при эпилепсии. В результате удалось установить, что в нормальном состоянии постоянно ведущий канал отсутствует и нет явно выделенных сигналов определенной частоты; перед окончанием острых судорог во всей полосе частот тета-ритма ведет сигнал из септума; при спонтанной судороге в тета-ритме постоянно присутствует сигнал определенной частоты (около 6,5 Гц) с высокой степенью когерентности, при этом ведет сигнал из гиппокампа.
Введение
Для изучения электрической активности мозга традиционно применяют электроэнцефалографию (ЭЭГ) – метод, позволяющий судить об активности различных структур мозга в норме и при патологии по квазипериодическим колебаниям электрического потенциала. Особый интерес представляют исследования электрической активности в гиппокампе – структуре мозга, играющей ключевую роль в процессах запоминания осознанной информации. Наиболее ярко выраженным ритмом гиппокампа является тета-ритм, т.е. колебания с частотой 4-12 Гц. Это наиболее интенсивный и стабильный сигнал, который может быть зарегистрирован в электроэнцефалограмме мозга человека и млекопитающих. Впервые тета-ритм был описан в ЭЭГ кролика в 1938 г. в работе Юнга (Jung) и Корнмюллера (Kornmuller) [1].
Как показали работы разных исследователей (Виноградова и др, 1975; Buzsaki et al., 1983; Leung, 1984; Stewart and Fox, 1990; Lee et al., 1994; Vinogradova, 1995; Vertes, Koscis, 1997; Кичигина и др., 1998; Виноградова и др., 2000; Buzsaki, 2002; Vertes, 2005; Buzsaki, 2005) [2], тета-ритм связан с двумя входами в гиппокамп от энторинальной коры и от медиальной септальной области (МС). Энторинальная кора образует ритмический возбуждающий вход, в то время как МС – ритмический тормозный вход.
Однако до настоящего времени, несмотря на усилия многих исследователей, вопрос о функциональной роли тета-ритма остается одним из самых сложных в нейрофизиологии и не теряет своей актуальности.
Колебания в нейронных системах, необходимые для осуществления когнитивных функций в здоровом мозге, при патологиях (в частности, при эпилепсии) могут взаимодействовать с нормальными мозговыми процессами [1, 2]. Так, при выяснении механизмов височной эпилепсии были обнаружены значительные морфологические, биохимические и электрофизиологические изменения в гиппокампально-энторинальной системе, на изучении которой и была сосредоточена большая часть исследований. В то же время не менее важным является выяснение роли второго основного входа в гиппокамп – от медиальной септальной области. Исходя из данных о значении септума в организации электрической активности гиппокампа, можно предположить, что эта область мозга играет определенную роль в процессе формирования патологического очага в гиппокампе при эпилепсии. Изучение значения септума в эпилептогенезе необходимо для более полного понимания механизмов данной патологии. Исследования в этом направлении могут привести к созданию новых подходов в терапии этого трудноизлечимого заболевания и, следовательно, являются чрезвычайно актуальными.
Традиционно для анализа ЭЭГ применяется преобразование Фурье. Однако применение данного метода не всегда эффективно, главным образом, из-за существенной нестационарности сигналов ЭЭГ. В настоящей работе, как и в ряде других работ [3, 4], предложено исследовать ЭЭГ при помощи вейвлет-преобразования, позволяющего проводить анализ нестационарных сигналов не только в частотном, но и во временном диапазонах.
Постановка задачи и описание алгоритма
Целью настоящей работы является использование вейвлет-преобразований для исследования особенностей тета-ритма электроэнцефалограммы септо-гиппокампальной системы мозга в норме и при эпилепсии. Акцент сделан на исследование фазовых соотношений между сигналами на частоте тета-ритма и выявление синфазных участков, а также ведущей (опережающей по фазе) области мозга с использованием знака разности фаз.
Вейвлет-преобразование одномерного сигнала состоит в его разложении по базису, сконструированному из обладающей определенными свойствами солитоноподобной функции (вейвлета) посредством масштабных изменений и переносов. Каждая из функций этого базиса характеризует как определенную пространственную (временную) частоту, так и ее локализацию в физическом пространстве (времени).
Таким образом, в отличие от традиционно применяемого для анализа сигналов преобразования Фурье, вейвлет-преобразование способно предоставить двумерную развертку исследуемого одномерного сигнала, при этом частота и координата рассматриваются как независимые переменные. В результате появляется возможность анализировать свойства сигнала одновременно в физическом (время, координата) и в частотном пространствах.
Рассмотрим пространство L²(R) функций f(t), определенных на всей действительной оси R(-∞,∞) и обладающих конечной энергией (нормой):
 . . | (0) |
Функции, образующие L²(R), должны стремиться к нулю на ±∞. Рассмотрим в качестве базисных функций вейвлеты – хорошо локализованные в пространстве солитоноподобные волны ψ(t).
Для данного класса функций введем систему сдвигов вдоль оси. В простейшем случае это может быть сдвиг на целое число единиц, т.е. преобразование вида ψ(t-k). Введём аналог синусоидальной частоты. Для определенности представим его в виде целочисленной степени числа 2. В этом случае мы получим класс функций: ψ(2jt-k), здесь j и k – целые числа. С помощью комбинаций дискретных масштабных преобразований (1/2j) и сдвигов (k/2j) можно получить любую частоту и покрыть всю действительную ось, имея единственный базисный вейвлет ψ(t). Таким образом, данные преобразования позволяют сформировать базис функционального пространства L²(R). Функцию ψ(t) часто называют материнским вейвлетом.
По определению нормы:
 | (0) |
(здесь звездочка обозначает комплексное сопряжение) получаем:
 , , | (0) |
т.е. если вейвлет ψ(t) L²(R) имеет единичную норму, то все вейвлеты семейства {ψjk} вида
 . . | (0) |
также нормированы на единицу, т.е. ||ψjk||2=||ψ||2=1.
Вейвлет ψ(t) L²(R) называется ортогональным, если семейство {ψjk} представляет собой ортонормированный базис функционального пространства L²(R), т.е.
 . . | (0) |
В таком случае каждая функция f
 L²(R) может быть представлена в виде ряда
L²(R) может быть представлена в виде ряда  , , | (0) |
равномерная сходимость которого в L²(R) означает, что
 . . | (0) |
Рассмотрим базис функционального пространства L²(R), полученный с помощью непрерывных масштабных преобразований и переносов вейвлета φ(t) с произвольными значениями базисных параметров – масштабного коэффициента a и параметра b:
 , a, b ϵ R, ψ ϵ L²(R). , a, b ϵ R, ψ ϵ L²(R). | (0) |
Данный базис позволяет определить интегральное вейвлет-преобразование:
 . . | (0) |
Аналогично преобразованию Фурье, коэффициенты cjk=
 . . | (0) |
Вместо [Wψf](a,b) для коэффициентов (амплитуд) вейвлет-преобразования можно использовать обозначения W(a,b) или Wψf, или W[f].
Итак, каждая функция из L²(R) может быть получена суперпозицией масштабных преобразований и сдвигов базисного вейвлета, т.е. является композицией «вейвлетных волн».
Для анализа сигналов хорошо подходит непрерывное вейвлет-преобразование. Его некоторая избыточность, связанная с непрерывным изменением масштабного коэффициента a и параметра сдвига b, позволяет более полно представить и проанализировать содержащуюся в данных информацию. В данной работе в качестве материнского вейвлета был выбран вейвлет Морле:
 . . | (0) |
Данный вейвлет является синусоидальной волной, модулированной кривой Гаусса с дисперсией
 . Параметр
. Параметр  называется центральной частотой.
называется центральной частотой.Особенностью вейвлета Морле, как и других комплексных вейвлетов, дающей дополнительные преимущества при анализе сигналов, является то, что Wx(a,b) можно записать в терминах фазы φx(a,b) и модуля ║Wx(a,b)║. При этом, по определению, локальная фаза вейвлет-преобразования выражается через отношение его мнимой (
 ) и действительной (
) и действительной ( ) частей следующим образом:
) частей следующим образом:  . . | (0) |
В этом случае появляется возможность выявить взаимосвязь сигналов при наличии временной задержки между ними с помощью вычисления разности фаз φx,y(a,b)= φx(a,b)- φy(a,b) или
| . | (0) |
Кроме этого, преобразование с использованием вейвлета Морле позволяет получить распределение разностей фаз, позволяющее судить о возможности синхронизации сигналов.
Исходные данные представляли собой фрагменты ЭЭГ, записанные с частотой дискретизации 6 кГц. Записи имеют вид wav-файлов, содержащих два канала, в которых хранятся последовательные величины потенциалов. Первый канал представляет собой сигнал из гиппокампа, второй – сигнал из септума.
В результате вейвлет-преобразования исходных данных на выходе получаются две матрицы комплексных коэффициентов размерности MxN, где N – длина векторов исходных данных, а M – длина вектора, отвечающего за полосу частот, в рамках которой производится вейвлет-преобразование.
После получения коэффициентов вейвлет-разложения производится вычисление различных характеристик вейвлет-преобразования: матриц амплитуд, матрицы разности фаз вейвлет-коэффициентов и матрицы значений вейвлет-когерентности. Полученные характеристики после статистической обработки, для более наглядного отображения результатов, были представлены в виде гистограмм (для разности фаз и амплитуд), графиков стандартного отклонения разности фаз, вейвлет-когерентности.
Результаты
С помощью вейвлет-преобразований была проанализирована электрическая активность мозга здорового животного, ЭЭГ при острых судорогах, ЭЭГ при спонтанных судорогах, ЭЭГ хронически больного животного.
Результаты вейвлет-преобразования сигнала электрической активности мозга здорового животного приведены на рис. 1. Представлены графики сигналов из двух областей мозга: гиппокампа и септума, амплитуды вейвлет-преобразований данных сигналов в диапазоне частот тета-ритма от 4 до 12 Гц и график разности фаз колебаний из этих двух областей.
На рис. 2. показано распределение амплитуд для гиппокампа и септума здорового животного. Видно, что ширина распределения значительно меньше в септуме, чем в гиппокампе.
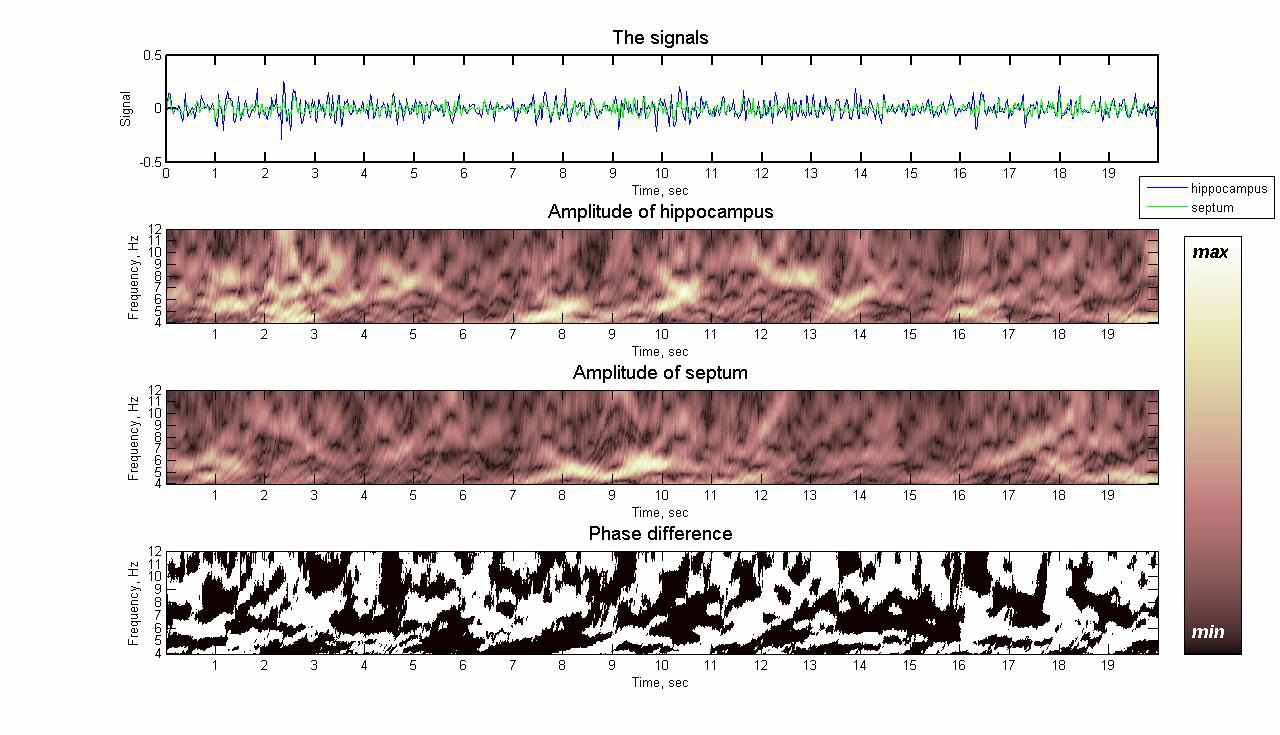
Рис. 1. Активность мозга здорового животного
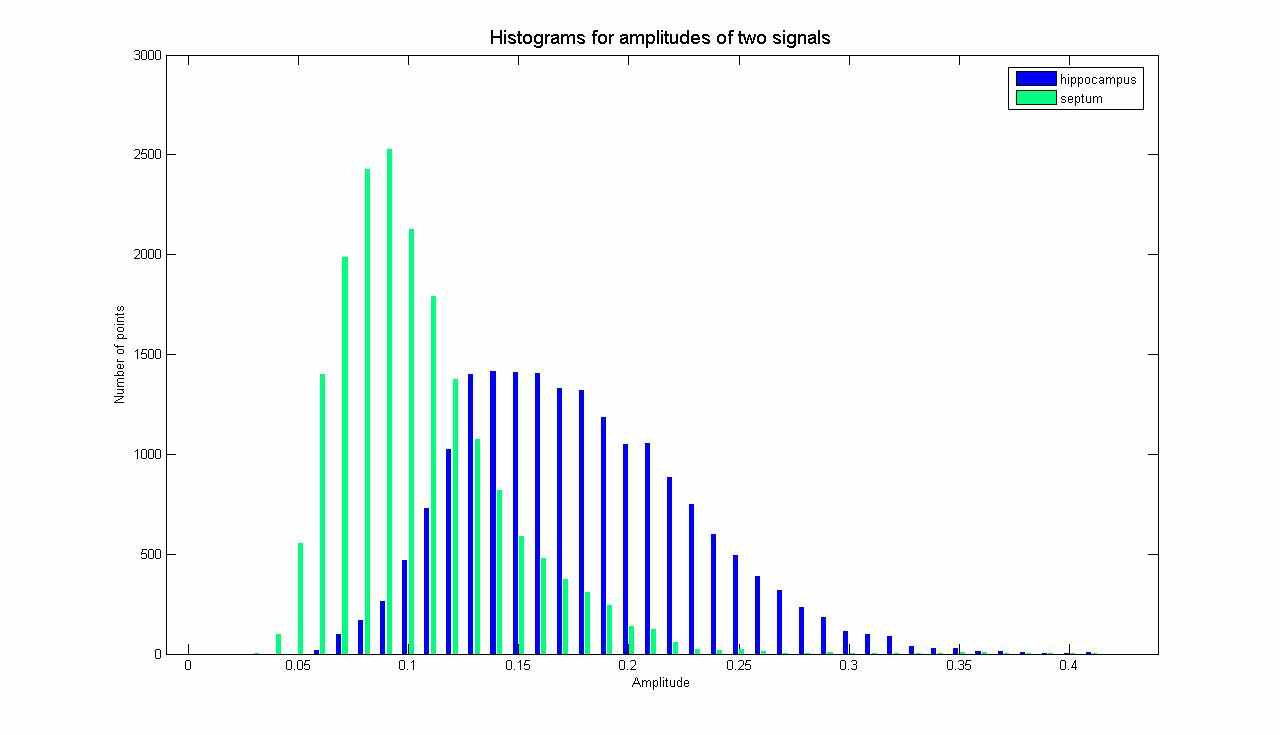
Рис. 2. Гистограммы распределения амплитуд вейвлет-коэффициентов
для гиппокампа и септума здорового животного
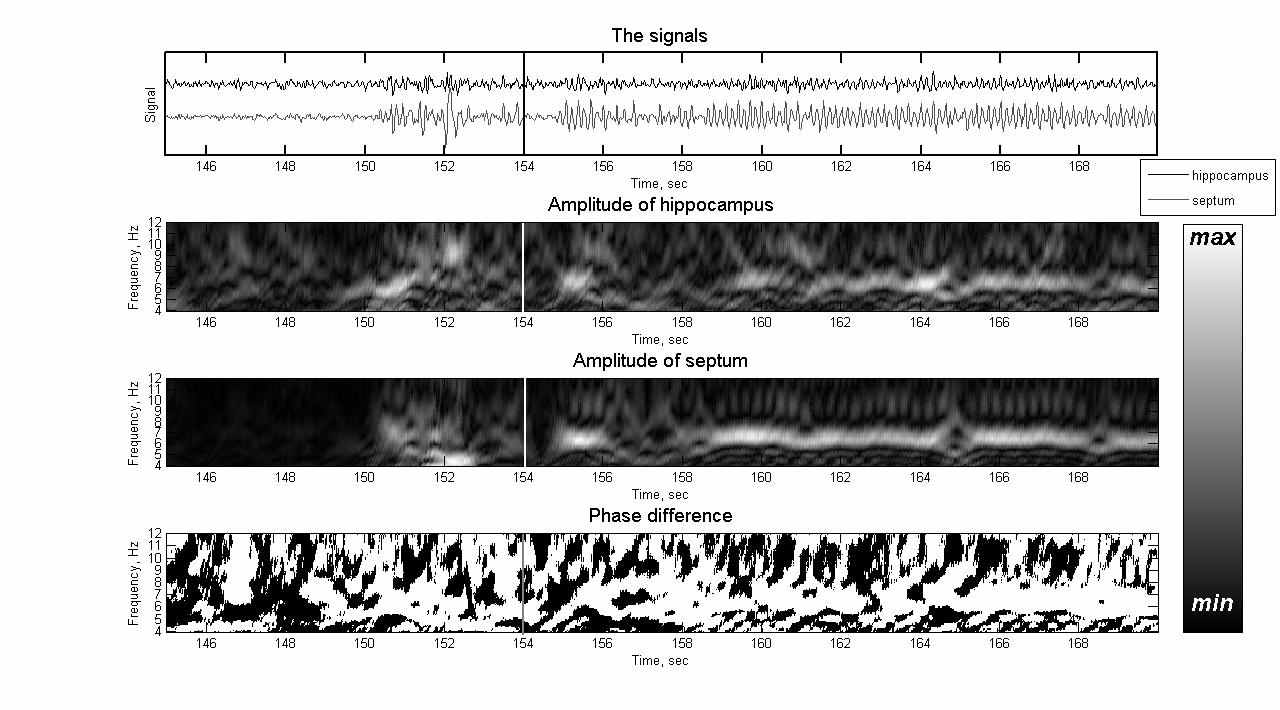
Рис. 3. Активность мозга при спонтанной судороге
(начало судороги отмечено чертой)
При спонтанной судороге (рис. 3), в отличие от ЭЭГ в норме (рис. 1), в полосе тета-ритма постоянно присутствует сигнал фиксированной частоты (около 6,5 Гц) с высокой степенью когерентности.
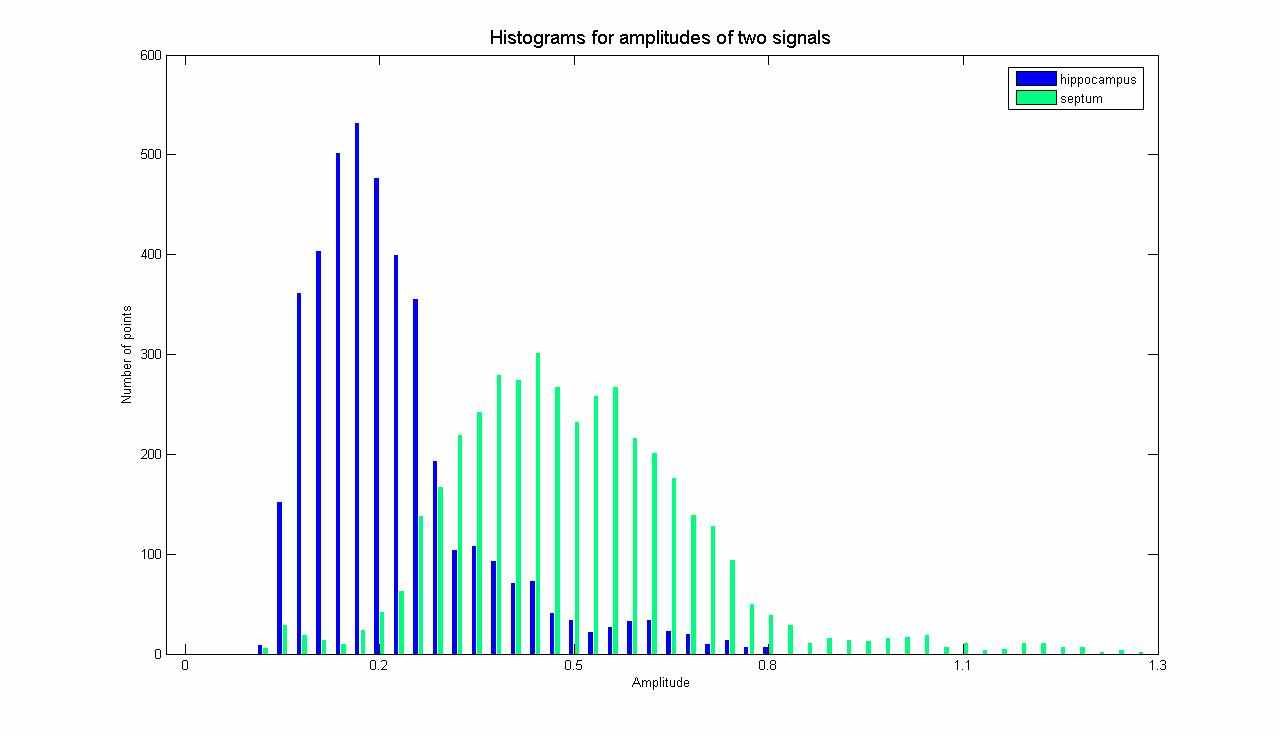
Рис. 4. Гистограммы распределения амплитуд вейвлет-коэффициентов
для гиппокампа и септума при спонтанной судороге
На рис. 4 показано распределение амплитуд для гиппокампа и септума при спонтанной судороге. Видно, что ширина распределения значительно меньше в гиппокампе, чем в септуме, что прямо противоположно ситуации для здорового животного (см. рис. 2).
Разность фаз сигналов до и во время спонтанной судороги показана на рис. 5. Видно, что до судороги, т.е. в норме, разность фаз в среднем нулевая, в то время как во время спонтанной судороги разность фаз становится положительной, т.е. сигнал из гиппокампа опережает по фазе сигнал септума.

Рис. 5. Гистограммы распределения разности фаз полученных в гиппокампе
и септуме во время и после спонтанной судороги на частоте 6,5 Гц
Выводы
Результаты проведенных исследований показывают, что вейвлет-преобразования являются современным эффективным методом анализа ЭЭГ. В частности, продемонстрирована возможность отслеживания изменений тета-ритма в гиппокампе и септуме с одновременным определением опережающего по фазе канала на основе анализа разности фаз вейвлет-преобразования сигналов.
В результате, с помощью вейвлет-преобразования при анализе экспериментальных записей ЭЭГ нормальных и больных животных нам удалось: (i) отличать стадии развития эпилепсии у животных; (ii) показать, что в нормальном состоянии постоянно ведущий канал отсутствует и в тета-ритме нет явно выделенных сигналов определенной частоты. Также отсутствуют участки длительной синхронизации каналов; (iii) перед окончанием острых судорог всегда присутствует отрезок времени, на котором во всей полосе частот тета-ритма ведет сигнал из септума; (iv) при спонтанной судороге в тета-ритме постоянно присутствует сигнал определенной частоты (около 6,5 Гц) с высокой степенью когерентности. При этом сигнал из гиппокампа опережает по фазе сигнал септума.
Список литературы
- Виноградова О.С. Гиппокамп и память / О.С. Виноградова. М.: Наука. 1975.
- Кичигина В.Ф. Механизмы регуляции и функциональное значение тета-осцилляций в септо-гиппокампальной системе мозга: Диссертация доктора биологических наук / ИТЭБ РАН – Пущино, 2006. – 207 с.
- Le Van Quyen M., Bragin A. Analysis of dynamic brain oscillations: methodological advances // Trends in Neurosciences. – 2007. – V.30. №7. P.365373.
- Голуб В.А. Выявление патологических паттернов ЭЭГ с помощью вейвлет-преобразования / В.А. Голуб, И.Н. Козлова, Н.П. Сереженко // Вестник ВГУ, Серия: Систем. анализ и информ. технологии. 2007. №2. С.6164.
Работа выполнена при частичной поддержке РФФИ, проект № 07-04-01187.
УДК 004.032.26(06) Нейронные сети