
Isbn 978-5-7262-1226 нейроинформатика 2010
| Вид материала | Документы |
СодержаниеПостановка задачи Подбор параметров задачи T = 3,5 мс (характеристическое время разряда мембраны); Δt |
- Isbn 978-5-7262-1226 нейроинформатика 2010, 142.85kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 136.25kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 9.21kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 113.94kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 89.17kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 124.83kb.
- Isbn 978-5-7262-1226 нейроинформатика 2010, 101.88kb.
- Isbn 978-5-7262-1377 нейроинформатика 2011, 107.92kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 127.94kb.
- Isbn 978-5-7262-1375 нейроинформатика 2011, 25.66kb.
ISBN 978-5-7262-1226-5. НЕЙРОИНФОРМАТИКА – 2010. Часть 1
А.Н. ПОКРОВСКИЙ
- Санкт-Петербургский государственный университет
- anpokr@petrodvorets.spb.ru
- ВЫДЕЛЕНИЕ СИНАПТИЧЕСКИХ ТОКОВ
- ИЗ ВЫЗВАННОГО ПОТЕНЦИАЛА
Предлагается выделять из вызванного потенциала коры мозга компоненты, соответствующие токам отдельных типов синапсов, методом подбора квазирешений модельной задачи. В качестве базовой информации наряду с записями вызванного потенциала используется кусочно-линейная аппроксимация формы тока синапсов. Задача сведена к подбору конечного числа неизвестных параметров при аппроксимации участков записи вызванного потенциала. Рассмотрены примеры.
Введение
Проблема разделения вызванных потенциалов на компоненты различного происхождения возникла с появлением записей вызванных потенциалов. В связи с этой проблемой изучались токи синапсов [1],[2] и послойное отведение вызванных потенциалов (ВП) коры мозга [3]. Позже стало использоваться синхронное послойное отведение ВП коры мозга системой электродов [4]. Вершиной экспериментальных исследований является работа [5], в которой зарегистрированы ВП колонки коры мозга, возникающие в ответ на импульсы одного-единственного нейрона таламуса. Но даже в этих записях внеклеточных потенциалов в начале ответа имеется наложение ВП и внеклеточных потенциалов от нервных импульсов в ветвях аксона, и разделить их только экспериментальными методами невозможно. Необходимо использовать теоретические методы [6], [7] и математические модели.
Главной трудностью в рассматриваемой задаче является то обстоятельство, что в задаче содержится много процессов и параметров, и не все они могут быть измерены в одном и том же эксперименте. Поэтому часть данных приходится постулировать или подбирать, опираясь на результаты других экспериментов, выполненных подчас при других условиях. Результат вычислений зависит от того, насколько удачен был выбор этих гипотез и параметров. Например, форма синаптических токов, вычисленных в [8] – [10], не совпадает с формой токов, регистрируемых в других экспериментах. В связи с этим ниже вводится дополнительная гипотеза о форме тока через синапсы, и вместо решения обратной задачи восстановления синаптического тока решается задача подбора конечного числа параметров. Это соответствует известному методу вычисления квазирешений обратной задачи [11].
Данная работа является развитием исследований [7-10].
Постановка задачи
Современный многоканальный внеклеточный электрод в коре мозга изображён на известной из литературы фотографии (рис. 1, слева). Ось этого электрода приблизительно параллельна направлению апикальных дендритов коры. Вызванный потенциал (рис. 1, справа) определяется в основном внеклеточными потенциалами апикальных дендритов.
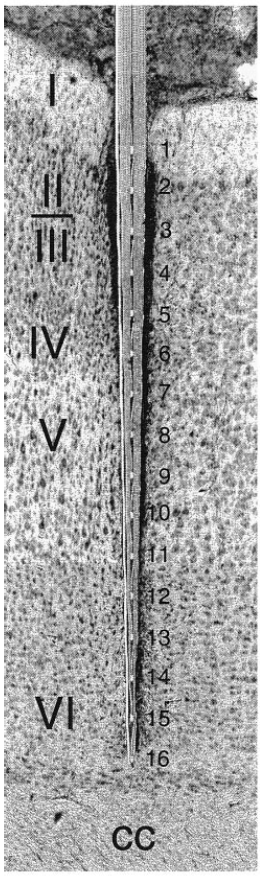

Рис. 1. Слева – многоканальный электрод в коре мозга. Справа – записи электродом такого типа внеклеточных потенциалов, возникающих от импульсов одного аксона ([5], Fig. 1B, N1). Время записи 5 мс
Далее формулируются и решаются следующие задачи:
1) Формулировка модели тока синапсов.
2) Формулировка математической модели внутриклеточных и внеклеточных потенциалов.
3) Алгоритм вычисления средних внеклеточных потенциалов апикальных дендритов при заданных токах в синапсах.
4) Подбор параметров задачи (в том числе латентности и амплитуды синаптических) токов для рассматриваемого экземпляра вызванного потенциала.
Токи через синапсы и их кусочно-линейная аппроксимация
Для формулировки модели тока синапсов рассмотрим данные из [1] (Fig. 7, B и Fig. 8, C) с записями токов возбуждающих и тормозящих синапсов при фиксации потенциала мембраны нейрона.

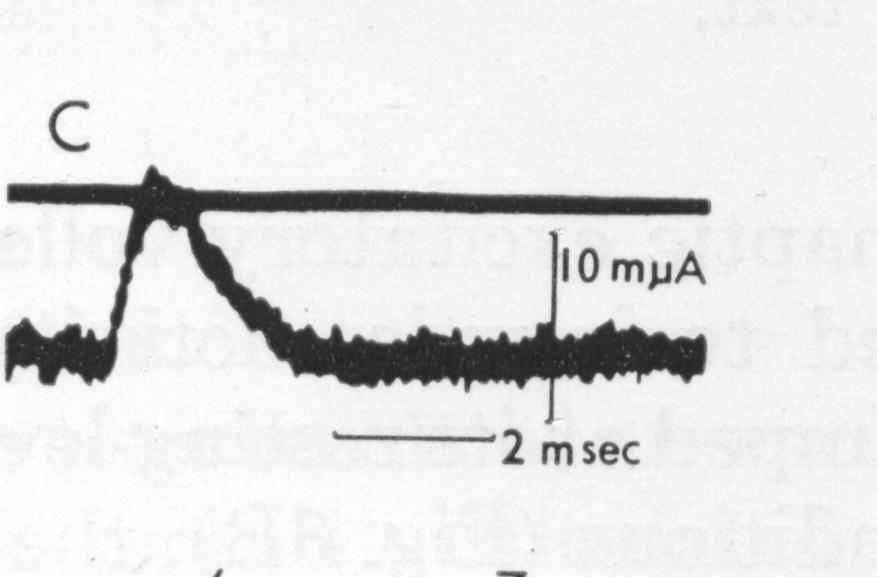


Рис. 2. Вверху – ток при фиксации потенциала, внизу – аппроксимация. B – возбуждающие синапсы, C – тормозящие синапсы
И
з рис. 2 следует, что кусочно-линейная аппроксимация токов синапсов может оказаться вполне удовлетворительной при моделировании компонентов вызванного потенциала коры.Рабочие гипотезы
1. Вызванные потенциалы создаются только апикальными дендритами.
2. Кора – плоская, все дендриты – одинаковые параллельные цилиндры, направлены поперёк коры, по оси Z, и распределены равномерно по плоскости коры.
3. Плотность каждого типа синапсов на дендритах зависит только от координаты z. Все синапсы каждого типа активируются одновременно.
4. Внутриклеточные потенциалы дендритов малы по сравнению с порогом спайков, и мембрана дендритов рассматривается как пассивный проводник
5. Предполагается, что все синапсы на дендритах расположены на тех же глубинах в коре, что и контакты электрода.

Обозначение переменных
u(z,t) – внутренний потенциал дендритов (в милливольтах);
φ(z,t) – средний внеклеточный потенциал в коре (в милливольтах);
Isyn(z,t) – синаптический ток через поверхность дендрита;
λ – характеристическая длина дендритов;
T – характеристическое время разряда мембраны;
g – характеристическая проводимость;
k, j – номер канала электрода; N – число каналов электрода.
Переход к безразмерным переменным
t – безразмерное время (в долях T);
z – безразмерная длина (в долях λ ); zk – безразмерная координата глубины k-того канала электрода.
ψ – приведенный синаптический ток.
Уравнения модели
(Заимствованы из [8], [9], [10])








Подбираемые параметры
T – характеристическое время разряда мембраны;
Δt – латентность синаптических токов (для всех уровней одинаковая);
Δz=zk+1 – zk – для всех уровней одинаковы;
K6 , K7 , K8 , K9 , K10 , K11 ,K12 – коэффициенты амплитуды синаптического тока.
Пример вычисления потенциала
Для кусочно-линейного возбуждающего приведенного тока, (как на рис. 2, слева) изображенного на рис. 3, слева, на рис. 3, справа построены вычисленные по формуле (8) потенциалы.


Рис. 3. Слева – ток, справа – вычисленные потенциалы. N=7, zk+1-zk=1, T=5 мс.
Время в миллисекундах, амплитуда в милливольтах
Подбор параметров задачи
Из потенциалов 16-ти уровней, изображенных на графике рис. 1, справа, для вычислений использовались только 7 средних (с 6-го по 12-й) уровней, на которых заметны ответы на импульс нейрона таламуса.
Для графиков на рис. 4 (справа) и рис. 5 были подобраны следующие значения параметров:
T = 3,5 мс (характеристическое время разряда мембраны);
Δt = 0,89 мс - латентность синаптических токов (для всех уровней одинаковая);
Δz = zk+1 – zk = 1,7 - для всех уровней одинаковы;
K6 = 0,09; K7 = 0,07; K8 = 0,5; K9 = 0,55; K10 = 0,32; K11 = 0,02;
K12 = 0,01; - коэффициенты амплитуды синаптического тока.
Результаты подбора параметров
Результат подбора синаптических токов:
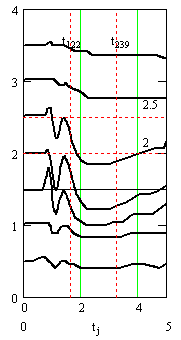
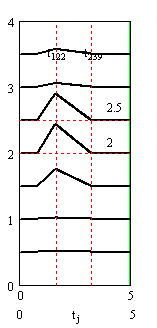
Рис. 4. Результаты подбора параметров и вычисления квазирешения обратной задачи. Слева – центральная часть (уровни с 6 по 12) записей вызванных потенциалов (из рис.1 справа) в увеличенном масштабе. На правом графике – вычисленные синаптические токи. Время в миллисекундах, потенциалы и токи в милливольтах
Вертикальными пунктирными прямыми на графиках рис. 4 и рис. 5 отмечены моменты максимума синаптических токов и время окончания синаптических токов.
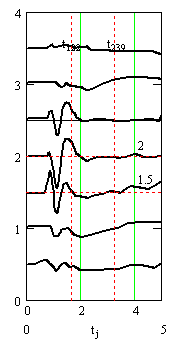
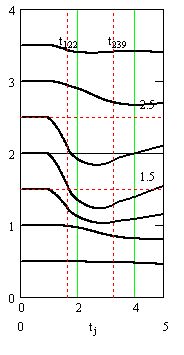
Рис. 5. Слева - внеклеточные потенциалы спайков в аксонах. Справа – график подобранного «чистого» вызванного потенциала. Сумма кривых на правом и левом графиках совпадает с соответствующими линиями графика на рис. 4, слева. Время в миллисекундах, потенциалы и токи в милливольтах
Вычисления (достаточно трудоёмкие) производились в системе Mathcad.
Выводы
Рассмотренный пример и вычисления с подбором констант показывают, что предлагаемый путь выделения компонентов из вызванных потенциалов заслуживает внимания. Любое уточнение модели приведёт к увеличению числа подбираемых констант.
Из высокой трудоёмкости данного метода вытекает желательность и необходимость автоматизации вычислений, то есть разработки системы специальных программ для анализа записей послойного отведения вызванных потенциалов.
Список литературы
Araki T., Terzuolo C.A.. Membrane currents in spinal motoneurons associated with the action potential and synaptic activity // J. Neurophysiology. 1962. V.25. No 6. P. 772-789.
Экклс Дж. Физиология синапсов. М.: Мир, 1966.
Andersen P., Eccles J. C., Loyning Y.. Recurrent inhibition in the hippocampus with identification of the inhibitory cell and its synapses // Nature (Lond.). 1963. V. 198. P. 541-542.
Покровский А.Н., Прохоренко Ю.П., Торнуев Ю.В. О происхождении раннего положительного компонента первичного ответа // Нейрофизиология. 1974. Т. 6. № 4. С. 455-458.
Swadlow H. A., Gusev A.G., Bezdudnaya T. Activation of a cortical column by a thalamocortical impulse // J. Neuroscience. 2002. No 22(17). P. 7766-7773.
Покровский А.Н. Основные задачи теории измерений электрической активности нейронных структур ЦНС // Материалы V симпозиума по вопросам нейрофизиологии. Вильнюс, 1967. С. 43-44.
Покровский А.Н. К вопросу о механизме возникновения электрических потенциалов в нервной ткани // Биофизика. 1970. Т. 15. № 5. С. 881-887.
Покровский А.Н. Моделирование внеклеточного потенциала в слое параллельных дендритов // Нейроинформатика-2006, VIII Всероссийская научно-техническая конференция, сборник научных трудов. 2006. Ч. 1. М.: МИФИ. С. 118-123.
Покровский А.Н. Уточнённая математическая модель внеклеточного потенциала в слое параллельных дендритов // Нейроинформатика-2007, IX Всероссийская научно-техническая конференция, сборник научных трудов. 2007. Ч. 2. М.: МИФИ. С. 25-31.
Покровский А.Н., Федоров А.С. Восстановление синаптических токов по вызванным потенциалам коры мозга // Нейроинформатика-2008, X Всероссийская научно-техническая конференция, сборник научных трудов. 2008. Ч. 1. М.: МИФИ. С. 126-132.
Тихонов А.Н., Арсенин В.Я. Методы решения некорректных задач. Гл. 1. М.: Наука, 1974.
Работа выполнена при частичной поддержке РФФИ, проект № 09-01-00473-а.
УДК 004.032.26(06) Нейронные сети