
Морфофункциональное состояние и свободнорадикальный гомеостаз щитовидной железы и надпочечников половозрелых и неполовозрелых самцов крыс при адаптации к периодическому охлаждению 03. 00. 13 физиология
| Вид материала | Автореферат |
- Влияние бишофитсодержащей рапы соленого озера «Малое Лиманское» Астраханской области, 306.45kb.
- Влияние прерывистой гипобарической гипоксии на морфофункциональные изменения щитовидной, 405.72kb.
- Современные методы лечения узловых образований щитовидной железы, 110.17kb.
- Урок №61 «Функции желез внутренней секреции», 38.42kb.
- Заболевания щитовидном железы, 94.48kb.
- Заболевания щитовидном железы (лекции), 219.78kb.
- Хирургическое лечение щитовидной железы, 32.35kb.
- Влияние сульфидно-иловой лечебной грязи на функциональные параметры и свободнорадикальный, 281.37kb.
- Локальная, системная и сочетанная лучевая терапия костных метастазов, 150.74kb.
- Внелёгочный туберкулёз Туберкулез внелегочный, 2730.21kb.
На правах рукописи
Яковенкова Людмила Александровна
МОРФОФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ И СВОБОДНОРАДИКАЛЬНЫЙ ГОМЕОСТАЗ ЩИТОВИДНОЙ ЖЕЛЕЗЫ И НАДПОЧЕЧНИКОВ ПОЛОВОЗРЕЛЫХ И НЕПОЛОВОЗРЕЛЫХ САМЦОВ КРЫС ПРИ АДАПТАЦИИ К ПЕРИОДИЧЕСКОМУ ОХЛАЖДЕНИЮ
03.00.13 – физиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Астрахань 2009
Работа выполнена на кафедре физиологии и морфологии человека и животных Государственного образовательного учреждения высшего профессионального образования «Астраханский государственный университет», г. Астрахань
Научный руководитель: Кандидат биологических наук
Ломтева Наталья Аркадьевна
Официальные оппоненты: Доктор медицинских наук, профессор
Удочкина Лариса Альбертовна
Доктор биологических наук, профессор
Джандарова Тамара Исмаиловна
Ведущая организация: Санкт-Петербургский государственный университет
Защита диссертации состоится 16 октября 2009 года в 12-30 часов на заседании объединенного диссертационного совета ДМ 212.009.01 при Астраханском государственном университете по адресу: 414000, г.Астрахань, пл.Шаумяна, 1
С диссертацией можно ознакомиться в научной библиотеке АГУ по адресу: 414000, г. Астрахань, пл. Шаумяна, 1
Автореферат разослан «______» сентября 2009 года.
Ученый секретарь
диссертационного совета,
доктор биологических наук Ю.В. Нестеров
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы: Холод является одним из естественных факторов среды, воздействующих на человека. Ведущая роль в процессах формирования адаптации организма к действию факторов среды, одним из которых является холод, принадлежит нейроэндокринной системе (Розен В.Б., 1984; Филаретов А.А., 1987; Држевецкая И.А., 1994; Божко А.П. и др., 1994; Hangalapura B.N. et al., 2004; Wan R. et al., 2004; Козырева Т.В., 2005; Баженов Ю.И. и др., 2006; Александрова Н.В. и др., 2006). Большинство работ посвящено изучению развития адаптационного звена в ответ на длительное воздействие стресс-индуцирующего фактора (Шмидт Ю.Д., 2003; Venditti P. et al., 2004; Козырева Т.В., 2005; Каленова Л.Ф. и др., 2005; Ахалая М.Я. и др., 2006; Maeda, T., 2007; Shevchuk N.A., 2007), тогда как многие параметры начинают изменяться с первых минут воздействия.
Установление факта мгновенного реагирования на действие стрессоров со стороны нейроэндокринной системы (Белякова Е.И, 2005; Белякова Е.И. и др., 2005, Поздняков О.Г. и др., 2005) делает актуальным изучение ранних изменений параметров функционального состояния желез внутренней секреции.
Адаптация к условиям среды, связанная с серией периодических воздействий стрессирующего фактора, распространена в окружающем мире гораздо шире, чем одномоментный перевод организма в новые условия на длительный срок (Le Blanc J., 1992; Bittel J., 1992). Постоянный интерес к данной проблеме и ее актуальность связаны с тем, что холод является одним из естественных факторов среды, воздействующих на человека.
При холодовой адаптации развитие несократительного термогенеза включает участие гормонов щитовидной железы, коры и мозгового слоя надпочечников. Гормоны щитовидной железы имеют большое значение при холодовой адаптации (Lauberg P. еt al., 2005), являясь основными модуляторами энергетического метаболизма и термогенеза (Bagchi N. et al., 1973; Frane L. et al., 1990; Jansen M. et al., 1994; Silvestri E. еt al., 2005). Молекулярной основой, обеспечивающей несократительный термогенез у адаптированных к холоду животных, является активация свободнорадикального окисления в митохондриях калоригенных органов и тканей (Venditti P., 2004). Холодовая адаптация повышает устойчивость к стрессам (Божко А.П. и др., 1994; Rintamäki H., 2006), при этом происходит стимуляция параметров антиокислительной защиты (Selman C. et al., 2000; Январева И.Н. и др., 2001; Ахалая М.Я. и др., 2006; Heise K. еt al., 2007; Sinha S. et al., 2009). Важная роль в становлении механизма холодовой адаптации отводится симпатоадреналовой и надпочечниковой системам (Козырева Т.В., 2005).
Адаптационные возможности различаются у животных разного возраста, так как в процессе онтогенеза происходит формирование и становление механизмов адаптации (Bozhkov A.I. et al., 2006; Ravi Kiran T. et al., 2006), однако возрастной аспект адаптации представлен в литературе недостаточно.
Необходим комплексный подход к изучению ранней ответной реакции со стороны щитовидной железы и надпочечников у животных разных возрастных групп в условиях адаптации к периодическому охлаждению.
Цель исследования – проанализировать морфофункциональное состояние и свободнорадикальный гомеостаз щитовидной железы и надпочечников половозрелых и неполовозрелых самцов крыс на ранних этапах холодового воздействия в условиях адаптации к периодическому охлаждению.
Задачи исследования:
- Определить уровень тироксина, трийодтиронина и кортикостерона в плазме крови половозрелых и неполовозрелых самцов крыс на ранних этапах холодового воздействия в условиях адаптации к периодическому охлаждению и сопоставить результаты с динамикой уровня свободнорадикальных продуктов и антиоксидантных ферментов;
- Проследить динамику морфофункционального состояния щитовидной железы и надпочечников самцов крыс на ранних этапах холодового воздействия в условиях адаптации и выявить возрастные особенности гистоструктурных перестроек эндокринных органов;
- Исследовать динамику уровня малонового диальдегида, скорости спонтанного и аскорбатзависимого перекисного окисления липидов в щитовидной железе и надпочечниках на ранних этапах холодового воздействия у половозрелых и неполовозрелых самцов крыс при адаптации к периодическому охлаждению;
- Выявить динамику каталазной активности в щитовидной железе и надпочечниках половозрелых и неполовозрелых самцов крыс на ранних этапах холодового воздействия в условиях адаптации к периодическому охлаждению.
Научная новизна. В работе впервые исследована динамика свободнорадикальных процессов и морфофункционального состояния щитовидной железы и надпочечников в течение 15-ти, 30, 45, 60 и 90 минут холодового воздействия у самцов крыс разного возраста. Выявлены особенности морфофункционального состояния щитовидной железы у животных, адаптированных к условиям холодового воздействия и динамики этих изменений при остром охлаждении. Впервые обнаружено, что активация морфофункционального состояния щитовидной железы в ответ на холодовое воздействие зависит от его длительности и различается у адаптированных к периодическому воздействию самцов крыс более ранним проявлением.
Показано, что холодовое воздействие приводит к разнонаправленным изменениям интенсивности процессов перекисного окисления липидов: у адаптированных животных происходит интенсификация, у неадаптированных - снижение уровня липопероксидации как в ткани щитовидной железы, так и в ткани надпочечников независимо от возраста.
Впервые установлено, что морфофункциональное состояние надпочечников отличается у адаптированных и неадаптированных к периодическому охлаждению животных. Показано, что у адаптированных самцов крыс обеих возрастных групп реакция коры надпочечников на острое холодовое воздействие снижена.
Впервые обнаружены возрастные закономерности изменения уровня гормонов щитовидной железы и надпочечников в динамике холодового воздействия в условиях адаптации. Выявлено, что ответная реакция и щитовидной железы и надпочечников начинается в ранний период холодового воздействия (после 15 мин охлаждения), тогда как снижение уровня гормонов щитовидной железы в крови в результате их поступления в ткани-мишени происходит раньше, чем гормонов надпочечников.
Основные положения, выносимые на защиту
1. Ранняя реакция щитовидной железы и надпочечников на острое охлаждение заключается в изменении концентрации гормонов в крови и в морфологических изменениях этих желез, которые обнаруживаются уже на 15 минуте воздействия;
2. В результате 20-ти дневного трехчасового периодического охлаждения самцов крыс формируются компенсаторно-приспособительные механизмы функционирования щитовидной железы и надпочечников, которые выражаются в гистоструктурной перестройке органов и изменении уровня кортикостерона в крови;
3. Адаптационная стратегия имеет ряд принципиальных различий в зависимости от возраста: неспецифические компоненты ответной реакции преобладают у неполовозрелых самцов крыс, а в реакции половозрелых животных на периодическое охлаждение проявляются специфические (тиреоидные, свободнорадикальные, антиоксидантные) механизмы.
Теоретическая и практическая значимость. Результаты исследований позволяют углубить существующие представления о физиологических механизмах регуляции и временной динамике адаптивных процессов при стрессе в зависимости от возраста. Полученные данные углубляют представления о формировании компенсаторно-приспособительных механизмов при периодическом охлаждении, важны для понимания роли щитовидной железы и надпочечников в регуляции адаптационных процессов. Теоретическая значимость работы состоит в обнаружении существенных различий динамики ответной реакции организма на действие острого стресса у адаптированных и неадаптированных животных.
Материалы исследований используются в учебном процессе при чтении лекционных курсов «Физиология человека и животных», «Биохимия», «Экологическая физиология», «Экология человека» в Астраханском государственном университете.
Апробация работы. Результаты исследования доложены и обсуждены на итоговой научной конференции АГПУ им. С.М. Кирова (Астрахань, 1999); VII итоговой научной конференции молодых ученых и студентов СГМА (Ставрополь, 1999); Международной научной конференции «Свободные радикалы, антиоксиданты и старение», посвященной 75-летию академика РАЕН, д.б.н., профессора Д.Л. Теплого (Астрахань, 2006); VIII и IX Международных научных конференциях «Эколого-биологические проблемы бассейна Каспийского моря» (Астрахань, 2005; 2006); XVI Международной конференции студентов, аспирантов и молодых ученых «Ломоносов» (Москва, 2009); VI Сибирском физиологическом съезде (Барнаул, 2008); конференции молодых ученых «Биомедицинские технологии, мехатроника и робототехника». По теме диссертации опубликовано 14 работ, в т.ч. 3 из них – в изданиях, рекомендованных ВАК РФ.
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, материалов и методов исследования и обсуждения результатов собственных исследований, заключения и выводов. Общий объем диссертации – 162 страницы, 30 таблиц и 26 рисунков. Библиографический список включает 216 источников, из них 95 – иностранных.
Материал и методы исследования
Эксперименты выполнены на 175 беспородных белых крысах самцах двух возрастных групп: взрослых половозрелых массой 200±15 г., возраст которых – 16 ± 2 недель и неполовозрелых 5-ти недельных крысах массой 50±7 г. Крысы содержались в стандартных условиях вивария при естественном освещении и свободном доступе к воде и пище. Все эксперименты соответствуют этическим нормам, изложенным в Женевской конвенции (1971), «Международным рекомендациям по проведению медико-биологических исследований с использованием животных» (1985). Все опыты проводились после введения животным этаминала натрия в дозе 4 мг/100 г массы тела.
Холодовой стресс индуцировали помещением животных в холодильную камеру при температуре +4ºС (Кормильцина Н.К., 1990; Линчевская А.А. и др., 1986). Для этого крыс в индивидуальных клетках, не ограничивающих их подвижность, помещали в холодильную камеру.
Проведены четыре серии экспериментов. В первой и второй сериях участвовали неадаптированные крысы (половозрелые и неполовозрелые). В третьей и четвертой сериях – животные, адаптированные к условиям холода (половозрелые и неполовозрелые).
В первой и второй сериях экспериментов (неадаптированные самцы крыс) животные однократно подвергались холодовому воздействию. Длительность экспозиции составляла 15, 30, 45, 60 и 90 мин соответственно. Одна из групп служила контролем. Таким образом, все животные были разделены на 6 групп.
В третьей и четвертой экспериментальных сериях холодовому воздействию подвергались самцы крыс, предварительно адаптированные к условиям охлаждения. Для адаптации животные подвергались ежедневной холодовой экспозиции при температуре +4°С в течение 3 часов на протяжении 20 дней (Колосова Н.Г. и др., 1999). Перед декапитацией самцы крыс однократно охлаждались 15, 30, 45, 60 и 90 мин соответственно, при этом, в зависимости от длительности воздействия были сформированы 5 экспериментальных групп. 2 группы животных служили контролем. Таким образом, в этих сериях было сформировано 7 групп животных. Первая группа – контроль неадаптированный – интактные животные. Вторая группа – контроль адаптированный – животные подвергались холодовой экспозиции ежедневно при температуре + 4°С в течение 3 часов на протяжении 20 дней. На 21 сутки крыс декапитировали, не охлаждая. Третья-седьмая группы – животные подвергались холодовой экспозиции при тех же условиях. На 21-е сутки крыс охлаждали 15, 30, 45, 60 и 90 мин каждую группу соответственно.
Перед исследованием функционального состояния щитовидной железы и надпочечников определяли динамику свободнорадикальных показателей на уровне целостного организма. Для этого изучали степень перекисного гемолиза эритроцитов и уровень перекисного окисления липидов в печени.
Критериями оценки функционального состояния щитовидной железы (ЩЖ) и надпочечников были: уровень трийодтиронина (Т3), тироксина (Т4), кортикостерона (КС) в плазме крови; 2) относительная масса щитовидной железы и надпочечников; 3) морфометрические показатели секреторного эпителия надпочечников и щитовидной железы; 4) уровень перекисного окисления (ПОЛ) липидов щитовидной железы и надпочечников; 5) каталазная активность в щитовидной железе и надпочечниках.
Морфологические и гистологические методы исследования щитовидной железы и надпочечников. После декапитации органы фиксировали в течение 3 суток при +4°С в 10 % растворе формалина на фосфатном буфере (рН 7,4). После соответствующей проводки через спирты возрастающей крепости органы заливали в парафин с использованием в качестве промежуточной среды бензола. Срезы толщиной 6 мкм, ориентированные по длине железы (срединная часть органа), окрашивали гематоксилином – эозином.
При морфометрическом анализе щитовидной железы измеряли высоту тироцитов (Ht), площадь ядер этих клеток (Sn), площади фолликулов (Sf), и фолликулярной полости (Sk).
В ткани надпочечников определяли: толщину коры надпочечников, толщину клубочковой, пучковой и сетчатой зон коры надпочечников, объем кортикоцитов и объем ядер кортикоцитов клубочковой, пучковой и сетчатой зон коры надпочечников. Морфометрию проводили, используя цифровые фотографии, получаемые с помощью цифрового фотоаппарата «Canon zoom lens» 3 X Power Shot A 95, MegaPixels 5.0 и микроскопа «Ломо» при увеличении 10х20, 10х40, монокулярного оптического адаптера и лицензированых компьютерных программ анализа и редактирования изображений. Число измеренных клеток – 50, число измеренных ядер – 100 от каждого животного. Величину изучаемых показателей выражали в мкм.
Интенсивность процессов перекисного окисления липидов (ПОЛ) определяли по уровню малонового диальдегида (МДА), скорости спонтанного (спПОЛ) и аскорбатзависимого (аскПОЛ) (Стальная И.Д. и др., 1977) в гомогенатах тканей щитовидной железы и надпочечников. Содержание МДА выражали в нмоль МДА/0,05 г сырого веса щитовидной железы и надпочечников, кинетические параметры – в нмоль МДА/час.
Иммуноферментным методом определяли уровень тироксина, трийодтиронина и кортикостерона в плазме крови. Определяли общую фракцию гормонов в плазме крови, результаты выражали в нмоль/л плазмы.
Активность каталазы оценивалась по методу М.А. Королюка, Л.И. Иванова (1988). Статистическая обработка полученных в ходе эксперимента данных проводилась на компьютере с использованием программ Excel 2000, Statistica 6,0. При обработке полученных данных использованы следующие показатели: расчет относительных показателей, определение средних величин, достоверности по критерию Стьюдента и коэффициента корреляции.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Динамика свободнорадикальных показателей на уровне целостного организма. Холодовое воздействие приводило к снижению степени ПГЭ. У половозрелых животных достоверно значимое снижение происходило к 90 мин холодового воздействия, у неполовозрелых с 15 по 45 мин. ПОЛ в печени под воздействием охлаждения изменялось незначительно. Происходило возрастание скорости аскПОЛ у неполовозрелых самцов крыс к 90 мин холодового воздействия, тогда как остальные показатели ПОЛ не изменялись.
Динамика уровня Т3, Т4 и КС. У неадаптированных неполовозрелых животных уровень тиреоидных гормонов при охлаждении практически не изменялся, уровень КС снижался с 45 по 90 мин холодового воздействия (Рис.1) с 1,43±0,290 до 0,70±0,08.
У адаптированных самцов крыс в контроле уровень КС у неполовозрелых животных был ниже, а у половозрелых выше в сравнении с интактными самцами. При холодовом воздействии у адаптированных половозрелых самцов крыс уровень Т4 (Табл.1) снижался на 45 и 90 мин охлаждения, уровень Т3 практически не изменялся (1,02±0,10). У адаптированных половозрелых животных холодовое воздействие приводило к возрастанию уровня КС (Табл.1) на 15 мин экспозиции (42,01±1,49), и это значение было максимальным в течение 90 минут исследования. При продолжающемся охлаждении уровень КС снижался, начиная с 30 мин и до конца воздействия. У адаптированных неполовозрелых животных холодовое воздействие приводило к увеличению уровня Т3 (Табл.1) с 15 мин по сравнению с контрольными самцами, оставаясь высоким до конца охлаждения; уровень Т4, напротив, был ниже относительно контрольной группы на всем протяжении холодового воздействия. Уровень КС у адаптированных неполовозрелых самцов крыс повышался с 15 по 30 мин холодового воздействия по сравнению с контролем, тогда как к 45 мин охлаждения уровень КС снижался (12,8±0,63).
Таблица 1
Изменение уровня Т3, Т4 и КС у адаптированных самцов крыс
| Горм. | | КНА | КА | 15 мин | 30 мин | 45 мин | 60 мин | 90 мин |
| Т 3 нмоль/л | ПЛ. | 0,89±0,04 | 1,02±0,10 | 1,10±0,07 | 1,03±0,08 | 1,02±0,10 | 0,97±0,06 | 0,97±0,10 |
| НЛ. | 1,41±0,17 | 0,95±0,06 • | 1,44±0,10 ** | 1,30±0,10 * | 0,99±0,07 ° | 1,35±0,09 ** ° | 1,65±0,11 *** ° | |
| Т 4 нмоль/л | ПЛ. | 0,90±0,13 | 1,43±0,29 | 1,25±0,08 | 0,95±0,10 ° | 0,70±0,08 * | 1,49±0,07 °°° | 0,72±0,07 * °°° |
| НЛ. | 1,58±0,15 | 1,72±0,18 | 0,57±0,13 ** | 1,26±0,15 * °° | 0,79±0,14 ** ° | 0,48±0,05 *** | 0,71±0,11 *** | |
| КС нмоль/л | ПЛ. | 22,27±1,49 | 29,84±1,08 •• | 42,01±1,49 *** | 27,0±0,87 °°° | 30,9±0,69 °° | 13,3±0,66 *** °°° | 21,6±0,56 *** °°° |
| НЛ. | 25,98±1,96 | 16,34±1,48 •• | 24,39±1,44 ** | 23,2±0,78 ** | 12,8±0,63 * °°° | 16,0±0,50 °° | 15,7±0,87 |
Условные обозначения (здесь и далее): • - достоверность различий между адаптированным и неадаптированным контролем • - р<0,05; •• - p<0,01; ••• - p<0,001; КНА – контроль неадаптированный, КА – контроль адаптированный; Пол – половозрелые животные, Неп. – неполовозрелые животные.

Рис. 1 Сравнение изменения уровня кортикостерона у адаптированных и неадаптированных животных при холодовом воздействии
Условные обозначения: НА – неадаптированные животные; А – адаптированные животные.
Таким образом, в динамике уровня гормонов были выявлены следующие особенности: 1) у неадаптированных животных холодовое воздействие не изменяло уровень тиреоидных гормонов и приводило к снижению КС на 45-90 мин; уровень всех исследуемых гормонов у неадаптированных животных выше в сравнении с адаптированными; 2) 20-дневное периодическое охлаждение способствовало возрастанию уровня КС у половозрелых животных и снижению у неполовозрелых; 3) у адаптированных животных уровень изучаемых гормонов (Т3, Т4, КС) изменялся на ранних этапах холодового воздействия (на 15 мин); 4) уровень Т4 у адаптированных животных обеих возрастных групп снижался; уровень Т3 у половозрелых животных не изменялся, у неполовозрелых – возрастал; у адаптированных неполовозрелых самцов уровень тиреоидных гормонов изменялся в противоположном направлении: Т3 возрастал, Т4 снижался; 5) у адаптированных животных обеих возрастных групп уровень КС возрастал наиболее интенсивно на 15 мин холодового воздействия, при дальнейшем охлаждении происходило его снижение; 6) у адаптированных и неадаптированных животных наблюдалась сходная динамика изменения уровня гормонов, адаптация приводила к более быстрому проявлению ответной реакции.
Морфометрические показатели щитовидной железы и надпочечников. У неадаптированных половозрелых самцов крыс при холодовом воздействии происходило увеличение относительной массы надпочечников, тогда как относительная масса ЩЖ практически не изменялась. Относительная масса надпочечников и ЩЖ у неадаптированных неполовозрелых самцов крыс имела наибольшие отличия от интактных животных к 45 мин холодового воздействия, но относительная масса ЩЖ возрастала с 0,156±0,012 до 0,242±0,012, тогда как надпочечников – уменьшалась в сравнении с контрольной группой с 0,147±0,002 до 0,220±0,001.
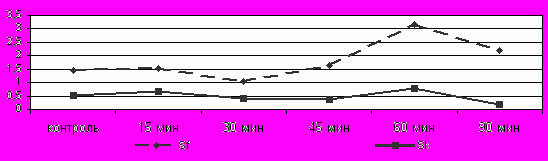
Рис. 2 Изменение площади фолликула и фолликулярной полости щитовидной железы неадаптированных половозрелых самцов крыс
Условные обозначения (здесь и далее): достоверность различий: * - в сравнении с контролем; ˚ - в сравнении с предыдущей группой * (˚) - р<0,05; ** (˚˚) - p<0,01; *** (˚˚˚) - p<0,001.
У неадаптированных половозрелых самцов крыс холодовое воздействие (Рис.2) приводило к уменьшению Sf, достигая наименьшего значения на 45 мин охлаждения. К этому же времени уменьшалась и Sk. Ht возрастала к 30 мин холодового воздействия, снижаясь при более длительном охлаждении. У неадаптированных неполовозрелых животных наиболее значимое снижение морфометрических параметров наблюдалось на 30-й минуте холодовой экспозиции (Sf, Sk, Ht, Sn достоверно снижались, р≤0,001 в сравнении с контролем). Более длительное охлаждение приводило к увеличению Sf и Sk на 60 мин холодового воздействия (р≤0,001), а Ht – на 90 мин (р≤0,001) экспозиции относительно контрольных животных.
У неполовозрелых неадаптированных животных происходило значительное увеличение толщины коры надпочечников и всех ее зон на всем протяжении холодового воздействия. Изменение площади кортикоцитов и площади ядер кортикоцитов зон коры надпочечников зависело от длительности охлаждения и было разнонаправленным.
У
 адаптированных контрольных животных происходило увеличение Sf и Sk. У адаптированных половозрелых животных наибольшие изменения морфометрических показателей ЩЖ при холодовом воздействии происходили на 15 мин экспозиции холодом, когда Sf и Sk возрастали, а Ht и Sn, наоборот, снижались по сравнению с контролем. На 60 мин холодовой экспозиции все морфометрические показатели возрастали в сравнении с контрольной группой. Sn изменялась на всем протяжении холодового воздействия разнонаправлено. У адаптированных неполовозрелых животных холодовое воздействие приводило к возрастанию всех изучаемых морфометрических показателей ЩЖ. При этом наиболее значимые различия происходили на 30 мин охлаждения, когда были максимальны Sf, Sk и Ht, и на 60 мин экспозиции площадь Sn была максимальна, а Sf и Sk минимальны.
адаптированных контрольных животных происходило увеличение Sf и Sk. У адаптированных половозрелых животных наибольшие изменения морфометрических показателей ЩЖ при холодовом воздействии происходили на 15 мин экспозиции холодом, когда Sf и Sk возрастали, а Ht и Sn, наоборот, снижались по сравнению с контролем. На 60 мин холодовой экспозиции все морфометрические показатели возрастали в сравнении с контрольной группой. Sn изменялась на всем протяжении холодового воздействия разнонаправлено. У адаптированных неполовозрелых животных холодовое воздействие приводило к возрастанию всех изучаемых морфометрических показателей ЩЖ. При этом наиболее значимые различия происходили на 30 мин охлаждения, когда были максимальны Sf, Sk и Ht, и на 60 мин экспозиции площадь Sn была максимальна, а Sf и Sk минимальны.
Рис.3 Изменение толщины коры и зон коры надпочечников адаптированных половозрелых самцов крыс
У адаптированных самцов крыс в контроле происходило возрастание коры надпочечников у половозрелых животных за счет увеличения пучковой зоны, а у неполовозрелых – сетчатой и клубочковой зон коры надпочечников в сравнении с интактными самцами. Холодовое воздействие у адаптированных неполовозрелых животных приводило к снижению толщины коры надпочечников к 60 мин экспозиции (Рис.3). Изменение толщины зон коры надпочечников у адаптированных неполовозрелых самцов крыс зависело от длительности холодового воздействия. Толщина клубочковой и пучковой зон возрастала на 30-45 мин охлаждения и снижалась в остальные временные интервалы. Толщина сетчатой зоны снижалась в начальный период холодового воздействия до 30 мин. Холодовое воздействие у адаптированных половозрелых самцов крыс приводило к снижению коры относительно контрольной группы с 15 по 45 мин. У адаптированных половозрелых животных толщина клубочковой зоны снижалась на всем протяжении холодового воздействия, толщина пучковой зоны снижалась до 45 мин воздействия, тогда как толщина сетчатой зоны возрастала на 15 и 45 мин охлаждения. Относительная масса ЩЖ и надпочечников у адаптированных животных обеих возрастных групп при охлаждении практически не изменялись.
Таким образом, были выявлены следующие особенности морфометрических показателей: 1) холодовое воздействие вызывало ответную реакцию со стороны эндокринных органов: происходило реагирование со стороны ЩЖ и надпочечников, начиная с 15 минуты холодовой экспозиции; 2) в период 20-дневной адаптации происходила гистоструктурная перестройка ЩЖ и надпочечников к последующему воздействию: в ЩЖ – накопление коллоида, в надпочечниках – увеличение толщины коры; 3) адаптация приводила к снижению реакции коры надпочечников на холодовое воздействие: у неадаптированных животных происходило увеличение толщины коры надпочечников, тогда как у адаптированных самцов крыс толщина коры при холодовом воздействии снижалась; 4) у адаптированных половозрелых животных активации коры надпочечников на острое холодовое воздействие отсутствовала, у адаптированных неполовозрелых животных - снижалась; 5) в ЩЖ адаптация способствовала более раннему проявлению ответной реакции (у неадаптированных животных рост показателей происходил лишь к 60 мин, у адаптированных на 15-30 мин); 6) у адаптированных животных наибольшие морфометрические изменения в ЩЖ наблюдались на 60 мин холодового воздействия: у половозрелых животных происходило максимальное возрастание всех изучаемых параметров, у неполовозрелых – Sk и Sf возрастали, а Sn и Ht – снижались.
Динамика ПОЛ. Содержание МДА в ЩЖ у неадаптированных неполовозрелых самцов крыс снижалось на 45 мин холодового воздействия; скорость спПОЛ уменьшалась на всем протяжении охлаждения за исключением 30 мин, скорость аскПОЛ снижалась к 15 мин холодового воздействия, тогда как дальнейшее охлаждение способствовало возрастанию показателя. В надпочечниках при охлаждении изменялись лишь кинетические показатели ПОЛ - происходило снижение спПОЛ и аскПОЛ с 45 по 90 мин холодового воздействия.
У контрольных адаптированных животных происходило снижение уровня ПОЛ и в ЩЖ и в надпочечниках в сравнении с интактными самцами. У адаптированных неполовозрелых самцов крыс скорость аскПОЛ в ЩЖ (Табл.2) снижалась на всем протяжении холодового воздействия, за исключением 45 мин; скорость спПОЛ в ЩЖ снижалась к 15 - 30 мин холодового воздействия.
Таблица 2
Динамика МДА и кинетических показателей ПОЛ в щитовидной железе адаптированных самцов крыс
| Воздействие | Скорость аскПОЛ, нмоль МДА/ч | Скорость спПОЛ, нмоль МДА/ч | ||
| Пол. | Неп. | Пол. | Неп. | |
| КНА | 101,00±8,13 ΔΔΔ | 41,60±3,21 | 97,02±5,75 ΔΔΔ | 20,53±1,54 |
| КА | 104,81±12,92 ΔΔΔ | 34,22±3,28 •• | 94,75±11,59 ΔΔ | 28,71±4,47 •• |
| 15 мин | 76,56±4,77 ΔΔΔ *** | 36,73±3,88 * | 84,89±7,53 ΔΔΔ ** | 40,32±5,21 ** |
| 30 мин | 83,23±7,4 ΔΔΔ *** | 22,43±4,02 *** °°° | 90,01±6,29 ΔΔΔ * | 24,28±2,76 °° |
| 45 мин | 100,90±10,06 ΔΔΔ | 28,65±6,21 *** °°° | 97,43±8,83 ΔΔΔ °°° | 21,03±1,88 |
| 60 мин | 92,66±8,38 ΔΔΔ ** | 26,45±6,01 *** | 96,53±9,34 ΔΔΔ | 33,22±4,58 °° |
| 90 мин | 95,58±10,28 ΔΔΔ ** | 24,12±4,74 *** | 90,74±9,22 ΔΔΔ ° | 20,04±2,88 ** °°° |
Условные обозначения (здесь и далее): Δ - достоверность различий между половозрелыми и неполовозрелыми животными ΔΔ - p<0,01; ΔΔΔ - p<0,001.
В надпочечниках адаптированных неполовозрелых животных к 30 мин холодового воздействия происходило повышение скорости аскПОЛ (Табл.3). После 45 мин воздействия холодом скорость аскПОЛ возвращалась к контрольному уровню, вновь повышаясь к 60 - 90 мин воздействия по сравнению с контрольной группой.
Влияние холодового воздействия на скорость спПОЛ в ЩЖ (Табл.2) у адаптированных половозрелых самцов крыс зависело от времени действия данного фактора. Происходило увеличение скорости спПОЛ на 15 мин холодового воздействия, тогда как после 90 мин охлаждения скорость спПОЛ снижалась. Холодовое воздействие приводило к снижению скорости аскПОЛ в течение всего периода, начиная с 30 мин.
Таблица 3
Динамика МДА и кинетических показателей ПОЛ в надпочечниках адаптированных самцов крыс
| воздействие | Скорость аскПОЛ, нмоль МДА/ч | Скорость спПОЛ, нмоль МДА/ч | ||
| Половозрелые | Неполовозрелые | Половозрелые | Неполовозрелые | |
| КНА | 73,44±7,88 ΔΔ • | 38,9±5,38 ••• | 68,23±4,77 ΔΔΔ | 23,5±2,34 |
| КА | 65,74±4,64 ΔΔΔ | 23,33±2,71 | 67,45±4,38 ΔΔΔ | 24,56±1,30 |
| 15 мин | 69,91±3,97 ΔΔΔ | 23,55±5,06 | 68,07±4,60 ΔΔΔ | 21,09±1,89 |
| 30 мин | 70,77±4,05 ΔΔΔ ** | 27,62±4,12 ° | 72,52±0,72 ΔΔΔ | 21,52±2,67 |
| 45 мин | 66,72±4,32 ΔΔ | 35,58±7,67 ** °°° | 71,25±4,10 ΔΔΔ * | 21,04±1,88 |
| 60 мин | 72,95±4,74 ΔΔΔ **° | 31,99±5,19 * | 68,18±4,71 ΔΔΔ | 19,93±3,02 |
| 90 мин | 73,36±5,51 ΔΔΔ *** | 41,69±2,03 *** °° | 69,39±5,37 ΔΔΔ | 20,04±2,88 |
В надпочечниках адаптированных половозрелых животных скорость аскПОЛ (Табл.3) возрастала с 45 мин холодового воздействия, оставаясь высокой до конца охлаждения. Холодовое воздействие способствовало увеличению исходного содержания МДА к 15 и 90 мин. ПОЛ в ЩЖ имело отрицательную корреляцию с уровнем Т4, а в надпочечниках положительную корреляцию с уровнем КС.
Динамика каталазной ативности. Каталазная активность в ЩЖ у неадаптированных неполовозрелых самцов повышалась на всем протяжении охлаждения, за исключением 45 мин экспозиции (Рис.4), тогда как в надпочечниках - снижалась.
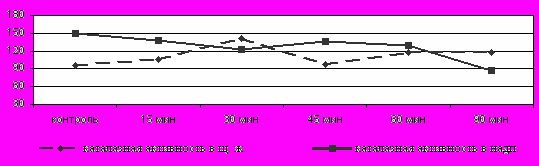
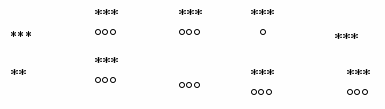
Рис. 4 Изменение каталазной активности в щитовидной железе и надпочечниках у неадаптированных неполовозрелых самцов крыс
Была выявлена положительная коррелляционная зависимость (р≤0,001) между уровнем тиреоидных гормонов и каталазной активностью на 30 минуте охлаждения.
У контрольных адаптированных самцов крыс происходило возрастание каталазной активности в ЩЖ и снижение в надпочечниках в сравнении с интаткным контролем. Холодовое воздействие приводило к значительному снижению каталазной активности у адаптированных половозрелых животных в ЩЖ и надпочечниках (Табл.4) на всем протяжении холодовой экспозиции.
Таблица 4
Изменение каталазной активности в щитовидной железе и надпочечниках адаптированных половозрелых самцов крыс
| | КНА | КА | 15 мин | 30 мин | 45 мин | 60 мин | 90 мин |
| ЩЖ | 156,9±1,40 | 285,7±2,40 ••• | 192,4±2,91 *** | 168,9±1,85 *** °°° | 160,6±2,28 *** ° | 269,7±1,29 *** °°° | 276,5±1,18 ** °° |
| Надпоч. | 659,3±1,20 | 334,5±0,11 ••• | 70,0±1,10 *** | 185,2±2,23 *** °°° | 113,4±1,24 *** °°° | 276,7±2,10 *** °°° | 322,2±0,69 *** °°° |
У
 адаптированных неполовозрелых животных холодовое воздействие в ЩЖ (Рис.5) приводило к значительному снижению каталазной активности на всем протяжении, а в надпочечниках к возрастанию показателя на 15 мин экспозиции.
адаптированных неполовозрелых животных холодовое воздействие в ЩЖ (Рис.5) приводило к значительному снижению каталазной активности на всем протяжении, а в надпочечниках к возрастанию показателя на 15 мин экспозиции. 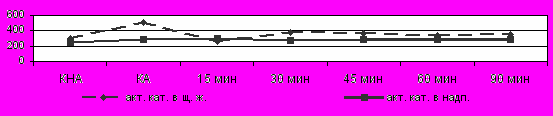
Рис. 5 Изменение каталазной активности в щитовидной железе и надпочечниках у адаптированных неполовозрелых самцов крыс
Таким образом, были выявлены следующие особенности свободнорадикального гомеостаза: 1) уровень ПОЛ у половозрелых животных выше, в сравнении с неполовозрелыми; 2) 20-дневное периодическое охлаждение приводило к снижению ПОЛ в ЩЖ и надпочечниках, возрастанию каталазной активности в ЩЖ и снижению в надпочечниках; 3) холодовое воздействие приводило к разнонаправленным изменениям ПОЛ: у адаптированных животных происходило возрастание, у неадаптированных снижение интенсивности ПОЛ в ЩЖ и надпочечниках; 4) динамика каталазной активности различалась в зависимости от изучаемой ткани и возраста животных: у половозрелых животных каталазная активность снижалась, у неполовозрелых – снижалась в ЩЖ и возрастала в надпочечниках; у неадаптированных животных изменения были противоположны: возрастание каталазной активности в ЩЖ и снижение в надпочечниках.
Полученные результаты показали, что холодовое воздействие вызывало ответную реакцию со стороны эндокринных органов, при этом происходило изменение уровня тиреоидных гормонов и КС, гистоструктуры ЩЖ и надпочечников, свободнорадикального окисления и каталазной активности уже на 15 мин экспозиции. Поступление гормонов щитовидной железы и надпочечников в ткани и органы-мишени происходило в разные временные периоды холодового воздействия. Так наблюдалось снижение уровня тироксина в крови адаптированных животных на 15 минуте холодового воздействия, а уровня кортикостерона на 45-60 мин охлаждения. Изменение уровня гормонов в крови согласуется с динамикой кинетических показателей ПОЛ. Происходило изменение скорости спПОЛ и аскПОЛ в щитовидной железе с 15 по 90 мин охлаждения, а в надпочечниках – с 45 по 90 мин холодовой экспозиции.
20-дневное периодическое охлаждение приводило к формированию компенсаторно-приспособительных механизмов функционирования щитовидной железы и надпочечников, которые выражались в гистоструктурной перестройке органов (адаптационной гипертрофии коры надпочечников и накоплении коллоида в фолликулах щитовидной железы), изменении уровня кортикостерона в крови и процессов свободнорадикального окисления. Это приводило к ограничению развития реакции коры надпочечников на острое холодовое воздействие у адаптированных животных в сравнении с неадаптированными самцами крыс, у которых происходило значительное возрастание коры надпочечников. При периодических воздействиях происходит адаптивная перестройка, повышается метаболизм макромолекул, при повторном воздействии механизм ответной реакции запускается сразу, т.к. протекает на фоне измененного адаптированного к стрессорному воздействию метаболизма клеточных структур.
Как известно, существуют несколько критериев, отражающих развитие стресс-реакции. К ним относятся уровень кортикостероидов в крови, изменение относительной массы надпочечников, изменение толщины коры надпочечников и другие. Как показали наши исследования наиболее информативным показателем оказались морфометрические: толщина коры и толщина пучковой зоны коры надпочечников, которые проявили наиболее четкие различия в ответной реакции на острое охлаждение у адаптированных и неадаптированных животных. Уровень кортикостероидов в плазме крови не всегда напрямую отражает изменение функциональной активности коры надпочечников в ответ на стрессовое воздействие, т.к. параллельно с выбросом в кровь гормонов происходит и их утилизации тканями и органами-мишенями.
Формирование компенсаторно-приспособительных механизмов функционирования щитовидной железы и надпочечников при периодическом охлаждении зависело от возраста. У адаптированных неполовозрелых животных проявлялись неспецифические компоненты ответной реакции, которые выражались в реакции на острое холодовое воздействие коры надпочечников. У адаптированных половозрелых животных начинают преобладать специфические (тиреоидные, свободнорадикальные, антиоксидантные) механизмы. Это согласуется с литературными данными, (Пронина Т.С., Шитов Л.А. 2004) о более высоких потенциальных возможностях адаптации у взрослых животных. В процессе онтогенеза происходит формирование и становление механизмов адаптации.
ВЫВОДЫ
- Динамика уровня кортикостерона при холодовом воздействии у адаптированных и неадаптированных животных сходна: возрастает в ранний период охлаждения (15-30 мин) и снижается в период с 45 по 90 мин. независимо от возраста. Уровень тиреоидных гормонов при периодическом холодовом воздействии изменяется в зависимости от возраста: у неполовозрелых самцов уровень трийодтиронина возрастал, а уровень тироксина снижался; у половозрелых крыс снижался только уровень тироксина;
- В период 20-дневного периодического охлаждения происходит формирование компенсаторно-приспособительной реакции щитовидной железы; у адаптированных животных обеих возрастных групп реакция щитовидной железы на холодовое воздействие проявляется раньше на 15-30 минутах; наиболее интенсивное возрастание морфометрических показателей щитовидной железы происходит на 60 минуте холодового воздействия;
- Периодическое 20-дневное охлаждение в надпочечниках приводит к увеличению толщины коры, способствуя формированию компенсаторно-приспособительных механизмов; у адаптированных животных происходит снижение реакции коры надпочечников на холодовое воздействие: у неадаптированных животных толщина коры надпочечников увеличивается, у адаптированных - уменьшается; толщина коры надпочечников у адаптированных животных снижается на всем протяжении холодового воздействия;
- Направление изменения интенсивности перекисного окисления липидов при однократном холодовом воздействии отличается у адаптированных и неадаптированных самцов крыс: у адаптированных животных происходит активация свободнорадикальных процессов, а у неадаптированных -снижение интенсивности ПОЛ в тканях щитовидной железы и надпочечников.
- Интенсивность перекисного окисления липидов в ткани надпочечников положительно коррелирует с уровнем кортикостерона; установлена отрицательная корреляция установлена между интенсивностью процессов ПОЛ в ткани щитовидной железы и уровнем тироксина в крови.
- Динамика каталазной активности при холодовом воздействии имеет возрастные, тканеспецифические особенности и различается у адаптированных и неадаптированных животных: у неадаптированных животных каталазная активность при охлаждении возрастает в щитовидной железе и снижается в надпочечниках; у адаптированных самцов крыс изменения противоположны; у половозрелых животных каталазная активность в обоих органах снижается;
Список научных работ, опубликованных по теме диссертации
1. Тимонина, Л.А. (Яковенкова Л.А.) Изменение показателей перекисного окисления липидов в тканях мозга и печени под действием стресс-индуцирующих агентов [Текст] / М.В. Мажитова, А.Г. Глинина, Д.Л.Теплый, Т.А. Кузьмина, Н.Е. Береговая, Л.А Тимонина // Тезисы докладов итоговой научной конференции АГПУ им. С.М. Кирова. Астрахань: Издательство АГПУ. - 1999. - С. 64. ISBN 5-88200-420-9-(общ), - ISBN 5-88200-426-8. – 0,063 п.л, личный вклад 16,7 %.
2. Тимонина, Л.А. (Яковенкова Л.А.) Влияние доксорубицина на перекисное окисление липидов печени и мозга белых крыс [Текст] / М.В. Мажитова, Т.А. Кузьмина, Л.А. Тимонина // Материалы VII итоговой научной конференции молодых ученых и студентов СГМА. Ставрополь: Издательство СГМА. - 1999. - С. 231-233. - 0,188 п.л, личный вклад 33,3 %.
3. Просина, Л.А. (Яковенкова Л.А.) Морфофункциональное состояние щитовидной железы самцов крыс на ранней стадии холодового воздействия. [Текст] / Е.И. Кондратенко, Л.А. Просина // Материалы VIII Международной научной конференции "Эколого-биологические проблемы бассейна Каспийского моря». Астрахань: Изд. дом «Астраханский университет». - 2005. - С. 206-208. - ISBN 5-88200-851-4. - 0,188 п.л, личный вклад 50 %.
4. Просина, Л.А. (Яковенкова Л.А.) Ранняя динамика уровня МДА и активности каталазы в гомогенатах тканей щитовидной железы и надпочечников у крыс, адаптированных к периодическому охлаждению [Текст] / Л.А. Просина // Материалы Международной научной конференции «Свободные радикалы, антиоксиданты и старение». Астрахань: Изд. дом «Астраханский университет». - 2006. - С. 163-165. ISBN 5-88200-909-х. - 0,188 п.л, личный вклад 100 %.
5. Просина, Л.А. (Яковенкова Л.А.) Перекисное окисление липидов и активность каталазы в крови крыс, получавших витамин Е, в условиях острого холодового воздействия [Текст] / К.А. Джумалиев, О.Н. Позднякова, Л.А. Просина // Современные наукоемкие технологии. - 2006. - № 2. - С. 67. - 0,063 п.л, личный вклад 33,3 %.
6. Просина, Л.А. (Яковенкова Л.А.) Ранние изменения уровня продуктов перекисного окисления липидов в крови и тканях у крыс, адаптированных к холодовому воздействию [Текст] / О.Н. Позднякова, Л.А. Просина, Е.И. Кондратенко, Е.В. Курьянова, О.Г. Поздняков // Вестник поморского университета. Серия: «Естественные и точные науки». - Выпуск 3. 2006. - С.42-44. – ISBN 1728-7340. (Издание рекомендованное ВАК). - 0,188 п.л, личный вклад 20 %.
7. Просина, Л.А. (Яковенкова Л.А.) Особенности ориентировочно-исследовательской реакции у крыс в процессе выработки адаптации к холоду [Текст] / О.Н. Позднякова, Л.А. Просина // Материалы IX Международной научной конференции "Эколого-биологические проблемы бассейна Каспийского моря». Астрахань: Изд-во АГУ. - 2006. - С.212-213. - ISBN 5-88200-907-3. - 0,063 п.л, личный вклад 50 %.
8. Просина, Л.А. (Яковенкова Л.А.) Ранние изменения уровня продуктов перекисного окисления липидов в крови у крыс, адаптированных и неадаптированных к холоду [Текст] / О.Н. Позднякова, Л.А. Просина // Вестник Астраханского государственного технического университета. Астрахань: Изд-во АГТУ. 2006. - №3 (32). - С. 207-211. –ISSN 1812-9498. (Издание рекомендованное ВАК). - 0,25 п.л, личный вклад 50 %.
9. Просина, Л.А. (Яковенкова Л.А.) Ранняя динамика уровня малонового диальдегида в гомогенатах тканей щитовидной железы и надпочечников у неполовозрелых самцов крыс, адаптированных к периодическому охлаждению [Текст] / Л.А. Просина // Сборник научных трудов «Фундаментальные исследования в биологии и медицине». - Вып.3. – Ставрополь: Изд-во СевКавГТУ. - 2007. - С.268. - 0,063 п.л, личный вклад 100 %.
10. Просина, Л.А. (Яковенкова Л.А.) Свободнорадикальные процессы в ткани щитовидной железы и надпочечников неполовозрелых самцов крыс в ранний период холодового воздействия и их зависимость от адаптации к периодическому охлаждению [Текст] / Л.А. Просина, Н.А. Ломтева // Естественные науки. - 2007. - №4 (21). С. 73-77. - ISBN -1818-5-507-Х. (Издание рекомендованное ВАК). - 0,25 п.л, личный вклад 50 %.
11. Просина, Л.А. (Яковенкова Л.А.) Ранние изменения морфометрических показателей щитовидной железы половозрелых и неполовозрелых самцов крыс, адаптированных к периодическому холодовому воздействию [Текст] / Л.А. Просина // Сборник научных статей, посвященный 35-летию кафедры анатомии, физиологии и гигиены человека и 85-летию со дня рождения основателя кафедры, заслуженного деятеля науки РФ, доктора медицинских наук, профессора Држевецкой И.А. «Физиологические проблемы адаптации». - Ставрополь: Изд-во СГУ. - 2008. - С.219-222. - 0,25 п.л, личный вклад 100 %.
12. Просина, Л.А. (Яковенкова Л.А.) Изменение уровня малонового диальдегида в гомогенатах тканей щитовидной железы у неполовозрелых крыс, адаптированных к периодическому охлаждению [Текст] / Л.А. Просина // Тезисы докладов VI Сибирского физиологического съезда.– Барнаул: Принтэкспресс - 2008. - в 2-х томах. Т.II. - С.65. - 0,063 п.л, личный вклад 100 %.
13. Яковенкова, Л.А. Ранняя динамика тиреоидных гормонов и кортикостерона в крови неполовозрелых самцов крыс, адаптированных к периодическому холодовому воздействию [Текст] / Л.А. Яковенкова // Материалы докладов XVI Международной конференции студентов, аспирантов и молодых ученых «Ломоносов» / Отв. ред. И.А. Алешковский, П.Н. Костылев, А.И. Андреев. [Электронный ресурс] – М.: Издательство МГУ, 2009. – 1 электрон. опт. – С. 54. ISBN 978-5-317-02774-2. - 0,063 п.л, личный вклад 100 %.
14. Яковенкова, Л.А. Морфометрические изменения щитовидной железы неполовозрелых самцов крыс в ранний период холодового воздействия [Текст] / Л.А. Яковенкова // Сборник трудов конференции молодых ученых. Выпуск 2. Биомедицинские технологии, мехатроника и робототехника. – СПб: СПбГУ ИТМО, 2009. – С 48. ISSN 978-5-7577-0335-0. - 0,063 п.л, личный вклад 100 %.
Принятые сокращения и условные обозначения
ПОЛ – перекисное окисление липидов,
МДА – малоновый диальдегид,
Т3 – трийодтиронин,
Т4 – тироксин
КС – кортикостерон
ПГЭ – перекисный гемолиз эритроцитов
ЩЖ – щитовидная железа
аскПОЛ – аскорбатзависимое ПОЛ
спПОЛ – спонтанное ПОЛ
Sf – площадь фолликула
Sk – площадь фолликулярной полости
Ht – высота тироцитов
Sn – площадь ядра тироцитов.