
И. И. Ш м альгаузен избранные труды организм как целое в и ндивидуальном и и сторическом издательство «наука» москва 1982 развитии
| Вид материала | Книга |
- Сергей Лычагин «Макиавелли Н. Избранные произведения.», 922.62kb.
- И. И. Веселовског о издательство "наука" Москва 1967 Эта книга, 1700kb.
- Избранные научные труды. М.: Наука, 1988, с. 722-734, 363.13kb.
- А. Н. Леонтьев Избранные психологические произведения, 6448.08kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 206.83kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 238.78kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 208.81kb.
- Д. Н. Мамине-Сибиряке Книга, 262.07kb.
- Литература Особенности типологического подхода и метода исследования личности, 2747.84kb.
- А. Н. Леонтьев Избранные психологические произведения, 6931kb.
4. Адаптивные модификации как изменения,
обусловленные существованием сложной
исторически сложившейся реакционной системы
Мы много говорили о том, что мутация может иметь разное выражение в зависимости, с одной стороны, от генетических различий и, с другой стороны, от различий во внешней среде. Новые мутации оказываются часто весьма мало устойчивыми, т. е. в своем проявлении и выражении оказываются сильно зависимыми от изменений как самого генотипа, так и от изменений в факторах внешней среды. Новые мутации, как правило, не адаптивны в своих выражениях, и соответственно их модификации также не представляют собой адаптации. У дрозофилы мутация «pennant», по мере повышения температуры воспитания личинок, получает все более укороченные крылья; мутация «short wing» получает также при повышении температуры все более резкое выражение; то же самое наблюдается у мутации «Ваг», у которой уменьшается число фасеток глаз. У мутации «Abnormal abdomen» выражение усиливается при воспитании на влажном корме. Мутация «bent» получает при низкой температуре целый Ряд добавочных выражений — кроме изогнутых крыльев, еще погрубение глаз, нарушение расположения щетинок на груди и изменение в одной из жилок крыла.
Ничего приспособительного в этих реакциях нет. Если, однако, какая-либо мутация, в каком-либо из своих выражений окажется в известных условиях благоприятной, то это быстро приведет к отбору этих благоприятных форм реагирования при одновременном подавлении других выражений этой мутации
138 Организм как целое в индивидуальном и историческом развитии
(через подбор модификаторов). Благоприятное выражение мутации, или адаптивная форма реагирования, приобретет тогда устойчивый характер и войдет в состав системы нормальных реакций данной мутации.
Устойчивость выражения, однако, вовсе не означает независимости от внешней среды, она указывает лишь на строгую определенность реакции. Реакция же может быть различной в разных условиях. Классическим примером двух различных, но совершенно определенных реакций является китайский первоцвет (Primula chinensis), известный в нескольких наследственно стойких расах. Известны расы, дающие при разных температурах белые цветы, и расы, обладающие при тех же условиях красными цветами. Здесь эти различия обусловлены различиями в генотипе. Однако известна раса того же первоцвета, дающая при умеренной температуре белые цветы, а в оранжерее (при температуре около 30°) — красные. Здесь имеются, следовательно, у одного генотипа (даже одной особи) две определенные нормы реакций на различные температуры. Обе реакции обусловлены наследственными свойствами данной расы, и, конечно, нельзя ту или иную окраску назвать ненаследственной. Однако изменение красной окраски цветов в белую при переносе растения из оранжереи в комнату зависит от изменения температуры. Такое изменение фенотипа мы называем модификацией. Мы можем говорить о модификации лишь при сравнении одного фенотипа с другим, при условии идентичности генотипа.
Раньше мы говорили о «выражении» известной мутации при определенных условиях. Однако мы видели, что это выражение зависит не от одного гена, а от всего генотипа, и, следовательно, понятия «выражение» мутации и «выражение» генотипа, т. е. фенотип, совпадают. Изменение выражения мутации, или изменение фенотипа, при изменении внешних условий мы должны назвать модификацией. Если бы вся естественная популяция дрозофилы состояла из формы «Abnormal adbomen», которая при разных условиях развития (на свежем или на подсохшем корме) дает либо уродливое брюшко, либо «нормальное», мы бы при сравнении этих фенотипов друг с другом говорили о модификациях, вызываемых изменением характера пищи. Теперь же мы говорим обычно лишь о «проявлении» признака уродливого брюшка при определенных условиях культуры. «Уродливое брюшко» является модификацией у мутанта, но, конечно, модификацией не адаптивной, т. е. морфозом. Точно так же проявление и выражение удвоения конечностей у мутации «reduplicated» находится в такой сильной зависимости от температуры среды, в которой развиваются личинки, что мы также можем сказать, что эта мутация характеризуется именно существованием специального температурного морфоза (удвоение ножек как неадая-тивная реакция на низкую температуру).
Глава IV. Прогрессивная эволюция. Адаптациогенез 139
Генотипически определенная форма может, однако, давать не две определенные, ярко выраженные и качественно друг от друга отличные реакции, но много таких реакций. В очень многих случаях такие реакции отличаются количественно и связаны непрерывными переходами. В работах В. Шульца [W. Schultz, 1920, 1922] и Н. А. Ильина [1926, 1927] исследованы изменения пигментации горностаевого кролика при разных температурах. При «нормальной» температуре эти кролики имеют белую шерсть, и только нос, уши и хвост.— черные. При понижении температуры ниже 16° черную окраску получают задние лапы, а при температуре ниже 14° — и передние. При температурах ниже 2—3° черная окраска шерсти начинает распространяться на все тело. С другой стороны, при повышении температуры выше 25° появляются белые волосы на ушах, а затем, при температуре выше 29°, и на хвосте и на носу, так что весь кролик становится белым. Каждая часть тела имеет свой порог раздражения, т. е. свою максимальную температуру, при которой еще возможно образование черного пигмента. Разная высота этого порога зависит от различного кровоснабжения кожи в разных частях тела, с чем связано и различное охлаждение кожи при понижении температуры в окружающей среде. Вместе с тем в разных генетических линиях этих кроликов порог раздражения может сильно различаться. У саксонской расы порог раздражения кожи бока равен + 21°, у польской расы +11°, а у московской, как сказано, около + 2°. Имеются и наследственные индивидуальные различия в высоте порога раздражения.
Здесь мы имеем, следовательно, дело с одной реакцией, обладающей различными «нормами» для разных частей тела и, следовательно, распространяющейся на различные его участки при разных температурах среды.
Переходы здесь скачкообразные — при постепенном охлаждении достижение известного порога сказывается черной пигментацией, сразу захватывающей целый участок покровов (например, всю лапу). В других случаях признак может иметь количественное выражение, и тогда постепенное изменение, скажем, температуры, проявится в таком же постепенном изменении известных признаков. Так, например, приведенный уже пример мутации «pennant» у дрозофилы отличается крыльями, похожими по виду на нормальные, однако их величина сильно зависима от температуры. При повышении температуры от 16 до 30° величина крыльев постепенно уменьшается [Harnly M. H., HarnlyM. L., 1936]. Или, например, по опытам Семнера [Sumner, 1909] повышение температуры приводит к развитию более длинных хвостов 11 ушей у мышей и крыс, а понижение температуры связано с их Укорочением. У дафний, по Вольтереку [Woltereck, 1909], усиленное питание приводит к возрастанию высоты их шлема. Вообще, Условия питания, конечно, имеют большое значение для разви-
140 Организм как целое в индивидуальном и историческом развитии
тия фенотипа, и их изменения отражаются главным образом на изменении величины всего организма или его частей. И все же даже такой признак, как величина тела, определяется генотипом и характеризует его не в меньшей мере, чем другие фенотипи-ческие признаки, по которым мы только и можем иметь суждение о генотипе.
Как же мы должны рассматривать эти фенотипические изменения, зависящие от факторов внешней среды? Прежде всего,
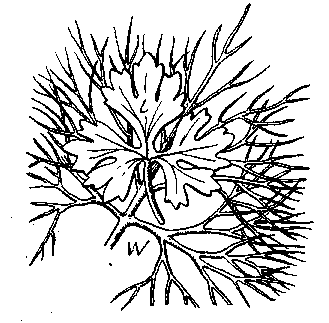
Рис. 16. Водный (W) и воздушный (L) лист водяного лютика (Ranunculus delphinifolius) [Гёбелъ, 1928]
отметим, что организм в своем развитии вообще полностью зависит от внешней среды, без которой невозможны ни жизнь, ни развитие. Организм, конечно, настолько тесно связан со средой, что ее изменения не могут не влечь за собой изменений развивающегося организма. Однако по отношению к изменениям отдельных факторов среды организм может оказаться весьма устойчивым в том смысле, что в известных пределах изменений он заметно не меняется. Так, при изменении температуры внешней среды от 3 до 14° и точно так же в пределах от 16 до 25° горностаевый кролик не изменяет своей окраски.
Для очень многих признаков можно указать пределы колебаний факторов внешней среды, внутри которых данные признаки заметно не меняются. Это и есть «нормальное» выражение признака, и возможность его развития ограничивается известными пределами «нормальной» среды. Нормальной же средой является та обычная среда, в которой исторически развивался данный организм. Такая выработка известных весьма устойчивых форм есть всегда результат долгого исторического развития. Если организм обладает не одной, а двумя или более резко выраженными «нормами», то он и живет нормально не в одной среде и исторически развивается как вид сразу в нескольких средах. Так, например, водяной лютик имеет два определенно и резко выраженных фенотипа — один из них характерен для жизни в воде, другой —для суши (рис. 16). Для стрелолиста характерны даже три нормы — одна развивается на суше, другая в мелкой воДе (плавающие листья) и третья — в глубокой воде (подводные лей' товидные листья). Различные особи этого вида жили и раньше в различных средах, и все эти нормы реакций представляют ре' зультат исторического развития вида в этих разнообразных усло-
Глава IV. Прогрессивная эволюция. Адаптациогенез 141
виях. Чем разнообразнее условия, в которых развивается организм, тем более разнообразны и его нормы реакций, тем более резко выражается его модификационная изменчивость.
Модификационная изменчивость имеет, следовательно, также свою историю. Однако эта история идентична с историей генотипа и обусловлена она теми же факторами (естественный отбор в тех средах, в которых обитает организм). Исторически развиваются только организмы с их наследственной базой (генотипом), выражение которой (фенотип) зависит, конечно, также и от условий развития данной особи.
Характер модификационной изменчивости может быть различным, и это определяется, как сказано, историческим развитием и осуществляется через механизм индивидуального развития организма, который может показывать большую или меньшую степень автономности, т. е. большую или меньшую независимость от колебаний факторов среды в известных пределах. Нелегко решить вопрос о том, в каком направлении изменяется модификационная изменчивость в течение эволюции. Во всяком случае, неправильно безоговорочно принимать сокращение модификационной изменчивости, а тем более говорить об историческом «фиксировании» модификаций, как это делают Вайденрайх и другие неоламаркисты. Нельзя также говорить о прогрессивном «застывании форм» или об освобождении организма из-под власти среды и т. п., как это мы видели у Бойрлена и других идеалистов. Неверно также, если говорят о первичном характере модификационной изменчивости и противопоставляют ее мутационной как вторичной. Оба рода изменчивости исторически развивались одновременно и выражают различную степень зависимости вида (мутации) или особи (модификации) от изменений внешней среды. Правильнее было бы поставить совершенно обратный вопрос о прогрессивном развитии видовой устойчивости организма, т. е. прочного наследования (мутации являются нарушениями этой прочности наследования), и о развитии индивидуальной устойчивости организма, т. е. о возникновении «автономного» механизма развития, ведущего к определенной типичной структуре независимо от колебаний факторов среды (модификации можно бы рассматривать как нарушения «типичной» структуры). Видовая устойчивость развивалась путем выдифференцировки устойчивого наследственного механизма, связанного со сложной системой корреляций как аппарата его реализации (т. е. онтогенеза), а индивидуальная устойчивость развивалась путем установления регуляторного характера и прогрессивной автономизации этого аппарата. В результате этого достигается максимальная защита Формообразовательных процессов от влияния случайных уклонений от нормальных условий развития. Во всяком случае, мы не можем отделить модификационной изменчивости от мутацион-
142 Организм как целое в индивидуальном и историческом развитии
ной и должны их историю рассматривать совместно, так как в эволюции они взаимозависимы.
Мы уже связали раньше модификации (т. е. изменения фенотипа) с мутациями (т. е. изменением генотипа), рассматривая первые как видоизменение конкретных форм выражения вторых (мутаций) при различных условиях внешней среды. Новые мутации имеют часто не совсем постоянное выражение, т. е. оказываются весьма зависимыми от изменений в факторах внешней среды. Конечно, всякий организм, а тем более мутантный, в своем развитии должен в той или иной мере зависеть от факторов внешней среды.
Однако совершенно невероятно, чтобы новый признак, развившийся при известных нормальных условиях среды и дающий организму хотя бы небольшие преимущества, т. е. полезный, развился при иных условиях, хотя и иначе, но все-таки как полезный в этих условиях признак. Признание этого было бы равносильно признанию чуда. Мы уже говорили, что и сама полезность признака относительна и есть результат исторического развития организма (подбор комбинированных мутаций и их модификаторов) в определенной среде. Тем более мы должны сказать, что если модификационное изменение оказывается для организма выгодным, то эта целесообразность его реагирования на изменение факторов внешней среды есть, во всяком случае, результат исторического развития формы реагирования. Это может быть, однако, достигнуто исключительно через естественный отбор генотипов при встречающихся в природе уклонениях факторов внешней среды от обычной нормы. Целесообразные реакции и наблюдаются только в пределах таких изменений факторов среды, которые в природе действительно встречаются.
В прогрессивной эволюции имеют значение только такие мутации, которые дают организму известные преимущества, и то же самое надо сказать и о модификациях. Целесообразность последних не может быть положена в основу каких-либо попыток объяснения эволюции (ламаркизм), а должна быть рассматриваема как результат очень сложного исторического процесса развития организма в непостоянной внешней среде. Это процесс не менее, а более сложный, чем история развития признаков, однозначно определяемых известной комбинацией мутаций.
Нужно думать, что в процессе эволюции образование новых признаков путем подбора мутаций идет одновременно с установлением определенных, также целесообразных норм реакций, определяющих полезное выражение данного признака при различных конкретно встречающихся в данной среде условиях развития. Таким образом, одновременно с новым признаком развивается и механизм, определяющий его развитие в разных условиях, т. е, устанавливаются различные его модификации. Если организм развивается в относительно постоянной среде, то его
Глава IV. Прогрессивная эволюция. Лдаптацидгёнёз 143
модификабельность будет небольшой; если эта среда сильно изменчива, то модификабельность может быть выше. Если организм живет одновременно в двух разных средах (одни особи — в одной, другие — в другой), то он может принять две различные формы (водяной лютик). Если организм по своей организации сильно зависим от меняющихся факторов внешней среды и xie может активно менять свое отношение к этим факторам (растения) , то модификационная изменчивость должна быть выше. Если же организм в значительной мере сам выбирает себе условия среды (подвижность животных) и среда, следовательно, оказывается для него более постоянной, то модификационная изменчивость будет ниже. Еще ниже становится модификационная изменчивость, если в самой организации развиваются механизмы, изолирующие организм от изменений факторов внешней среды. Так, например, теплокровность высших позвоночных освобождает их в значительной мере от температурных колебаний среды.
Во всех этих случаях история организма наложила яркий отпечаток на особенности его модификационной изменчивости, и это лучше всего доказывает историческую обоснованность последней. Никакой иной механизм, как только естественный отбор организмов, обладающих наиболее благоприятной системой реакций, обеспечивающей их жизнь в данных меняющихся условиях, не может быть ответственным за развитие целесообразной модификационной изменчивости. Эта форма изменчивости поэтому никак не может быть положена в основу объяснения эволюционного процесса. Целесообразность модификаций сама возникает только в процессе эволюции. Отдельная модификация может рассматриваться как закономерное отклонение процесса развития особи (от «нормы») под влиянием изменения известного фактора внешней среды (по сравнению с «нормой»). Если это уклонение адаптивно, то его закономерность тем более обусловлена исторически создавшимся внутренним механизмом развития особи.
Наследственный механизм развития вообще зависим в своих изменениях как от внутренних факторов (мутации), так и от внешних (модификации). В случае ярко выраженной зависимости адаптивных изменений от изменений в факторах внешней среды мы будем говорить о реакциях организма. Закономерность наступления индивидуальной адаптивной реакции безусловно позволяет говорить об историческом обосновании известной более или менее сложной внутренней реакционной системы развивающегося организма. Если адаптивное изменение вида обусловлено изменением в наследственных факторах (генотипе), мы также должны допустить очень сложное согласованное изменение внутреннего механизма развития (может быть, даже того самого, как и в случае адаптивной модификации). В этом случае мы будем говорить об изменении корреляционной системы. Если изменение не адаптив-
144 Организм как целое в индивидуальном и историческом развитии
но, то можно говорить о нарушении корреляционной системы. При рассмотрении конкретной эволюции мы всегда имеем, следовательно, дело с организмами, обладающими уже известной исторически сложившейся структурой и функциями, известным механизмом, обеспечивающим преемственность форм при размножении (наследственность), и известным (реакционным и корреляционным) механизмом, обеспечивающим развитие всей организации при известных условиях внешней среды. Такие живые организмы обладают видовой пластичностью, определяемой, прежде всего, способностью к изменению наследственной основы (мутации) , а также индивидуальной пластичностью, т. е. модификацион-ной изменчивостью. После всех сделанных предварительных замечаний мы все это можем взять уже как данное и рассмотреть, как идет адаптация организма при изменении условий его существования.
5. Выпадение излишних формообразовательных реакций как кажущееся закрепление модификаций (аккомодаций)
Мы отметили в предыдущем, что характер модификационной изменчивости вырабатывается исторически (через естественный отбор) в зависимости от тех условий, в которых живет организм. Если эти условия разнообразны и изменчивы и организм не обладает средствами быстрой защиты или изоляции от резких изменений среды, то обычно модификационная изменчивость приобретает большой размах и иногда приводит к своеобразному мо-дификационному полиморфизму (многие растения).
Если такой индивидуально весьма пластичный организм меняет свою среду (т. е. либо сама среда изменяется, либо организм переселяется в иную среду) на более постоянную, то некоторая его модификационная реакция станет почти постоянной, а другие, рассчитанные (в своем историческом развитии) на такие условия среды, которые здесь уже не встречаются, окажутся совершенно излишними. Однако мы в первой части настоящей работы уже отметили, что реакционные и корреляционные системы, определяющие развитие организма и его частей, не только вырабатываются в процессе естественного отбора, но и поддерживаются при его постоянном участии. Механизмы, потерявшие свое значение для организма, не контролируются отбором, так как их нарушения не сказываются на жизнеспособности организма в данных условиях существования. Поэтому в этих реакционных и корреляционных системах происходит беспорядочное накопление мутационных отклонений (и в первую очередь сдвигов во времени наступления известных реакций), которые приводят эти механизмы к расстройству. Распад реакционных и корреляционных систем имеет своим неизбежным следствием сначала недоразвитие, а затем и полное выпадение тех признаков и черт
Глава IV. Прогрессивная эволюция. Адаптациогенез 145
организации, развитие которых ими контролировалось (редукция органов). Вследствие распада установившегося внутреннего механизма развития исторически приобретенная способность к адаптивной реакции уже не может осуществиться даже при наличии того внешнего раздражителя, который раньше определял наступление этой формообразовательной реакции. Таким образом, у организма сохраняется лишь тот фенотип, который соответствует условиям развития в новой среде. Другие его исторически (в свое время) сложившиеся формы реагирования (модификации) теряются. Если у организма имеется только один ярко выраженный фенотип, то обычно говорят о наследственной структуре (неправильно, так как наследственна лишь норма реакции при данных условиях), и в данном случае, при утере других (якобы ненаследственных) фенотипов, ламаркисты говорят о наследственном «фиксировании» модификаций, о превращении их в мутации и т. п.
В. Кирпичников [1935] выдвигает для объяснения тех же явлений утраты излишних приспособлений гипотезу косвенного OT6opat действующего при эволюции всего организма через систему корреляционных зависимостей также разрушающим образом на механизмы, утратившие свое значение. Эти совершенно правильные соображения должны быть приняты во внимание. Результат получается тот же самый. Мне, однако, кажется, что распад вследствие отсутствия отбора как фактора, поддерживающего данный реакционный механизм, должен происходить быстрее — даже без других изменений, приобретаемых в силу положительного отбора (а без этого не может сказаться его косвенное разрушающее влияние), так как мутации отрицательные, нарушающие развитие, вообще встречаются неизмеримо чаще, чем положительные.
На каком-либо более конкретном примере нетрудно показать полную ошибочность мнения ламаркистов о фиксировании модификации. Возьмем сначала пример растения с довольно широкой нормой реакций, т. е. обладающего значительной модификацион-ной изменчивостью (объясняющейся разнообразием условий, в которых оно живет, например, при широком распространении семян ветром). Возьмем, например, обыкновенный одуванчик, который растет где угодно, в том числе и в горных местностях (рис. 17). В долинах он имеет всем нам знакомый вид, а высоко в горах Дает карликовую форму, напоминая по виду многие другие альпийские растения [Боынье, 1890]. Эта горная модификация одуванчика имеет адаптивный характер — низкорослость (близость к нагретой почве) предохраняет его от вымерзания при ночном охлаждении воздуха, а глубокие корни способствуют использованию небольших количеств воды. Если бы это растение не обладало приспособлением для широкого распространения семян при Помощи ветра, то оно могло бы, так сказать, прочно осесть в го-
146 Организм как целое