
Регуляция активности свободных мультиферментных комплексов цикла кальвина у высших растений
| Вид материала | Автореферат |
- Лекция 11 Тема: Высшие растения. Происхождение высших наземных растений. Отдел Моховидные, 276.22kb.
- Культура столонов и регуляция роста растений и клубнеобразования у картофеля in vitro, 342.87kb.
- Эпигенетическая регуляция фотосинтетической ассимиляции со 2 у хлопчатника (03. 00., 544.35kb.
- План по теме «Анатомия и морфология высших растений» Цель курса, 22.96kb.
- №3. "Фотосинтез", 852.51kb.
- Учебно-методический комплекс дисциплины "Физиология растений" вузовского компонента, 1046.86kb.
- Конференция по систематике и морфологии растений, посвященная 300-летию со дня рождения, 28.08kb.
- Синтез, исследование строения и no-донорной активности нитрозильных комплексов железа, 509.45kb.
- Рабочая программа дисциплины «ботаника» Код дисциплины по учебному плану опд., 301.75kb.
- Общая характеристика высших растений, 83.92kb.
4.1. Возрастные изменения активности свободных мультиферментных комплексов.
В работе (Suss et al., 1993) установлено, что свободные мультиферментные комплексы цикла Кальвина элюировались в широком диапазоне концентраций KCl – 0-600мМ. Молекулярная масса мультиферментных комплексов составляла от 200 до 1000 кД.
Ранее из листьев хлопчатника были выделены свободные мультиферментные комплексы с молекулярной массой 520 и 240 кД (Бабаджанова М.П., Бабаджанова М.А., Алиев К.А., 2006). Мультиферментный комплекс с молекулярной массой 240 кД был выделен в денатурирующих условиях, как и комплекс из листьев шпината с молекулярной массой 284 кД (Rault et al., 1993). Денатурирующие условия при выделении мультиферментных комплексов были использованы для диссоциации их на мультиферментные комплексы с меньшими величинами молекулярных масс.
При выделении из листьев хлопчатника в фазах бутонизации и цветения мультиферментного комплекса в обычных щадящих условиях на профиле элюции свободных белков при ионообменной хроматографии на ДЭАЭ–целлюлозе суммарный белок выходил в виде двух отчётливых пиков, резко различавшихся по содержанию белка. Фракция с наибольшим содержанием белка представляла собой мультиферментный комплекс с молекулярной массой 520 кД. У фракции с меньшим содержанием белков не была определена ни величина молекулярной массы, ни ферментативные активности (Бабаджанова М.П., Бабаджанова М.А., Алиев, 2002, 2006).
На рис. 6 изображён профиль элюции при ионообменной хроматографии на ДЭАЭ - целлюлозе линейным градиентом концентраций NaCl свободных белков, выделенных из листьев хлопчатника сорта 108-Ф в фазе 5-6 настоящих листьев. Как видно на рисунке 6, белки элюировались в виде одного отчётливого пика. Наибольшее количество белка элюировалось во фракциях 3,4,5 при концентрации NaCl 100,150 и 200 мМ соответственно. В фазе бутонизации (рис. 7) отчётливо видны два пика (фракция 4 и 8), резко различавшихся по содержанию белка. При выделении мультиферментного комплекса из листьев хлопчатника в фазе массового цветения профиль элюции белков был такой же, как и в фазе бутонизации. Белки фракции 4 элюировались при концентрации NaCl 150-160 мМ, а белки фракции 8 – при концентрации NaCl 370-380 мМ. Белки фракции 4 (рис.6,7) и 8 (рис. 7) брали для определения молекулярной массы и активности ферментов.
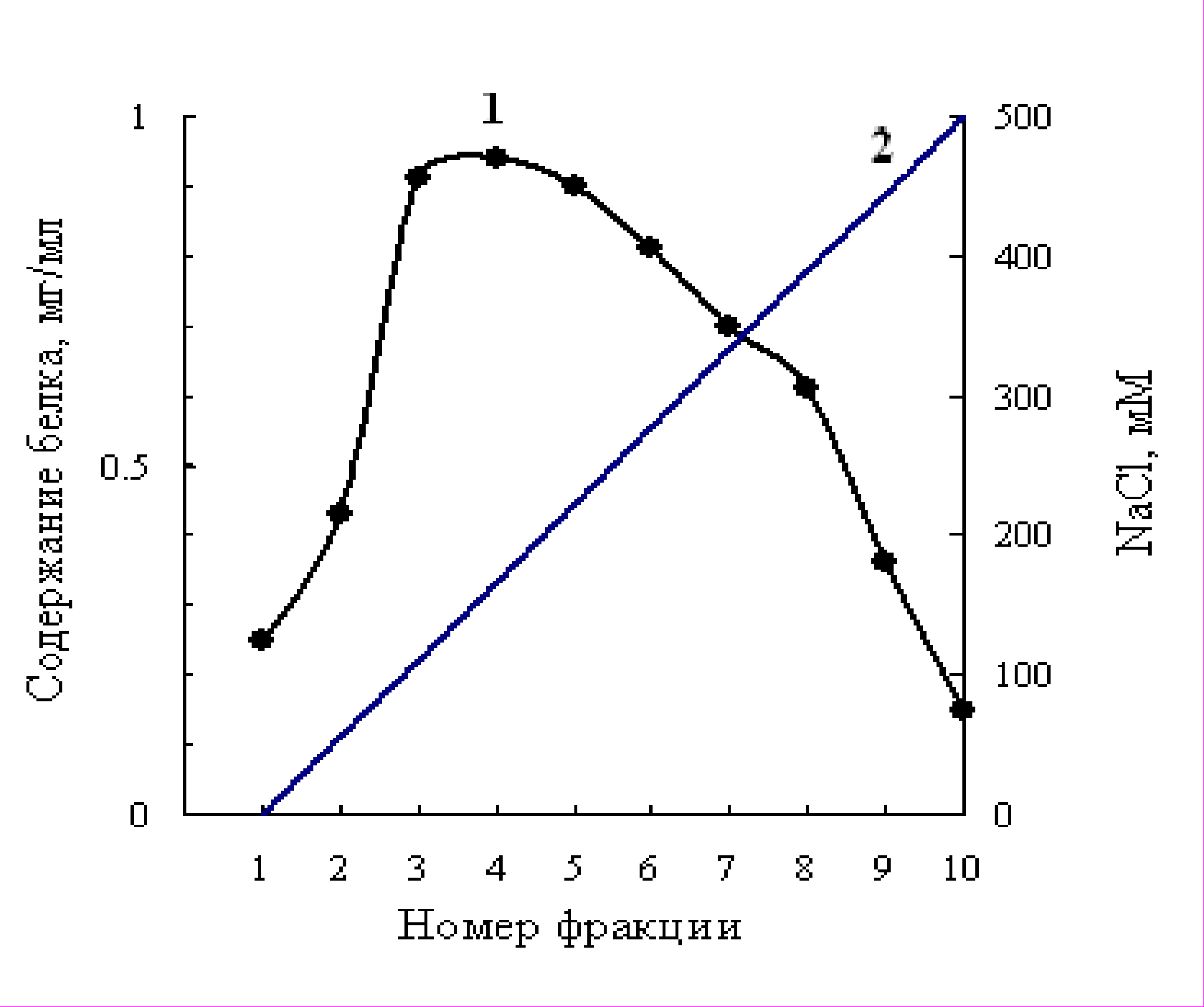
Рис. 6. Профиль элюции белков, выделенных из листьев хлопчатника в фазе 5-6 настоящих листьев, при ионообменной хроматографии на ДЭАЭ–целлюлозе линейным градиентом концентраций NaCl.
Рис. 7. Профиль элюции белков выделенных из листьев хлопчатника в фазе бутонизации, при ионообменной хроматографии на ДЭАЭ–целлюлозе линейным градиентом концентраций NaCl.
Белки 1-го пика (рис. 5.6, фракция 4) проявились в виде одной полосы, молекулярная масса которого оказалась равной 520 ± 20 кД. Начиная с фазы бутонизации растений обнаруживался второй мультиферментный комплекс с молекулярной массой 480 ± 15 кД (рис. 7, фракция 8).
В табл. 3 приведены результаты определения активностей этих мультиферментных комплексов.
Таблица 3
Активности мультиферментных комплексов с различными величинами молекулярных масс, выделенных из листьев хлопчатника сорта 108-Ф на различных фазах развития растений.
| Мол.м., кД | Тип активности | Субстрат | Активность на различных фазах, мкмоль продукта/(мин. мг белка) | ||
| 5-6 настоящих листьев | Бутонизация | Цветение | |||
| 520 | Рибозофосфатизо-меразная (РФИ) | рибозо-5-фосфат | 2673 ± 16 | 3080 ± 18 | 3376 ± 20 |
| 480 | не обнаружен | 3041 ± 17 | 3345 ± 20 | ||
| 520 | Фосфорибулокиназная (ФРК) | рибозо-5-фосфат + АТФ | 3490 ± 21 | 4185 ± 26 | 4432 ± 28 |
| 480 | не обнаружен | 3597± 21 | 3794 ± 25 | ||
| 520 | рибулозо-5-фосфат +АТФ | 3122 ± 19 | 3604 ± 24 | 3715 ± 23 | |
| 480 | не обнаружен | 3412 ± 20 | 3518 ± 22 | ||
| 520 | Рибулозобисфос-фат | рибозо-5-фосфат + АТФ | 1.38 ± 0.01 | 1.92 ± 0.01 | 2.33 ± 0.01 |
| 480 | не обнаружен | 1.68 ± 0.01 | 2.11 ± 0.01 | ||
| 520 | РБФ | 1.14 ± 0.01 | 1.55 ± 0.01 | 1.86 ± 0.01 | |
| 480 | не обнаружен | 1.45 ± 0.01 | 1.74 ± 0.01 | ||
В приведённых в табл. 3 данных видно, что независимо от величины молекулярной массы на всех фазах развития растений фосфорибулокиназная и РБФ-карбоксилазная активности мультиферментных комплексов были выше при использовании в качестве субстрата рибозо-5-фосфата + АТФ, а не в присутствии собственных специфических субстратов рибулозо-5-фосфата+АТФ для фосфорибулокиназы и рибулозо -1.5-бисфосфата для РБФ-карбоксилазы .
В фазе массовой бутонизации по сравнению с фазой 5–6 настоящих листьев величины различных типов активности мультиферментного комплекса с молекулярной массой 520 кД была значительно выше. Рибулозобисфосфаткарбоксилазная активность возросла на 16%, фосфорибулокиназная активность при использовании в качестве субстрата рибозо-5-фосфата + АТФ была выше на 20%, а в присутствии собственного специфического субстрата рибулозо-5-фосфата + АТФ – на 15%. РБФ-карбоксилазная активность мультиферментного комплекса при использованиив качестве субстрата рибозо-5-фосфата + АТФ увеличилась на 39%, а в присутствии собственного специфического субстрата рибулозо-1,5-бисфосфата – на 36%.
Величины всенх типов активностей мультиферментного комплекса с молекулярной массой 480 кД были ниже в сравнении с величинами акимвностей мультиферментного комплекса с молекулярной массой 520кД.
В фазе цветения по сравнению с фазой бутонизации рибозофосфатизомеразная активность у обоих мультиферментных комплексов возрасла на 10%. Фосфорибулокиназная активность как в присутствии собственного специфического субстрата, так и в присутствии рибозо-5-фосфата+АТФ почти не изменилась у обоих мультиферментных комплексов. РБФ-карбоксилазная активность мультиферментного комплекса с мол. м. 520 кД независимо от субстрата была выше на 7–10%, чем у мультиферментного комплекса с мол. м. 480 кД. При этом активность комплекса с мол. м. 520 кД в присутствии собственного специфического субстрата возрасла на 20%, а при использовании в качестве субстрата рибозо-5-фосфата+АТФ – на 26%.
Таким образом, в фазе цветения ферментативная активность мультиферментных комплексов была наибольшей независимо от величины молекулярной массы.
ГЛАВА 5. АКТИВНОСТЬ СВОБОДНЫХ МУЛЬТИФЕРМЕНТНЫХ КОМПЛЕКСОВ ЦИКЛА КАЛЬВИНА В ИСХОДНЫХ И МУТАНТНЫХ ФОРМАХ РАСТЕНИЙ.
5.1. Ферментативные активности свободных мультиферментных комплексов, выделенных из листьев хлопчатника сорта 108-Ф, его мутанта Дуплекс и инбредной линии Л-461.
Выделение и очистка ферментных препаратов, определение их активности проводились многократно. Поскольку во всех случаях были получены очень близкие результаты, в работе приведены типичные примеры.
Сравнительные исследования активностей мультиферментных комплексов из листьев хлопчатника сорта 108-Ф и его мутанта Дуплекс проводились на электрофоретически гомогенных препаратах, выделенных одновременно (в один и тот же день, в один и тот же час) в фазе бутонизации растений.
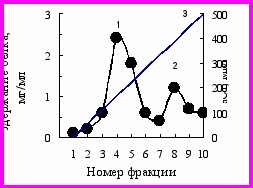
Профиль элюции свободных белков выделенных из листьев хлопчатника сорта 108-Ф и его мутанта Дуплекс при ионообменной хроматографии на ДЭАЭ-целюлозе линейным градиентом концентраций NaCl приведён на рис. 8.
2
1
Линейный градиент концентраций, NaCl
Рис. 8. Профиль элюции свободных белков выделенных из листьев хлопчатника сорта 108-Ф (1) и его мутанта Дуплекс (2) в фазе бутонизации при ионообменной хроматографии на ДЭАЭ – целлюлозе линейным градиентом концентраций NaCl.
Из представленных на рисунке 8 данных видно, что в обоих случаях белки элюировались в виде двух отчётливых пиков, резко различающихся по содержанию белков. По содержанию белка во фракции 6 (пик I) и во фракции 15 (пик II) мутант Дуплекс превосходил исходный сорт хлопчатника 108-Ф. Белки фракции 6 и 15 брали для определения молекулярной массы и активности ферментов.
Для обоих объектов получены аналогичные результаты. Молекулярная масса белка I пика (фракция 6) составляла 520±20кД, а белка II-го пика (фракция 15) - 480±15кД.
Результаты определения активностей электрофоретически гомогенных мультиферментных комплексов с различными величинами молекулярных масс, выделенных из листьев хлопчатника сорта 108-Ф и его мутанта Дуплекс приведены в таблице 4.
Таблица 4
Активности мультиферментных комплексов цикла Кальвина с различными величинами молекулярных масс, выделенных из листьев хлопчатника сорта 108-Ф и его продуктивного мутанта Дуплекс в фазе бутонизации растений.
| Мол. масса, кД | Тип активности | Субстрат | Удельная активность, мкмоль продукта/ мин на 1мг белка | |
| Сорт 108-Ф | Мутант Дуплекс | |||
| 520 480 | Рибозофосфатизомеразная | Рибозо-5-фосфат | 3010±31 2975±30 | 3175±33 3080±29 |
| 520 480 | Фосфорибулокиназная | Рибулозо-5-фосфат | 3625±38 3360±37 | 3820±41 3405±36 |
| 520 480 | Рибулозобисфосфат-карбоксилазная | Рибулозо-1,5-бисфосфат | 1.46±0.05 0.92±0.05 | 1.98±0.05 1.25±0.05 |
Из приведённых в табл. 4 данных видно, что мультиферментные комплексы выделенные из листьев хлопчатника сорта 108-Ф и у мутанта Дуплекс в фазе бутонизации, почти не различались по величинам рибозофосфатизомеразной и фосфорибулокиназной активности. Различия между мультиферментными комплексами были выявлены только по величинам РБФ-карбоксилазной активности. По величинам РБФ-карбоксилазной активности мультиферментные комплексы с различной молекулярной массой, выделенные из листьев мутанта Дуплекс значительно – на 35-36% превосходили аналогичные мультиферментные комплексы, выделенные из листьев исходного сорта хлопчатника 108-Ф.
Поскольку листья инбредной линии Л-461, как и у мутанта Дуплекс отличались от листьев хлопчатника сорта 108-Ф по форфобиологичеким характеристикам, показаниям фотосинтеза и продуктивности, то было важно выделить мультиферментные комплексы из листьев этого генотипа хлопчатника. Из листьев хлопчатника сорта 108-Ф и инбредной линии Л-461 в фазе массового цветения - начала плодообразования, одновременно были выделены электрофоретически гомогенные препараты свободных мультиферментных комплексов и определена их рибулозобисфосфаткарбоксилазная активность.
Профиль элюции свободных белков, выделенных из листьев хлопчатника сорта 108-Ф и линии Л-461 при ионообменной хроматографии на ДЭАЭ-целюлозе линейным градиентом концентраций NaCl представлен на рис. 9.
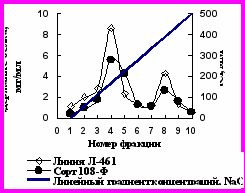
Рис. 9. Профиль элюции свободных белков, выделенных из листьев хлопчатника сорта 108-Ф и линии Л-461, при ионообменной хроматографии на ДЭАЭ-целюлозе линейным градиентом концентраций NaCl.
На рис. 9 видно, что белки элюировались в виде двух отчётливых пиков, резко различающихся по содержанию белка. По содержанию белка во фракции 4 сорт 108-Ф уступал линии Л-461 почти на 50%. Белок фракции 4 (1 пик) – это мультиферментный комплекс с молекулярной массой 520 кД, а белок фракции 8 (2 пик) - мультиферментный комплекс с молекулярной массой 480 кД.
Результаты определения рибулозобисфосфаткарбоксилазной активности мультиферментных комплексов с молекулярной массой 520 и 480 кД приведены в табл. 5.
Таблица 5
Рибулозобисфосфаткарбоксилазная активность мультиферментных комплексов цикла Кальвина из листьев различающихся по продуктивности генотипов хлопчатника. Фаза развития растений – массовое цветение – начало плодообразования.
| Объект | РБФ-карбоксилазная активность МФК, мкмоль СО2/мин на 1 мг белка | |
| 520 кД | 480 кД | |
| Сорт 108-Ф | 2.19 ± 0.01 | 1.36 ± 0.01 |
| Линия Л-461 | 2.73 ± 0.01 | 2.13 ± 0.01 |
Из приведенных в табл.5 данных видно, что мультиферментный комплекс с молекулярной массой 520 кД (1 пик, фракция 4), выделенный из линии хлопчатника Л-461, по рибулозобисфосфаткарбоксилазной активности превосходил сорт 108-Ф на 24%. Величина рибулозобисфосфаткарбокси-
лазной активности мультиферментного комплекса с молекулярной массой 480 кД (2 пик, фракция 8), выделенного из листьев хлопчатника сорта 108-Ф, была меньше на 56% в сравнении с активностью такого же мультиферментного комплекса, выделенного из листьев продуктивной линии Л-461.
Таким образом, мутант Дуплекс и инбредная линия Л-461 отличались от сорта 108-Ф и высоким содержанием водорастворимых белков в экстрактах из листьев, и высоким содержанием мультиферментных комплексов и большей рибулозобисфосфаткарбоксилазной активностью (Бабаджанова; Мирзорахимов и др., 1993; 2007б; 2010).
5.2. Ферментативные активности мультиферментных комплексов из листьев арабидопсиса исходной расы Энкхайм и его мутантов триплекс, 58/15.
Сравнительные исследования ферментативных активностей мультиферментных комплексов из листьев исходной формы арабидопсиса Энкхайм и его двух мутантов проводились на электрофоретически гомогенных ферментных препаратах, выделенных одновременно из листьев растений в фазе сформировавшейся розетки.
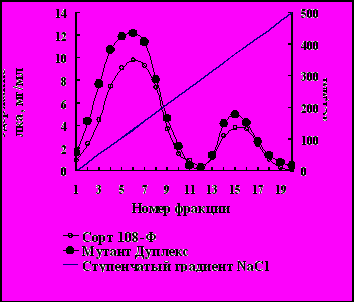
Профиль элюции свободных белков, выделенных из листьев арабидопсиса расы Энкхайм и его мутанта триплекс при ионообменной хроматографии на ДЭАЭ-целлюлозе в фазе формировавшейся розетки представлен на рис. 10.
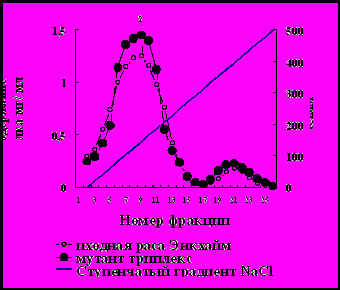
Линейный градиент концентраций NaCl
Рис.10. Профиль элюции свободных белков, выделенных из листьев арабидопсиса расы Энкхайм (I) и его мутантов триплекс (II) в фазе розетки при ионообменной хроматографии на ДЭАЭ-целлюлозе линейным градиентом концентраций NaCl.
На рис. 10 видно, что белки элюировались в виде двух отчётливых пиков, резко различавшихся по содержанию белка. По содержание белка во фракции 8 (пик 1) мутант триплекс превосходил исходную расу Энкхайм. Белки фракций 8 и 20 брали для определения молекулярной массы и активности ферментов. Молекулярная масса белка фракции 8 была равна 520 ± 20 кД, а белка фракции 20 – 480 ± 15 кД.
В табл. 6 приведены результаты определения ферментативных активностей этих мультиферментных комплексов
Таблица 6
Активности мультиферментных комплексов цикла Кальвина с различными величинами молекулярных масс, выделенных из листьев арабидопсиса расы Энкхайм и его продуктивного мутанта триплекс в фазе сформировавшейся розетки.
| Мол.масса, кД | Тип активности | Субстрат | Удельная активность, мкмоль продукта/ мин на 1мг белка | |
| Роса Энкхайм | Мутант триплекс | |||
| 520 480 | Рибозофосфатизомеразная | рибозо-5-фосфат | 2972±28 2986±27 | 3021±32 2992±30 |
| 520 480 | Фосфорибулокиназная | рибулозо-5-фосфат | 2425±22 2375±24 | 3606±38 3101±33 |
| 520 480 | Рибулозобисфосфат-карбоксилазная | рибулозо-1,5-бисфосфат | 1.67±0.05 1.24±0.05 | 1.96±0.05 1.45±0.05 |
Из представленных в табл. 6 данных видно, что у обоих объектов мультиферментные комплексы не различались по величинам рибозофосфатизомеразной активности.
Мультиферментные комплексы с различной молекулярной массой выделенные из листьев исходной расы арабидопсиса Энкхайм не различались по величинам фосфорибулокиназной активности. У мутанта же триплекс мультиферментный комплекс с молекулярной массой 520 кД превосходил по фосфорибулокиназной активности на 16% мультиферментный комплекс с молекулярной массой 480 кД.
По величинам РБФ-карбоксилазной активности мультиферментные комплексы с молекулярной массой 520 кД у обоих объектов превосходили мультиферментные комплексы с молекулярной массой 480 кД на 34-35%.
Величины всех ферментативных активностей мультиферментных комплексов, выделенных из листьев исходной расы Энкхайм были значительно ниже в сравнении с активностями мультиферментных комплексов, выделенными из листьев мутанта триплекс. Мультиферментный комплекс с молекулярной массой 520 кД, выделенный из листьев мутанта триплекс превосходил аналогичный мультиферментный комплекс листьев исходной расы Энкхайм по фосфорибулокиназной активности на 48%, а по РБФ-карбоксилазной –на 17%.
Величины активности мультиферментного комплекса с молекулярной массой 480 кД, выделенного из листьев мутанта триплекс, в сравнении с активностями аналогичного комплекса листьев исходной расы Энкхайм также были выше: фосфорибулокиназная – на 31%, РБФ-карбоксилазная – на 20%.
На рис. 11 представлен профиль элюции при ионообменной хроматографии на ДЭАЭ-целюлозе свободных белков, выделенных из листьев арабидопсиса росы Энкхайм и его низкопродуктивного мутанта 58/15.
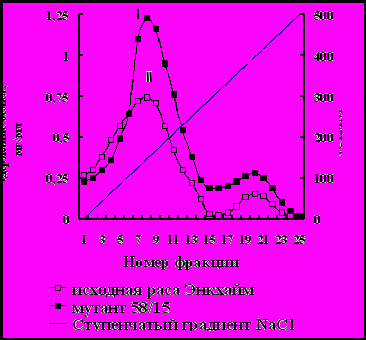
Линейный градиент концентраций, NaCl
Рис. 11. Профиль элюции свободных белков, выделенных из листьев арабидопсиса расы Энкхайм (I) и его мутанта 58/15 (II) в фазе розетки, при ионообменной хроматографии на ДЭАЭ-целюлозе с линейным градиентом концентраций NaCl.
Как видно из рис. 11 белки элюировались в виде двух отчётливых пиков, резко отличающихся по содержанию белка. Белки первого пика (фракция 8) имели молекулярную массу 520±20 кД, молекулярная масса белков второго пика (фракция 20) оказалась равной 480 кД.
В табл. 7 представлены результаты определения активностей мультиферментных комплексов с молекулярной массой 520 и 480 кД, выделенных из листьев мутанта 58/15 в фазе сформировавшейся розетки.
Из представленных в табл. 7 данных видно, что мультиферментные комплексы почти не различались по величинам рибозофосфатизомеразной активности. Различия между мультиферментными комплексами наблюдались по фосфорибулокиназной и РБФ-карбоксилазной активности. Мультиферментный комплекс с молекулярной массой 520кД превосходил мультиферментный комплекс с молекулярной массой 480кД по величине фосфорибулокиназной активности на 11%, а по величине РБФ-карбоксилазной активности - в 2.6 раза.
Таблица 7
Активности мультиферментных комплексов цикла Кальвина с различными величинами молекулярных масс, выделенных из листьев низкопродуктивного мутанта арабидопсиса 58/15.
| Мол.масса, кД | Тип активности | Субстрат | Удельная активность, мкмоль продукта/ мин на 1мг белка |
| 520 480 | Рибозофосфатизомеразная | рибозо-5-фосфат | 2042±23 1811±20 |
| 520 480 | Фосфорибулокиназная | рибулозо-5-фосфат | 2056±22 1895±19 |
| 520 480 | Рибулозобисфосфат-карбоксилазная | рибулозо-1,5-бисфосфат | 1.41±0.05 1.12±0.05 |
При сравнении ферментативных активностей мультиферментных комплексов, выделенных из листьев исходной расы Энкхайм и его мутантов наибольшие различия между ними выявлены по величинам фосфорибулокиназной и РБФ-карбоксилазной активности. По величинам этих активностей мультиферментные комплексы, выделенные из листьев продуктивного мутанта триплекс, значительно превосходили аналогичные мультиферментные комплексы, выделенные из листьев исходной расы Энкхайм и его низкопродуктивного мутанта 58/15.
Таким образом, независимо от генотипов растений мультиферментные комплексы, выделенные из листьев продуктивных мутантов, значительно превосходили исходные формы по содержанию водорастворимых белков в экстрактах из листьев мультиферментных комплексов и РБФ-карбоксилазной активности .